
Abstract
The coffee industry holds importance, providing livelihoods for millions of farmers globally and playing a vital role in the economies of coffee-producing countries. Environmental conditions such as drought and temperature fluctuations can adversely affect the quality and yield of coffee crops.Carotenoid cleavage oxygenases (CCO) enzymes are essential for coffee plants as they help break down carotenoids contributing to growth and stress resistance. However, knowledge about the CCO gene family in Coffee arabica was limited. In this study identified 21 CCO genes in Coffee arabica (C. arabica) revealing two subfamilies carotenoid cleavage dioxygenases (CCDs) and 9-cis-epoxy carotenoid dioxygenases (NCED) through phylogenic analysis. These subfamilies exhibited distribution patterns in terms of gene structure, domains, and motifs. The 21 CaCCO genes, comprising 5 NCED and 16 CCD genes were found across chromosomes. Promoter sequencing analysis revealed cis-elements that likely interact with plant stress-responsive, growth-related, and phytohormones, like auxin and abscisic acid. A comprehensive genome-wide comparison, between C. arabica and A. thaliana was conducted to understand the characteristics of CCO genes. RTqPCR data indicated that CaNCED5, CaNCED6, CaNCED12, and CaNCED20 are target genes involved in the growth of drought coffee plants leading to increased crop yield, in a conditions, with limited water availability. This reveals the role of coffee CCOs in responding to abiotic stress and identifies potential genes useful for breeding stress-resistant coffee varieties.
Similar content being viewed by others


Introduction
Carotenoids, the naturally occurring isoprenoids, are found in prokaryotes, fungi, bacteria, and plants [1, 2]. The carotenoid biosynthesis is carried out and implemented by the higher plants, algae, fungi, and bacteria; animals can take and consume carotenoids through the diet [3]. Furthermore, abscisic acid and strigolactone which are apocarotenoid hormones, play the role of carotenoids precursors [4]. Plants can use carotenoids for many necessary biological processes that determine plant growth and development [5]. Carotenoids are cleaved into various smaller compounds, which play crucial roles in signaling mechanisms and the production of hormones and volatile compounds [6], Carotenoids are not only the chemical precursors of phytohormones but also have very important roles in signaling transduction, growth, and development of plants [2, 7]. Many studies have already shown the various biological roles of the Carotenoid cleavage oxygenases (CCO) gene family in plants, including their implication in pigmentation, photosynthesis, protection against light, response to abiotic stresses, and the biosynthesis of aroma volatiles and plant hormones [8, 9]. Carotenoid Cleavage Oxygenases (CCOs) are the enzymes that break down the conjugated double bonds of carotenoid polyene bonds and thus form different apocarotenoids and their derivatives [10]. The CCOs of plants can be classified into two main subfamilies carotenoid cleavage dioxygenase (CCD) and 9-cis-epoxy carotenoid (NCED) depending on whether they promote the epoxidation of their substrates [11]. As a result of this, these carotenoids, with conjunctive double bonds, are metabolized differently through CCOs. This process is known as carotenoid cleavage oxygenases which are a kind of dioxygenase enzyme occurring in plants [12, 13].
The 9-Cis-epoxy carotenoid dioxygenase (NCED) is a key enzyme of ABA production, which is found in almost all plants. Its subfamily shows remarkable evolutionary conservation, with little variation and significant exon preservation, emphasizing the need to maintain its structural integrity for proper functioning [14,15,16,17]. The slow evolution of NCED genes specifies the multiple functions in different tissues of plants [15]. The diverse expression patterns and fluctuating activities of NCED in different parts of the plant may contribute to the tolerance of the plant to drought conditions and the production of abscisic acid (ABA) hormones to prepare the plant to coordinate plant responses to environmental signals [18, 19]. The discovery of the NCED gene greatly advanced our understanding of ABA production and the function of this hormone in plant growth. NCED’s involvement first identified in the maize ABA-deletion mutant Vp14 and in its overall biosynthesis process, was elucidated [20]. Additional research was undertaken in Arabidopsis revealing 9 CCO genes, of which 5 (AtNCED2, AtNCED3, AtNCED5, AtNCED6, and AtNCED9) had a direct association with ABA biosynthesis. The modification of AtNCED3 levels has been shown to boost drought tolerance through ABA accumulation [21]. NCED genes had the highest level of co-expression with the production of abscisic acid (ABA). All the sites of 9-cis-epoxy carotenoid cleavages of NCED enzymes, generated C15-xanthoxin - the potent precursor of ABA [22, 23]. The study of the NCED genes in several species such as Tamatim [24], cowpea [25], and rice [26] greatly increased our understanding of the evolution and functionality of NCED across plant taxa. This data put forward NCED as crucially vital in the ABA formation pathway that enables plants to make responses to environmental stress and developmental signals. Specifically, all NCED genes in plants mediate the biosynthesis of ABA and can cleave violaxanthin and neoxanthin into xanthaldehyde [27], which is the critical rate-limiting step and the first stage of ABA biosynthesis in plants [27]. After that, the xanthaldehyde is moved into the cytoplasm, where it undergoes a sequence of events that ultimately result in the formation of the plant hormone ABA. Different NCED genes have been studied in several plant species such as Arabidopsis [28], citrus [29], cotton [30], cucumber [31], wheat [32], and tobacco [23]. However, the NCED genes’ role in coffee is not completely known [12].
In the model plant Arabidopsis thaliana, the CCO family has four carotenoid cleavage dioxygenases (CCDs) (CCD1, CCD4, CCD7, and CCD8) [31]. This was followed by another important discovery, one of the members of the CCO family which was found, in S. Lycopersicum, and was named, CCDL [23, 33]. Studies of the genes of the CCD subfamily showed that these genes were involved in different physiological and developmental molecular processes in plants, including photosynthesis, response to abiotic or synthetic stresses, and apocarotenoids synthesis that include aromatic volatiles and strigolactones (SLs) [34, 35]. Studies show that CCD1 and CCD4 can selectively cleave β-carotene into the volatile, aromatic compound β-ionone. [36]. This is one of the main components of the floral scent of the plant [37]. At the same time, the cleavage activity of CCD4 can dramatically decrease the accumulation of β-carotene in plastids, which is also the main factor that affects the color of flowers and fruits of some plants [38]. CCD7 and CCD8 genes in the family form a complex regulatory network in association with other genes that mediate plant development and the biosynthesis of plant hormone aurolactone in a majority of plants [39]. Gene expression can regulate CCD7 and CCD8 genes when plants are developing and undergoing morphogenesis [40]. CCD gene family has already been discovered and studied in several plant species such as Arabidopsis [41], rice [42], sorghum [42], wheat [43], watermelon [31], pepper [44], tobacco [45], rapeseed [46], and maize [23].
