
Abstract
This paper discusses the species composition and population structure of small mammals inhabiting the north-taiga landscape of the Nadym Hills (Yamalo-Nenets Autonomous Okrug, Russia) with an emphasis on catenary patterns in the intralandscape differentiation. Five small mammal species (Cl. rutilus, S. caecutiens, S. araneus, A. oeconomus, and E. sibiricus) typical for the north-taiga subzone of Western Siberia are recorded in different parts of a northwest-facing landscape catena (upland, saddle, slope, and floodplain). In terms of abundance, the northern red-backed vole (Cl. rutilus) predominates in all catena localities and in the landscape as a whole. Other species codominate in certain localities, with the exception of S. araneus, who is common only on the upland. Throughout the entire study area and in the upper localities of the catena, Cl. rutilus and S. caecutiens form the core of small mammal communities. The total abundance of small mammals and the abundance of the dominant species gradually decline along the slope (i.e., from the upland to the floodplain). Resident and propagating small mammal populations are present in all catena localities. The composition, structure, and parameters of small mammal communities change along the slope following changes in afforestation, wetting, plant species richness, vegetation structure, and microtopography. In all catena localities and throughout the entire study area, small mammal communities feature low values of the diversity, evenness, and sustainability indices and high values of the dominance index, which is determined by harsh natural and climatic conditions. However, their generally undisturbed structure is typical for natural subarctic communities, which indicates their sustainability.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Sustainable nature use and biodiversity conservation in northern regions of Russia require knowledge of the impact exercised by modern global environmental and climatic changes that are most pronounced at high latitudes on natural complexes in these regions (Pavlov and Malkova, 2010; Moskalenko, 2012; Tishkov, 2012; Mokhov et al., 2020; Ferronsky, 2013). Accordingly, issues associated with assessing the state of biodiversity and dynamics at different organization levels of living systems and identifying biological indicators reflecting changes in regional landscapes are extremely relevant. Landscape catenae (i.e., series of morphological landscape units from the watershed to the foot of the slope (Chibilyov et al., 1999; Chernykh et al., 2022) can be used as regional models that make it possible to assess the state of biodiversity in modern conditions. The catenary approach assumes that changes in geosystem parameters occur as a result of the redistribution of moisture, loose material, chemical elements, and their compounds between relief elements along the slope vector. Since most of the terrain is represented by slopes of varying steepness, the catena is the most common organizational form of terrestrial landscapes (Mordkovich et al., 1985). Accordingly, the purpose of this study was to examine the species composition and population structure of small mammals as one of the components and status indicators of a typical north-taiga landscape taking into account changes in conditions along the catena vector.
MATERIALS AND METHODS
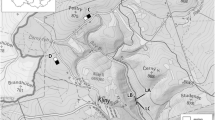
The field studies were conducted in the north-taiga subzone, northwest of the city of Nadym (Yamalo-Nenets Autonomous Okrug, Western Siberia), within the so-called Nadym Hills. The Nadym Hills are a suitable test polygon for identifying catenary patterns in landscape differentiation. Some of these hills located in the contact zone with the Nadym River valley are composed of light loamy deposits, which makes it possible to consider tops of these hills uplands whose conditions match the zonal values to the greatest possible extent. The test polygon is distinguished by significant (up to 40 m) elevation differences (Gataullin, 1991; Gosudarstvennaya geologicheskaya karta…, 2015; Chernykh et al., 2022).
Field landscape studies were conducted from July 13 to July 17, 2021. Six localities have been distinguished along a northwest-facing slope stretching from the top of one of the hills towards the Ngarka-Veloyakha River (a Nadym River tributary), and landscape descriptions of these localities were produced (Table 1). Each locality was described in accordance with the methodology commonly accepted in landscape science (Beruchashvili and Zhuchkova, 1997), including microtopography description, the establishment and description of a soil profile cut, and the description of a botanical sampling plot 10 × 10 m in size.
Small mammals were caught on sampling plots established on four of the six catenary positions (upland (ND-02-21), saddle (ND-05-21), upper part of the slope (ND-06-2), and floodplain (ND-03-21)) (Table 1) from July 13 to July 17, 2021, and from July 9 to July 14, 2022, using lines of break-back traps and pitfall traps (Karaseva et al., 2008; Ravkin and Livanov, 2008). The total exposure was 1075 break-back trap-days (hereinafter trap-days) and 250 cylinder-days (hereinafter cyl.-days); 82 animals were captured. Data on the numbers of exposed traps of different types and animals captured in various habitats in different years are presented in Table 2. The species of captured animals were identified based on a set of morphological characters (Vinogradov and Gromov, 1952; Yudin, 1989).