Genomic data for the majority of polyploids could not be obtained without requiring a significant investment of time and resources. Thus, the understanding of polyploid genome evolution has mainly been limited to model systems [47]. Recent significant advancements in DNA sequencing technologies have addressed this shortcoming. As a result, there are now exciting opportunities to study the genomes of polyploid plant species, both with and without previously sequenced genomes [48]. In this work, genetic alterations in the allopolyploid C. Arabica were examined using mRNA-seq data [49]. C. arabica, one of the most important commercial crops in tropical and subtropical developing nations, is a member of the Rubiaceae family, which has 124 species. It produces about 30% of the coffee beans produced worldwide [50]. C. arabica L. is an allotetraploid species (2n = 4x = 44) displaying a diploid-like meiotic behavior, that is believed to have been formed through the spontaneous hybridization of two diploid species, C. canephora and C. eugenioides [51]. C. canephora the species, also known as “robusta coffee” is considered one of the parents of C. arabica, which is responsible for approximately 70% of coffee production worldwide [52]. As per the latest national statistics, the average weekly consumption of coffee per person in Japan is approximately 11 cups [53]. The health effects of coffee are gaining increasing popularity [54].
This research also sheds light on the functional differentiation and evolutionary history of CCO genes in plants. Our primary objective was to identify the function and expression patterns of CCO genes in the coffee genome. We utilized RT-PCR and various bioinformatics tools to investigate their functions. The genome-wide identification and characterization conducted in this study will serve as a foundation for the cloning and functional analysis of these genes.
Materials and methods
Sequence retrieval
We obtained the gene annotation files of C. Arabica from the Phytozome database (https://phytozome-next.jgi.doe.gov/). The A. thaliana CCO peptide sequences were downloaded from NCBI (https://www.ncbi.nlm.nih.gov). To locate the CCO genes, in C. Arabica we utilized the RPE65 (PF03055.16) from the NCBI database as a query to perform Blast at Phytozome against the protein sequences of C. Arabica. The search, in the C. Arabica database resulted in identifying twenty-one sequences, which were cross-checked using NCBI CCD (Conserved Domain Database: http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) with parameters to confirm their accuracy [55].
Physio-chemical and subcellular analysis of CCO gene
The ProtParam (https://web.expasy.org/protparam/) tool was used to find out details, like the protein size, molecular weight, and isoelectric point of CaCCO peptides, for gene names, chromosomal positions, and protein sequences of CCO proteins referred to the Phytozome. To determine where the CCO genes were located within organelles we relied on the WoLF PSORT (https://wolfpsort.hgc.jp/) program [56].
Analysis of gene structure, cis-regulatory elements, and motifs
The Gene Structure Display Server (GSDS) v2.0 (http://gsds.cbi.pku.edu.cn/) was utilized to display the intron-exon structure of CCO genes. The PlantCare database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html) was employed to analyze cis-elements (CRES) associated with these genes using 1500 upstream promoter sequences. For motifs identification the MEME suit (http://meme.nbcr.net/meme/) program, with a value of 10 motifs was utilized. The discovered motif was visualized using TB tools [57].
Phylogenetic analysis
Phylogenetic analysis was constructed using the CCO amino acid (AA) sequences of C. Arabica, A. tequilana, A. thaliana, and S. lycopersicum. Using a bootstrapping value of 1000 replications, a phylogenetic tree was constructed from the aligned protein sequence using the maximum-joining (MJ) program in MEGA 11. ITOL (https://itol.embl.de/upload.cgi) was used to show and visualize the derived phylogenetic tree [58].
Duplication analysis, chromosomal mapping, and synteny investigation
The divergence period of the CCO genes was determined using the Ka/Ks ratio, with the TB tools. Gene pairs were calculated by measuring the Ka/Ks ratio of genes [59]. The time of divergence (DT) was then calculated using the formula T = Ks/2λ, where λ represents the substitution rate (6.56*10^-9). Gene duplication events were analyzed using MCScanX v1.0 with default settings to assess collinearity. A. thaliana and C. arabica were the two crops studied for synteny and a synteny graph was created using TB tools circus module. The start and end positions of the CCO gene were identified in the Phytozome database and its chromosomal mapping was done with Tbtools [16].
Protein interaction studies
The study also confirmed the protein interactions among CaCCO genes by utilizing the String database v0.761 (https://string-db.org). This online resource contributed to an accurate description of the complicated web of interactions among protein domains and these CaCCO genes of coffee [16, 56].
Plant material and experiment design
We investigated the C. arabica cultivar CAPBG2008, using seeds sourced from the Department of Plant Breeding and Genetics, University of the Punjab. The seeds of CAPBG2008 were planted in controlled conditions. Drought stress was initiated at the two-leaf stage. Each cultivar had five treated plants (5T) and five controlled plants (5 C), each with three replicates. In the control group, irrigation was regularly applied, while in the drought-treated group, irrigation was withheld for two weeks. Irrigation in the control blocks was administered uniformly. After four cycles of drought stress treatments, samples were collected from this experiment [60,61,62].
CaNCEDgene expression analysis in drought-stressed leaf by qRT-PCR
RNA extraction was carried out using the RNAeasy Isolation Reagent by Vazyme, in Nanjing, China. The RNA quality was evaluated through 0.8% agarose gel electrophoresis while its purity and concentration were determined using a NanoDrop 2000 Spectrophotometer. RNA samples with OD260/OD280 ratios falling between 1.90 and 2.10 were deemed suitable for examination. To analyze the CaNCED gene RNA samples underwent reverse transcription into cDNA utilizing the Hifair™ II 1st Strand cDNA Synthesis SuperMix for qPCR. Subsequent qRT PCR analyses were performed on a Light Cycler 480 apparatus with a 20 µL reaction mixture was treated with 5x gDNA Eraser at 37℃ for 5 min, followed by 65℃ for 2 min. Then, 5xqRT premix II and reverse transcript enzyme mix were used. RT reaction was conducted using 37℃ for 15 min; 98℃ for 2 min, and were kept 4℃. The Livak method was employed to determine the levels of gene transcripts, with each RT qPCR analysis consisting of three replicates [62, 63] (Table 1).
Results
Identification of CCO gene in C.arabica
A genome-wide analysis was conducted to identify the CaCCO genes within the coffee genome. The RPE65 (PF03055) domain served as the query in a BlastP search against the coffee genome hosted on Phytozome v13. Subsequently, 21 CaCCO proteins were identified and analyzed for CCO domains using the Pfam database. Furthermore, the molecular weights of CaCCO genes ranged between (CaCCD11) 51867.1 and (CaNCED5)18504.96 Da, averaging 53442.8581 Da. The approximate isoelectric points (pI) varied from 4.95 (CaNCED5) to 8.84 (CaNCED6), with an average of 6.182. The grand average of hydropathicity (GRAVY) ranges from − 0.808 (CaCCD8) to -0.167 (CaCCD1), with an average of -0.290. Upon closer inspection of gene orientations, it was found that twelve CaCCO genes were aligned in the forward direction, while the remaining ten were oriented in the reverse direction. The aliphatic index indicated a range of values from 72.96 to 90.14, with an average of 79.73, suggesting that all CaCCO proteins were likely to be stable at high temperatures (Table 2).
Examining the subcellular localization of the 21 CaCCO genes, 15 were found to be subcellularly located, with the cytoplasm, peroxisomes, and chloroplasts housing the majority of the genes. A small number were found in the nucleus and mitochondria. The smallest ones were also found, among other places, in plasmids, extracellular structures, vacuoles, and E.R (Fig. 1).

Heat map showing the subcellular locations of all 21 CaCCO genes in the plant cell’s mitochondria, plasmid, chloroplast, nucleus, and Golgi apparatus. The color purple indicates the highest functional significance of the relevant gene in the indicated region, while the color green indicates the least functional presence of the relevant gene in the specified region
Phylogenetic analysis
A phylogenetic tree was built using the reference sequences of C. arabica (Ca), A. tequilana (Ag), Arabidopsis (At), and S. lycopersicum (Sl) were systematically categorized into two distinct clades labeled I-II (NCED and CCD respectively). The study encompassed a total of 53 CCO genes, with 21 from C. arabica 13 from A. tequilana, 9 from A. thaliana, and 10 from S. lycopersicum. To enhance clarity and facilitate a comprehensive understanding of the phylogenetic relationships, each clade was denoted by a specific color scheme (Fig. 2) (S Table 1).

The phylogenetic relationships of CCO genes from four distinct crops of C. arabica, A. thaliana, A. tequilana, and S. lycopersicum were ascertained using a phylogenetic tree method, which is frequently used in research on Arabidopsis. The two CCO genes in the figure divide the four crop species according to the corresponding evolutionary features and traits. Clade I was represented by purple (CCD) and Clade II by orange color (NCED)
Analysis of gene structure, and motifs
In examining the exon-intron structures of five of the twenty-one genes CaNCED12, CaNCED13, CaNCED20, CaNCED5, and CaNCED6 stood out with a unique profile, featuring a singular exon and an absence of introns. The three genes CaCCD15, CaCCD9, CaCCDD2, and CaCCD4 have thirteen exons and twelve introns. CaCCD16 and CaCCD17 contained six exons and five introns and CaCCD7 had only one intron and two exons. These observations highlight significant genomic variations within the CaCCO gene family, highlighting the diversity in their exon-intron structure (Fig. 3).

The intron-exon structure is phylogenetically represented, and it shows that CCD genes contain less coding sequences than NCED genes. The number of introns and exons in several NCED and CCD genes has remained constant
Through our analysis of conserved motifs among 21 CaCCO, genes were subjected to motif analysis, revealing the presence of 10 different motifs. Interestingly, motifs one and seven were found to be conserved in nineteen genes. In addition, motif 3 seems to be present in all except one gene, showing that this motif plays a unique trait or regulatory function in the majority of the genes. Although, 10 motifs were found in seven genes, and six motifs were found in 8 genes, suggesting that a bunch of genes may have the same regulatory processes. This observation suggests that each CaCCO gene possesses distinct functions (Fig. 4).

The distribution of 10 motifs along the 21 members of the C. arabica CCO family
In the domain analysis of the 21 CaCCO proteins, a remarkable uniformity was observed, with all proteins featuring a singular domain identified as the RPE65 superfamily. This major domain further comprises subfamilies such as RPE65, PLN02258, PLN02969, and PLN02491. Particularly, the RPE65 superfamily is present in nine CaCCD proteins and two CaNCED proteins, while PLN02258 is found in three CaNCED proteins (CaNCED12, CaNCED13, and CaNCED20) exclusively. Moreover, RPE65 is exposed in 4 proteins of CaCCD, PLN02969 in 2 proteins of CaCCD and a single protein of CaCCD (CaCCD9). This demonstrates the conservation of the RPE65 domain in all 21 CaCCO proteins (Fig. 5).

Conserved Domain analysis leads to the identification that all 21 CaCCO proteins have a conserved RPE65 superfamily domain, which demonstrates more than one particular regulatory role in plant growth and development
Evaluation of duplication event of C. Arabica
During the investigation of Ka/Ks ratios, there were 90 duplicated pairs of CaCCO genes identified in the coffee genome. The results of Ka/Ks analysis showed that CaCCD11_CaCCD15 were found to have higher values of 1.0335. Contrariwise, CaCCD7_CaNCED12 showed lower Ka/Ks values (0.10843). Divergence time estimation, measured in million years ago (MYA), validates these findings. Lower MYA values for pairs like CaNCED5_CaNCED20 (0.6681) suggest more recent divergence; while higher MYA values for pairs such as CaCCD3_CaNCED13 (499.95) indicate ancient divergence from a common ancestor (Fig. 6)(S Table 2).