Abundance was expressed in number of individuals per 100 trap-days (or 100 cyl.-days). To ensure the comparability of abundance estimates with data collected by other researchers, values obtained using break-back traps were recalculated into ind./100 cyl.-days (Ravkin and Livanov, 2008). A complete morphophysiological examination of captured animals was performed (Shvarts et al., 1968). Their relative age was determined based on the thymus gland condition, teeth condition, and skull structure. The spatial distribution of species was quantified using the biotopic allocation index (Fij) (Pesenko, 1982).
The structure of communities was analyzed using diversity indices and multidimensional icon plots constructed on their basis (Bigon et al., 1989; Litvinov, 2004, 2008), sustainability indicators, and integrated community parameters (Gashev, 2000). In accordance with M. Bigon et al. (1989), diversity indices were computed based not only on abundance parameters, but also based on density (ind./km2) and biomass (g/km2) values estimated in accordance with Yu.S. Ravkin and I.V. Luk’yanova (1976). Due to the semiarboreal lifestyle of the Siberian chipmunk (Eutamias (=Tamias) sibiricus Laxmann, 1769), its density was estimated on the basis of its occurrence in snap traps using bird density formulas (Ravkin and Livanov, 2008) and assuming that the reliable detection distance does not exceed 25 m for this species. The community of small mammals was considered “a territorial grouping of phylogenetically similar species belonging to the same or adjacent trophic levels” (Brown, 1995; Litvinov, 2004, 2008). A comparative analysis of community structure was performed based on our materials collected in other subarctic regions of Western Siberia in 2019 and 2021–2022.
The total sampling effort was 5875 trap-days and 860 cyl.-days; in total, 441 individuals belonging to 12 species were captured.
Statistical data processing was performed using the following computer programs: Statan (Gashev, 2011), Theriologist’s Workplace (Gashev et al., 2012), and Microsoft Excel.
RESULTS
Distinct Features of the Nadym Catena
Let us examine geosystems of the Nadym Hills in the context of environmental factors that have a significant impact on the spatial structure of their small mammal population (Ravkin and Luk’yanova, 1976) and are manifested differentially within the studied catena (Table 1).
Relief
Microtopography changes along the slope vector. On the upland and in the upper part of the slope, mounds are formed at the base of trees, pronounced cryogenic heaving was noted in the saddle, terracing in the middle and lower parts of the slope, and tussocks in the floodplain.
Soils
All catena elements feature an elongated soil profile, a deep upper boundary of perennially frozen and parent rocks, high humus content in upper horizons, and high organic carbon content. The exception is the saddle locality, where perennially frozen rocks are located close to the surface, and parent rocks occur at a depth of less than 1 m. All drained localities of the studied catena, with the exception of the saddle, are distinguished by textural differentiation of the soil profile and the absence of pronounced gleyization traces.
Anthropogenic Impact
The absence of permafrost within the soil profile and the high organic carbon content discovered in the course of landscape studies could be determined by frequent fires (once every 30–50 years) (Mergelov, 2015), resulting in the presence of numerous piece of coal in the soil profile (Chernykh, 2022).
Vegetation
The plant species richness gradually increases from the upland to the floodplain. Concurrently, in areas with stagnant and sinter wetting (i.e., in the saddle and in the middle part of the slope, respectively), the local plant species richness is slightly higher than in adjacent localities due to the greater diversity of microbiotopes.