The expression ka/ks represents the ratio of mutations involving synonymous substitutions (ks) to mutations involving non-synonymous substitutions (ka). The gene duplication over selection and evolutionary pressure to paralogous pairings of C. Arabica genes were calculated based on ks and ka values
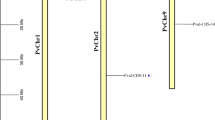
The chromosomal localization study revealed that CaCCO genes were distributed across multiple scaffolds. Specifically, CaCCD1, CaCCD2, CaCCD3, and CaCCD4 were located on scaffold 465, while CaNCED5 and CaNCED6 were found on scaffold 624. CaCCD7 and CaCCD8 were identified on Scaffold 449, CaCCD9 on Scaffold 616, CaCCD10 on Scaffold 634, and CaCCD11 on Scaffold 2123. CaNCED12 and CaNCED13 were situated on scaffold 770. CaCCD18 was present on scaffold 607, CaCCD19 on scaffold 263, CaNCED20 on scaffold 315, and CaCCD21 on scaffold 2652. Finally, CaCCD14, CaCCD15, CaCCD16, and CaCCD17 were located on scaffold 597. These findings provide insights into the chromosomal organization of CaCCO genes (Fig. 7).

The C. arabica genome’s chromosomal mapping of the CCO genes shows the existence of paralogous copies with plausible locations. During the time of selection pressure and chromosomal rearrangement, 5 NCED genes suffered duplication events. Despite these alternations, the duplication of NCED genes remained to be able to keep their original roles and develop favorable functional traits in the C.arabica genome
The syntenic analysis of CaCCO genes reveals both segmental and tandem duplications within the coffee genome. Tandem duplication occurs on the same chromosome and involves the consecutive duplication of DNA segments, resulting in a repeated sequence on the same chromosome. Scaffolds 597, 449, 465 and 624, show evidence of tandem duplication. Segmental duplication can occur on the same chromosome or different chromosomes. It involves the duplication of larger genomic segments, which can be present on the same chromosome or different chromosomes while Scaffolds 770, 2652, 315,263, and 607 indicate Segmental duplication events. This comprehensive analysis highlights the paralogous duplication mechanisms shaping the coffee genome (Fig. 8).

An extensive study of genome-wide synteny in C. arabica CCO genes revealed that genes sharing conserved areas are those with structurally similar sequences. Using blue and green hues, the method provides a visual depiction of the degree of similarity across genomic regions. A variety of dynamic processes and genomic rearrangements that enable genes to stay stable, and secure, adapt to their surroundings, and acquire new properties over time lead to gene duplication
Analysis of CaCCO Cis-regulatory elements
The presence and arrangement of multiple cis-regulatory elements, in the promoter region had an impact on how genes were expressed over time. The PlantCare database was explored to investigate the roles of CaCCO genes in coffee plants. The results revealed a variety of cis-elements in Coffee CCO genes that respond to factors like light, hormones, stress, and growth. In C. arabica the CCO gene family contained 82 elements categorized into responses to phytohormones, stress-responsive, and growth-related signals. The promoter regions displayed motifs such as Skn 1_motif, GCN4_motif, MRE, Box 4, CAT box, O2 site, and circadian elements; among these were motifs like TGA element influencing auxin sensitivity and ABRE motif associated with gibberellin response. Phytohormone responses included ABRE P box TGACG motif, TCA element, and CGTCA motif linked respectively to SA (acid) ABA (acid) MeJA (methyl jasmonate), and ethylene signaling pathways. Moreover; stress-responsive elements like ARE LTR MBS W box were tied to exposure well as cold and drought stress offering valuable insights into how these regulatory mechanisms strengthened the resilience of the CaCCO gene family, against environmental pressures. Analysis of cis-regulatory elements revealed that a significant portion of elements (45.06%) were linked to plant hormones, containing motifs such as CGTCA-motif and TGACG-motif for MeJA response, TCA-element for SA response, GARE-motif, TATC-box, and P-box for GA response, ABRE for ABA response, and TGA-element for auxin response. The second-largest group (26%) responded to light, featuring motifs like Box 4, MRE, and G-box. The third-largest group (27%) was associated with abiotic and biotic stress, containing motifs like LTR, TC-rich repeats, ARE, MBS, and W box. (Fig. 9) (S Table 3).

The cis-regulatory analysis of the CCO gene and the magnitude of each function are shown graphically. The intensity of the biochemical and physiological processes occurring in plants is ranked from red (highest) to blue (lowest)
Analysis of protein-protein interaction network
During a protein interaction investigation, a total of 9 nodes and 14 edges were observed. The average node degree was calculated to be 14, with an average local clustering coefficient of 3.11. Interestingly, the expected number of edges in this scenario was zero, and the p-value for protein-protein interaction enrichment was remarkably low at < 1.0e-16, indicating a significant enrichment of interactions. To meet the minimum required interaction score, a low confidence threshold of 0.150 was applied. Among the 21 CaCCO proteins examined, interactions were observed for only nine proteins (CaCCD18, CaNCED20, CaCCD15, CaCCD1, CaNCED13, CaCCD19, CaCCD17, CaCCD21, and CaCCD14). Notably, CaCCD18, CaNCED13, and CaCCD21 exhibited the highest number of interactions, being associated with 9 proteins, while the remaining proteins showed associations within themselves (Fig. 10).

Based on protein-protein interaction, nine genes were associated with the 21 CaCCO proteins. In the dataset, CaCCD18, CaNCED13, and CaCCD21 exhibited the highest number of interactions, being associated with 9 proteins
Quantitative analysis of CaNCED gene expression in drought-stressed leaves
To clarify the expression patterns of various NCED genes in response to abiotic stress, specifically drought stress, controlled conditions were employed. Quantitative analysis of CaNCED revealed that four out of the five genes studied, namely CaNCED5, CaNCED6, CaNCED12, and CaNCED20, exhibited significant regulation in response to the drought conditions. Interestingly, all the significant genes displayed downregulation. Conversely, CaNCED13 showed upregulation under drought stress, although this change was not statistically significant (Fig. 11).

The expression of NCED genes in response to abiotic stress (drought stress), in controlled conditions was employed. Quantitative analysis of CaNCED revealed that four out of the five genes showed significant results (Less than 0.05=*, Less than 0.01=**, ns = non significant).Graph was constructed with R programming language
Discussion
Drought stress severely affects coffee farming, which results in a 40–80% loss of yield and plays a role in affecting the physiology, growth, and quality of plants [60]. It makes coffee more susceptible to pests and diseases, thereby, necessitating the use of molecular techniques such as genome-wide identification of specific gene families like CCO, which are capable of combating abiotic stresses. Over the past few years, bioinformatics analysis has extensively explored the CCO gene family across various species [64, 65]. However, research on the CaCCO gene family in coffee has been comparatively limited, resulting in a knowledge gap regarding CCOs in coffee. This study identified and characterized 21 CaCCO genes in the coffee genome, shedding light on potential genes and pathways for developing resilient coffee varieties against abiotic stresses [66].
The physicochemical parameters of CaCCO genes in the coffee genome were evaluated to detect differences among proteins within the same clade. All identified CCO proteins displayed hydrophilic characteristics with negative GRAVY values, indicating an affinity for water interaction and net electrical charges at varying pH levels [31]. The aliphatic index showed that all of the 21 proteins were likely to be stable at high temperatures. Subcellular localization analysis showed divergent distribution of CaCCO proteins into chloroplasts, mitochondria, cytoplasm, cytosol, endoplasmic reticulum, nucleus, and plasma membrane. A remarkable fact was that most of the proteins were found in the Peroxisome (24%, 72 of 295), while cytoplasm and chloroplasts had equal proportions (19%, 57 of 295), implying that CaCCO proteins might have a significant function within these organelles [67]. Phylogenetic analysis can assist in understanding functional genomics by revealing similarities among subgroups [11, 33]. In our study, 53 CCO proteins with complete domain sequences were classified into two subfamilies based on sequence structures and phylogenetic relationships. The analysis identified five CaNCED proteins and sixteen CaCCD proteins, indicating potential functional similarities to AtNCED and AtCCD proteins, respectively, within this subgroup [68, 69].
Previous studies have emphasized the significance of exon-intron organization in the evolution of gene families [11]. The analysis revealed that members within the same population and clade shared similar exon, intron, and motif distributions, consistent with the structure of phylogenetic trees. While exons were universally present in all CCO genes, introns were absent in some proteins [70]. Notably, the NCED gene subfamily exhibited more conserved motifs compared to the CCD subfamily, a common feature in plant genomes. Additionally, cis-regulatory elements located within gene promoter regions play a critical role in regulating transcriptional activity and influencing gene expression patterns [70].
These findings suggest potential roles of CaCCO genes in regulating responses to various stresses, including drought [12]. Analyzing genomes across species offers insights into gene evolution and organization, facilitating the transfer of genomic data from well-studied taxa to less-explored ones [45]. In this study, we identified 90 pairs of paralogous genes in the genome, likely originating from gene duplication events. The duplication analysis revealed that Divergence time estimation revealed varying divergence times, with lower MYA values suggesting more recent divergence for pairs like CaNCED5_CaNCED20, while higher MYA values indicate ancient divergence for pairs such as CaCCD3_CaNCED13. Such duplications yield valuable insights into the expansion of gene families, a prevalent phenomenon in plants driven by tandem and segmental duplications [23].
Drought stress could reduce photosynthetic rates and transpiration in plants, resulting in crop yield losses [71]. Stomata were crucial in plant photosynthetic activities and transpiration [72]. The quantitative analysis of CaNCED revealed that four out of the five genes, including CaNCED5, CaNCED6, CaNCED12, and CaNCED20, exhibited significant downregulation in response to drought stress. Conversely, CaNCED13 showed upregulation under drought stress, although this change was not statistically significant. These findings suggested that a majority of CaNCED genes in coffee plants were susceptible to drought stress, as they demonstrated downregulation. These results helped build the concept of the adaptive and evolutionary value of the CaCCO genes. Extensive screening of a genome and detailed characterization performed by the experiment will help prepare coffee varieties resistant to abiotic stresses.
Conclusion
In this research, twenty-one CCO genes were found in the coffee genome, exhibiting different intron numbers varying from one to thirteen. The existence of cis-regulatory elements that respond to light, developmental progression stages, hormone signaling, and abiotic stress within the promoter regions of the CaCCO gene explains their roles in coffee plants’ reaction to abiotic stress. RT-qPCR data analysis revealed that the CaNCED5, CaNCED6, CaNCED12, and CaNCED20 genes could be useful in developing drought-resistant coffee varieties for the sake of higher yield under drought conditions. However, further research, including gene cloning and functional analysis, is necessary to confirm the significance of these genes across various physiological and biological processes.
Data availability
The data associated with this study are available in the manuscript.
References
Ahrazem O, Diretto G, Argandoña J, Rubio-Moraga Á, Julve JM, Orzáez D, Granell A, Gómez-Gómez L. Evolutionarily distinct carotenoid cleavage dioxygenases are responsible for crocetin production in Buddleja davidii. J Exp Bot. 2017;68(16):4663–77.
Sami A, Haider MZ, Shafiq M. Microbial nanoenzymes: Features and applications. In: Fungal Secondary Metabolites Elsevier; 2024: 353–367.
Schweiggert R, Carle R. Carotenoid deposition in plant and animal foods and its impact on bioavailability. Crit Rev Food Sci Nutr. 2017;57(9):1807–30.
Nisar N, Li L, Lu S, Khin NC, Pogson BJ. Carotenoid metabolism in plants. Mol Plant. 2015;8(1):68–82.
Cazzonelli CI. Carotenoids in nature: insights from plants and beyond. Funct Plant Biol. 2011;38(11):833–47.
Sun T, Rao S, Zhou X, Li L. Plant carotenoids: recent advances and future perspectives. Mol Hortic. 2022;2(1):3.
Peng Y, Zhang X, Liu Y, Chen X. Exploring heat-response mechanisms of microRNAs based on microarray data of rice post-meiosis panicle. International Journal of Genomics 2020, 2020.
Zhang X, Pei J, Zhao L, Tang F, Fang X, Xie J. Overexpression and characterization of CCD4 from Osmanthus fragrans and β-ionone biosynthesis from β-carotene in vitro. J Mol Catal B: Enzymatic. 2016;134:105–14.
Ren C-G, Kong C-C, Xie Z-H. Role of abscisic acid in strigolactone-induced salt stress tolerance in arbuscular mycorrhizal Sesbania cannabina seedlings. BMC Plant Biol. 2018;18(1):1–10.