The tree layer in the upper and middle parts of the catena is formed by four species: Pinus sibirica Du Tour, Larix sibirica Ledeb., Picea obovata Ledeb., and Betula subarctica Orlova. In addition, Salix caprea L. occurs in the lower part of the slope, while the coenosis-forming role of L. sibirica sharply decreases in the floodplain since it prefers more stable wetting conditions. The canopy density on the upland is average; in the saddle, it decreases by half, and then it sharply increases and reaches its maximum in the floodplain (Table 1). The shrub layer is absent on the upland and in the upper part of the slope, but it is present in locations with higher moisture supply. For instance, Betula nana L. s. str. and B. czerepanovii Orlova grow in the saddle. Lower down the catena, Salix glauca L., S. phylicifolia L. s. str., Rosa acicularis Lindl., Lonicera pallasii Ledeb., and Juniperus communis L. appear in the shrub layer. The grass–dwarf-shrub layer undergoes the most significant changes along the catena vector (Table 1). The following dwarf shrubs predominate among the eight species recorded on the upland and in the upper part of the slope: Ledum palustre L., Vaccinium vitis-idaea L., V. uliginosum L., V. myrtillus L., and Empetrum nigrum L. In the saddle and in the middle part of the slope, the species diversity in this layer increases due to the presence of swamp species: Oxycoccus microcarpus Turcz. ex Rupr., Petasites frigidus (L.) Fries, etc. In the lower part of the slope and in the floodplain, as the moisture supply, soil nutrition, and microbiotopic diversity change, species richness increases due to the presence of tall grasses (Chamaenerion angustifolium (L.) Scop., Angelica decurrens (Ledeb.) B.Fedtsch., Cacalia hastata L., Senecio nemorensis L., Thalictrum minus L., Veratrum lobelianum Bernh. etc.), gramineous species (Calamagrostis spp. and Poa sp.), taiga short grasses (Trientalis europaea L., Lycopodium annotinum L., etc.), and swamp and meadow forbs (Equisetum pratense Ehrh., Galium boreale L., G. uliginosum L., Rubus arcticus L., etc.). The total projective cover of the grass–dwarf-shrub layer reaches its maximum in the upper part of the slope and in the floodplain, while in the wettest localities (i.e., in the saddle and in the middle part of the slop), it decreases (Table 1).
Composition and Population Structure of Small Mammals
In total, five small mammal species have been recorded within the study area; they represent two orders: Eulipotyphla (Laxmann’s shrew (Sorex caecutiens Laxmann, 1788) and the common shrew (Sorex araneus L., 1758)) and Rodentia (northern red-backed vole (Clethrionomys (=Myodes) rutilus Pallas, 1779), the tundra vole (Alexandromys oeconomus Pallas, 1776), and the Siberian chipmunk (Eutamias (=Tamias) sibiricus Laxmann, 1769)). The name Clethrionomys is provided as the main generic synonym of the northern red-backed vole in accordance with B. Kryštufek et al. (2019), who substantiated its validity; other species names are provided in accordance with systematic catalogues (Pavlinov and Lisovsky, 2012; Lisovsky and al., 2019).
S. caecutiens and A. oeconomus belong to the transpalaearctic fauna type, S. araneus to the western (European) palaearctic type, and Cl. rutilus and E. sibiricus to the eastern (Siberian) palearctic type (Yudin, 1989; Volpert and Shadrina, 2002; Bobretsov, 2016).
A typical forest species, Cl. rutilus, makes the main contribution to the total abundance of small mammals. In 2021, it demonstrated a broad ecological valence within the study area: Cl. rutilus did not give preference to any of the catena localities and only slightly avoided the floodplain (Fij = –0.44) (Table 3). The maximum abundance of the species was noted on the upland and in the saddle; then it gradually decreased along the slope vector. The Cl. rutilus dominance index changes in a similar way: from 100% in the saddle (2022) and in the upper part of the slope (2021) to 33–50% in the floodplain. The abundance of other species throughout the entire study area is an order of magnitude lower compared to Cl. rutilus; however, their abundance levels are comparable to each other. Some catena localities are codominated by different species: upland by S. caecutiens (dominance index = 12.8%), saddle by S. caecutiens and E. sibiricus (11.1 and 5.6%, respectively), upper part of the slope by E. sibiricus (14.3%), and floodplain by A. oeconomus and E. sibiricus (33.3 and 33.3–50%). The analyzed sample indicates the stenotopicity of A. oeconomus, who occurs only in the floodplain, and the clear biotopic preference given to this habitat by E. sibiricus (Fij > 0.7). This can be explained by the ecological and biological characteristics of these species: A. oeconomus prefers wet floodplain biotopes, while the floodplain offers more diverse sources of seed foods for E. sibiricus. S. caecutiens does not demonstrate biotopic preferences in the upper part of the catena and is absent in samples collected on the slope and in the floodplain. S. araneus (2.6%) was caught only on the upland. Overall, throughout the study area and in the upper part of the catena (on the upland and in the saddle, where catches were richer), Cl. rutilus and S. caecutiens form the core of the small mammal community; their cumulative share in the total number of individuals was 92.7, 97.4, and 96.3%, respectively.
In 2021, the total abundance of small mammals in all studied localities of the Nadym catena was an order of magnitude higher than in 2022; this can be primarily attributed to different phases of the population cycle of the dominant species (Cl. rutilus), whose abundance fluctuates in a similar way (Table 3). It is obvious that other species feature similar population dynamics: in 2022, their numbers were so low that, despite the sufficient sampling effort, they were absent in catches.
Numerical abundance scores of individual species (Ravkin and Luk’yanova, 1976) vary by years and by habitats. In 2021, Cl. rutilus was numerous throughout the study area and in upper parts of the catena (on the upland and in the saddle) and common on the slope and in the floodplain. Other species were rare throughout the entire study area, but common in certain habitats: S. caecutiens on the upland and in the saddle (based on the results of the pitfall-trap census, it is abundant in the saddle), S. araneus on the upland, E. sibiricus in the saddle and in the floodplain, and A. oeconomus in the floodplain. In 2022, despite the larger number of exposed trap-days, rodents were not caught on the upland, while insectivores were not caught at all—not in a single habitat—which can be explained by their extremely low abundance levels. Still, Cl. rutilus remains a common species both throughout the study area and in all parts of the catena. E. sibiricus is common in the floodplain, but rare in the upper part of the slope and throughout the entire study area. In both years, the catches did not include very rare species with an abundance of less than 0.1 ind./100 trap-days (cyl.-days).
The unevenness of the spatial distribution of small mammals (huddling) increases along the slope vector as their total abundance decreases, the microrelief changes, and the species richness in lower vegetation layers goes up; this is manifested in a series of increasing values of the poor aggregation index from upper to lower catena localities (Table 4). High values of the conservatism index reflecting the total proportion of females and overwintered animals (Gashev, 2000) recorded in all habitats indicate that resident and propagating small mammal populations are present everywhere. In both years, the reproductive success index gradually increased along the slope vector; this can be explained by compensatory population processes activated under the conditions of lower population density featured by both small mammals in general and by the dominant species in lower parts of the catena.
Based on their anthropophobia degrees, Cl. rutilus, A. oeconomus, S. caecutiens, and E. sibiricus belong to the neutral ecological group, while S. araneus belongs to the group of synanthropes (Gashev, 2000). Values of the integrated anthropogenic adaptation index, which reflects specific contributions of various ecological groups to the community taking into account their ranked ecological and biological characteristics (K- and r-orientation, anthropophobia degree, consumption degree, preferred humidity, and habitat closeness degree) (Gashev, 2000), are the same for small mammal communities inhabiting all localities of the catena and throughout the entire study area (A = 1.11) and are identical to values obtained for small mammal communities studied by the authors in the natural north-taiga habitats of Western Siberia (vicinity of the Shukhtungort cordon, Malaya Sos’va State Nature Reserve, Beryozovo district, Khanty-Mansi Autonomous Okrug, and the vicinity of the Sterkh station, Kunovatskii Regional Wildlife Sanctuary, Shuryshkarskii district, Yamalo-Nenets Autonomous Okrug).
The composite well-being index reaches its lowest values in communities inhabiting the saddle (localities with stagnant wetting) and the highest values in communities inhabiting floodplains. Throughout the entire study area, this parameter featured average values in 2022 and below average values in 2021. This can be explained by the harsh natural and climatic conditions of this region.
Let us examine biomass as an emergent property of the studied communities and diversity indices and information multidimensional icon plots constructed for the Shannon (H) and Simpson’s (D) species diversity indices and the Shannon (J) and Simpson’s (E) evenness indices (Magurran, 1988; Litvinov, 2004, 2008) as a measure of species diversity.
For all small mammal species and communities as a whole, biomass trends by years and habitats are similar to abundance trends, since the density of species that statistically significantly differ from the dominant in body weight is insignificant (Table 3).