Ahrazem O, Gómez-Gómez L, Rodrigo MJ, Avalos J, Limón MC. Carotenoid cleavage oxygenases from microbes and photosynthetic organisms: features and functions. Int J Mol Sci. 2016;17(11):1781.
Yue X-Q, Zhang Y, Yang C-K, Li J-G, Rui X, Ding F, Hu F-C, Wang X-H, Ma W-Q, Zhou K-B. Genome-wide identification and expression analysis of carotenoid cleavage oxygenase genes in Litchi (Litchi chinensis Sonn). BMC Plant Biol. 2022;22(1):394.
Sami A, Haider MZ, Shafiq M, Sadiq S, Ahmad F. Genome-wide identification and in-silico expression analysis of CCO gene family in sunflower (Helianthus Annnus) against abiotic stress. Plant Mol Biol. 2024;114(2):1–19.
Sami A, Han S, Haider MZ, Khizar R, Ali Q, Shafiq M, Tabassum J, Khalid MN, Javed MA, Sajid M, et al. Genetics aspect of vitamin C (ascorbic acid) biosynthesis and signaling pathways in fruits and vegetables crops. Funct Integr Genom. 2024;24(2):73.
Sun L, Sun Y, Zhang M, Wang L, Ren J, Cui M, Wang Y, Ji K, Li P, Li Q. Suppression of 9-cis-epoxycarotenoid dioxygenase, which encodes a key enzyme in abscisic acid biosynthesis, alters fruit texture in transgenic tomato. Plant Physiol. 2012;158(1):283–98.
Liu X, Li X, Yang H, Yang R, Zhang D. Genome-wide characterization and expression profiling of ABA biosynthesis genes in a desert moss Syntrichia caninervis. Plants. 2023;12(5):1114.
Ali M, Shafiq M, Haider MZ, Sami A, Alam P, Albalawi T, Kamran Z, Sadiq S, Hussain M, Shahid MA et al. Genome-wide analysis of NPR1-like genes in citrus species and expression analysis in response to citrus canker (Xanthomonas axonopodis Pv. Citri). Front Plant Sci 2024, 15.
Ali Q, Sami A, Haider MZ, Ashfaq M, Javed MA. Antioxidant production promotes defense mechanism and different gene expression level in Zea mays under abiotic stress. Sci Rep. 2024;14(1):7114.
Wan T, Liu Z, Leitch IJ, Xin H, Maggs-Kölling G, Gong Y, Li Z, Marais E, Liao Y, Dai C. The Welwitschia genome reveals a unique biology underpinning extreme longevity in deserts. Nat Commun. 2021;12(1):4247.
Bhatti MHT, Sami A, Haider MZ, Shafiq M, Naeem S, Tariq MR, Ahmad S, Irfan U. Genetic Diversity of Vegetable Crops and Utilization in Food and Nutritional Security. In: Sustainable Utilization and Conservation of Plant Genetic Diversity Edited by Al-Khayri JM, Jain SM, Penna S. Singapore: Springer Nature Singapore; 2024: 171–197.
Felemban A, Braguy J, Zurbriggen MD, Al-Babili S. Apocarotenoids involved in plant development and stress response. Front Plant Sci. 2019;10:478231.
Hu Q, Ao C, Wang X, Wu Y, Du X. GhWRKY1-like, a WRKY transcription factor, mediates drought tolerance in Arabidopsis via modulating ABA biosynthesis. BMC Plant Biol. 2021;21:1–13.
Dhar MK, Mishra S, Bhat A, Chib S, Kaul S. Plant carotenoid cleavage oxygenases: structure–function relationships and role in development and metabolism. Brief Funct Genomics. 2020;19(1):1–9.
Haider MZ, Sami A, Shafiq M, Anwar W, Ali S, Ali Q, Muhammad S, Manzoor I, Shahid MA, Ali D. Genome-wide identification and in-silico expression analysis of carotenoid cleavage oxygenases gene family in Oryza sativa (rice) in response to abiotic stress. Front Plant Sci 2023, 14.
Burbidge A, Grieve TM, Jackson A, Thompson A, McCarty DR, Taylor IB. Characterization of the ABA-deficient tomato mutant notabilis and its relationship with maize Vp14. Plant J. 1999;17(4):427–31.
Iuchi S, Kobayashi M, Yamaguchi-Shinozaki K, Shinozaki K. A stress-inducible gene for 9-cis-epoxycarotenoid dioxygenase involved in abscisic acid biosynthesis under water stress in drought-tolerant cowpea. Plant Physiol. 2000;123(2):553–62.
Zhu G, Ye N, Zhang J. Glucose-induced delay of seed germination in rice is mediated by the suppression of ABA catabolism rather than an enhancement of ABA biosynthesis. Plant Cell Physiol. 2009;50(3):644–51.
Pei X, Wang X, Fu G, Chen B, Nazir MF, Pan Z, He S, Du X. Identification and functional analysis of 9-cis-epoxy carotenoid dioxygenase (NCED) homologs in G. Hirsutum. Int J Biol Macromol. 2021;182:298–310.
Tan BC, Joseph LM, Deng WT, Liu L, Li QB, Cline K, McCarty DR. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 2003;35(1):44–56.
Rodrigo M-J, Alquezar B, Zacarías L. Cloning and characterization of two 9-cis-epoxycarotenoid dioxygenase genes, differentially regulated during fruit maturation and under stress conditions, from orange (Citrus sinensis L. Osbeck). J Exp Bot. 2006;57(3):633–43.
Li Q, Yu X, Chen L, Zhao G, Li S, Zhou H, Dai Y, Sun N, Xie Y, Gao J. Genome-wide identification and expression analysis of the NCED family in cotton (Gossypium hirsutum L). PLoS ONE. 2021;16(2):e0246021.
Cheng D, Wang Z, Li S, Zhao J, Wei C, Zhang Y. Genome-wide identification of CCD gene family in six Cucurbitaceae species and its expression profiles in melon. Genes. 2022;13(2):262.
Lang J, Fu Y, Zhou Y, Cheng M, Deng M, Li M, Zhu T, Yang J, Guo X, Gui L. Myb10-D confers PHS‐3D resistance to pre‐harvest sprouting by regulating NCED in ABA biosynthesis pathway of wheat. New Phytol. 2021;230(5):1940–52.
Wei Y, Wan H, Wu Z, Wang R, Ruan M, Ye Q, Li Z, Zhou G, Yao Z, Yang Y. A comprehensive analysis of carotenoid cleavage dioxygenases genes in Solanum lycopersicum. Plant Mol Biology Report. 2016;34:512–23.
Ahrazem O, Rubio-Moraga A, Argandona-Picazo J, Castillo R, Gómez-Gómez L. Intron retention and rhythmic diel pattern regulation of carotenoid cleavage dioxygenase 2 during crocetin biosynthesis in saffron. Plant Mol Biol. 2016;91:355–74.
Chen H, Zuo X, Shao H, Fan S, Ma J, Zhang D, Zhao C, Yan X, Liu X, Han M. Genome-wide analysis of carotenoid cleavage oxygenase genes and their responses to various phytohormones and abiotic stresses in apple (Malus domestica). Plant Physiol Biochem. 2018;123:81–93.
Qi Z, Tong X, Zhang X, Lin H, Bu S, Zhao L. One-pot synthesis of dihydro-β-ionone from carotenoids using carotenoid cleavage dioxygenase and enoate reductase. Bioprocess Biosyst Eng. 2022;45(5):891–900.
Qi Z, Fan X, Zhu C, Chang D, Pei J, Zhao L. Overexpression and characterization of a novel plant carotenoid cleavage dioxygenase 1 from Morus notabilis. Chem Biodivers. 2022;19(2):e202100735.
Li T, Deng YJ, Liu JX, Duan AQ, Liu H, Xiong AS. DcCCD4 catalyzes the degradation of α-carotene and β‐carotene to affect carotenoid accumulation and taproot color in carrot. Plant J. 2021;108(4):1116–30.
Xue G, Hu L, Zhu L, Chen Y, Qiu C, Fan R, Ma X, Cao Z, Chen J, Shi J. Genome-wide identification and expression analysis of CCO Gene Family in Liriodendron chinense. Plants. 2023;12(10):1975.
Gao J, Zhang T, Xu B, Jia L, Xiao B, Liu H, Liu L, Yan H, Xia Q. CRISPR/Cas9-mediated mutagenesis of carotenoid cleavage dioxygenase 8 (CCD8) in tobacco affects shoot and root architecture. Int J Mol Sci. 2018;19(4):1062.
Priya R, Sneha P, Dass JFP, Manickavasagam M, Siva R. Exploring the codon patterns between CCD and NCED genes among different plant species. Comput Biol Med. 2019;114:103449.
Vallabhaneni R, Bradbury LM, Wurtzel ET. The carotenoid dioxygenase gene family in maize, sorghum, and rice. Arch Biochem Biophys. 2010;504(1):104–11.
Qin X, Fischer K, Yu S, Dubcovsky J, Tian L. Distinct expression and function of carotenoid metabolic genes and homoeologs in developing wheat grains. BMC Plant Biol. 2016;16:1–15.
Yao Y, Jia L, Cheng Y, Ruan M, Ye Q, Wang R, Yao Z, Zhou G, Liu J, Yu J. Evolutionary origin of the carotenoid cleavage oxygenase family in plants and expression of pepper genes in response to abiotic stresses. Front Plant Sci. 2022;12:792832.
Zhou Q, Li Q, Li P, Zhang S, Liu C, Jin J, Cao P, Yang Y. Carotenoid cleavage dioxygenases: identification, expression, and evolutionary analysis of this gene family in tobacco. Int J Mol Sci. 2019;20(22):5796.
Zhou X-T, Jia L-D, Duan M-Z, Chen X, Qiao C-L, Ma J-Q, Zhang C, Jing F-Y, Zhang S-S, Yang B. Genome-wide identification and expression profiling of the carotenoid cleavage dioxygenase (CCD) gene family in Brassica napus L. PLoS ONE. 2020;15(9):e0238179.
Soltis DE, Gitzendanner MA, Stull G, Chester M, Chanderbali A, Chamala S, Jordon-Thaden I, Soltis PS, Schnable PS, Barbazuk WB. The potential of genomics in plant systematics. Taxon. 2013;62(5):886–98.
Sun Y, Shang L, Zhu Q-H, Fan L, Guo L. Twenty years of plant genome sequencing: achievements and challenges. Trends Plant Sci 2022.
Lloyd A, Blary A, Charif D, Charpentier C, Tran J, Balzergue S, Delannoy E, Rigaill G, Jenczewski E. Homoeologous exchanges cause extensive dosage-dependent gene expression changes in an allopolyploid crop. New Phytol. 2018;217(1):367–77.
Batista-Santos P, Lidon F, Fortunato A, Leitão A, Lopes E, Partelli F, Ribeiro A, Ramalho J. The impact of cold on photosynthesis in genotypes of Coffea spp.—photosystem sensitivity, photoprotective mechanisms and gene expression. J Plant Physiol. 2011;168(8):792–806.
Lashermes P, Combes M-C. Diversity and genome evolution in coffee. Achiev Sustainable Cultivation Coffee BDS Publ Camb 2018:3–20.
Perrois C, Strickler SR, Mathieu G, Lepelley M, Bedon L, Michaux S, Husson J, Mueller L, Privat I. Differential regulation of caffeine metabolism in Coffea arabica (Arabica) and Coffea canephora (Robusta). Planta. 2015;241:179–91.
Nakagawa-Senda H, Hachiya T, Shimizu A, Hosono S, Oze I, Watanabe M, Matsuo K, Ito H, Hara M, Nishida Y. A genome-wide association study in the Japanese population identifies the 12q24 locus for habitual coffee consumption: the J-MICC study. Sci Rep. 2018;8(1):1493.
Cano-Marquina A, Tarín J, Cano A. The impact of coffee on health. Maturitas. 2013;75(1):7–21.
Domingues DS, Oliveira LS, Lemos SM, Barros GC, Ivamoto-Suzuki ST. A Bioinformatics Tool for efficient Retrieval of high-confidence terpene synthases terpene synthases (TPS) and application to the identification of TPS in Coffea and Quillaja. Plant secondary Metabolism Engineering: methods and protocols. Springer; 2022. pp. 43–53.
Shafiq M, Manzoor M, Bilal M, Manzoor T, Anees MM, Rizwan M, Haider MZ, Sami A, Haider MS. Genome-Wide Analysis of Plant Specific YABBY Transcription Factor Gene Family in Watermelon (Citrullus lanatus) and Arabidopsis. J Appl Res Plant Sci. 2024;5(01):63–78.
Islam MAU, Nupur JA, Shafiq M, Ali Q, Sami A, Shahid MA. In silico and computational analysis of zinc finger motif-associated homeodomain (ZF-HD) family genes in Chilli (Capsicum annuum L). BMC Genomics. 2023;24(1):603.
Hussain M, Javed MM, Sami A, Shafiq M, Ali Q, Mazhar HS-U-D, Tabassum J, Javed MA, Haider MZ, Hussain M, et al. Genome-wide analysis of plant specific YABBY transcription factor gene family in carrot (Dacus carota) and its comparison with Arabidopsis. BMC Genomic Data. 2024;25(1):26.
Ahmad F, Tomada S, Poonsiri T, Baric S. Molecular genetic variability of Cryphonectria hypovirus 1 associated with Cryphonectria parasitica in South Tyrol (northern Italy). Front Microbiol 2024, 15.
Mofatto LS, Carneiro FA, Vieira NG, Duarte KE, Vidal RO, Alekcevetch JC, Cotta MG, Verdeil J-L, Lapeyre-Montes F, Lartaud M, et al. Identification of candidate genes for drought tolerance in coffee by high-throughput sequencing in the shoot apex of different Coffea arabica cultivars. BMC Plant Biol. 2016;16(1):94.
Paula MFBd, Ságio SA, Lazzari F, Barreto HG, Paiva LV, Chalfun-Junior A. Efficiency of RNA extraction protocols in different types of coffee plant tissues. 2012.
Huded AKC, Jingade P, Mishra MK. A rapid and efficient SDS-based RNA isolation protocol from different tissues of coffee. 3 Biotech. 2018;8(3):183.
Ge Y, Qiu H, Zheng J. Physicochemical characteristics and anti-hyperlipidemic effect of polysaccharide from BaChu mushroom (Helvella Leucopus). Food Chemistry: X. 2022;15:100443.
Dong X, Yang Y, Zhang Z, Xiao Z, Bai X, Gao J, Hur Y, Hao S, He F. Genome-wide identification of WRKY genes and their response to cold stress in Coffea canephora. Forests. 2019;10(4):335.
Almas M, Sami A, Shafiq M, Bhatti M, Haider M, Hashmi M, KHALID M. Sale price comparison of saggian flower market: a case study. Bull Biol Allied Sci Res. 2023;2023(1):39–39.
Zanin FC, Freitas NC, Pinto RT, Máximo WPF, Diniz LEC, Paiva LV. The SAUR gene family in coffee: genome-wide identification and gene expression analysis during somatic embryogenesis. Mol Biol Rep 2022:1–12.
Ji F, Wu J, Zhang Z. Identification and characterization of CCD Gene Family in Rose (Rosa chinensis Jacq.‘Old blush’) and Gene Co-expression Network in Biosynthesis of Flower Scent. Horticulturae. 2023;9(1):115.
Zhang S, Guo Y, Zhang Y, Guo J, Li K, Fu W, Jia Z, Li W, Tran L-SP, Jia K-P. Genome-wide identification, characterization and expression profiles of the CCD gene family in Gossypium species. 3 Biotech. 2021;11(5):249.
Bülow L. R Hehl 2016 Bioinformatic identification of conserved cis-sequences in coregulated genes. Plant Synth Promoters: Methods Protocols 233 245.
Wei H, Liu G, Wang Y, Chen J, Chen Y, Lian B, Zhong F, Yu C, Zhang J. Genome-Wide Identification and Expression Analysis of Carotenoid Cleavage Oxygenase Genes in Crape Myrtle. Available at SSRN 4294416.
Farooq M, Hussain M, Wahid A, Siddique K. Drought stress in plants: an overview. Plant responses to drought stress: From morphological to molecular features 2012:1–33.
Liang YS, Jeon Y-A, Lim S-H, Kim JK, Lee J-Y, Kim Y-M, Lee Y-H, Ha S-H. Vascular-specific activity of the Arabidopsis carotenoid cleavage dioxygenase 7 gene promoter. Plant Cell Rep. 2011;30:973–80.
Acknowledgements
We would like to thank the Key Laboratory of kiwifruit resources development and utilization of Guizhou Universities (Qian Jiaoji [2022] 054), projects of Liupanshui Normal University (Biological Science, LPSSYYlzy2003, LPSSY2023XKTD09, Lpssyzxxm202304, LPSSYKYJJ201601) and the Science and Technology project of Liupanshui City (Grant #52020-2020-0906).
Funding
This work was supported by the Key Laboratory of kiwifruit resources development and utilization of Guizhou Universities (Qian Jiaoji [2022] 054), projects of Liupanshui Normal University (Biological Science, LPSSYYlzy2003, LPSSY2023XKTD09, Lpssyzxxm202304, LPSSYKYJJ201601) and the Science and Technology project of Liupanshui City (Grant #52020-2020-0906).
Author information
Authors and Affiliations
Contributions
SN, YW, and SH wrote the original draft and performed data analysis. MZH and AS did in-silico experiments, and data analysis, and edited the manuscript, MS, and PA edited the manuscript and performed data analysis, MHTB, AA, and JD edited the manuscript, and acquired the funding. IAS, MAM, and Q. Ali co-supervised the work, conceptualized, reviewed, and edited the manuscript, and provided the infrastructure.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Data transparency
The authors ensured that all data, materials, software applications, and custom codes supported the claims made in this article and fully complied with field standards. The authors have also considered the possibility of individual journal policies regarding research data sharing, considering the norms and expectations of our discipline. Therefore, the data is available in the supplementary materials or deposited in online databases.
Competing interest
The authors have no potential conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Naeem, S., Wang, Y., Han, S. et al. Genome-wide analysis and identification of Carotenoid Cleavage Oxygenase (CCO) gene family in coffee (coffee arabica) under abiotic stress. BMC Genom Data 25, 71 (2024). https://doi.org/10.1186/s12863-024-01248-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12863-024-01248-4