Small mammal communities in individual habitats and throughout the entire study area are distinguished by very low values of the species richness, species diversity, and evenness indices (as well as sustainability indices) and high values of the dominance index (Table 4). This is typical for high-latitude communities due to harsh natural and climatic conditions in their habitats. Concurrently, values of all community diversity and sustainability indices increase from the upland to the floodplain, with the exception of the dominance index, whose changes are inverse to the slope vector and are consistent with changes in diversity. Differences between upland and floodplain communities in diversity indices reach 1.7–3.3 times, in the dominance index 1.5–2.2 times, and in sustainability indices 1.2–13.6 times. Interestingly, resistant sustainability exceeds resilient sustainability by 2.2–3.0 times in communities inhabiting all localities of the catena (except for the floodplain); this is typical for communities inhabiting transformed landscapes and habitats at early succession stages (i.e., young communities) (Gashev, 2000). By contrast, in the floodplain community, the contribution of the resilient component to the overall sustainability is 2.5 times higher compared to the resistant component, which is typical for undisturbed habitats.
According to the methodological approach substantiated by Yu.N. Litvinov (2004, 2008), the shape of multidimensional icon plots can be used as a qualitative characteristic of the information structure and sustainability of communities (samples) and landscapes (sites) (Litvinov, 2004, 2008). Multidimensional icon plots constructed for the studied communities based on the computed abundance, density, and biomass indices have different scales, but a similar shape, which indicates the validity of diversity estimates produced on the basis of the available material (i.e., primary data on numbers of individuals belonging to different species). For communities with the participation of E. sibiricus inhabiting the saddle, slope, and floodplain, this indicates inter alia the adequacy of the approach used to estimate the density of this species (Fig. 1).

Multidimensional icon plots constructed for information diversity indices computed for small mammal communities inhabiting different localities of the Nadym catena (multidimensional icon plots in the left row were constructed for indices computed based on abundance parameters; in the central row, based on density; and, in the right row, based on biomass).
Multidimensional icon plots constructed for upland, saddle, and floodplain communities are flattened in the horizontal direction, which indicates that their diversity indices exceed their evenness indices. The shape of multidimensional icon plots constructed for these communities is similar to the shape of averaged multidimensional icon plots constructed for subarctic landscapes in the northern part of Central Siberia (Litvinov, 2004) and open landscapes of Western Siberia (Litvinov, 2008); however, compared to the latter, it is more elongated in the direction of Simpson’s diversity index, which gives greater weight to common species.
The above-noted similarity with communities of open landscapes can be somewhat explained by afforestation features in the studied localities: low canopy density on the upland and in the saddle and the semiopen floodplain habitat.
Multidimensional icon plots constructed for the slope community are elongated along the vertical axis in the direction of Simpson’s evenness index; their shape is similar to averaged multidimensional icon plots constructed for forest communities of Western Siberia (Litvinov, 2008) and mountain taiga communities of the Altai Mountains (Litvinov, 2004). This is consistent with the forest vegetation structure on the slope where canopy density amounts to 70% (Table 1).
The shape of the averaged multidimensional icon plot is consistent with shapes of multidimensional icon plots constructed for a number of subarctic communities in the northern part of Central Siberia (Litvinov, 2004); overall, it is similar to averaged multidimensional icon plots constructed for small mammal communities studied by the authors in the western part of the north-taiga zone of Western Siberia (Malaya Sos’va State Nature Reserve and Kunovatskii Regional Wildlife Sanctuary) and in the mountain tundra of the Polar Urals (Lake Khadatayoganlor); however, it is more harmonious both in diversity and evenness (Fig. 2).

Map of multidimensional icon plots constructed for small mammal communities inhabiting high-latitude regions of Western Siberia.
Compared to small mammal communities inhabiting a forest–tundra landscape in the vicinity of the Tazovsky rural locality (upper reaches of the Syambota-Yakha River) and a north-taiga landscape in the foothills of the Polar Urals (Nyrdvomenshor Stream floodplain, Sob River basin), the averaged multidimensional icon plot constructed for the small mammal community of the Nadym Hills is less elongated along the axis of the Shannon evenness index (J), reflecting the evenness of species with low abundance values, but it is more balanced along all other axes (E, H, and D).
Multidimensional icon plots constructed for all small mammal communities of the Nadym Hills are symmetrical along one or both axes. This, along with their similarity to multidimensional icon plots constructed for communities inhabiting natural high-latitude and high-mountain landscapes of Western Siberia and adjacent areas, indicates their undisturbed dominance structure, the decisive effect of natural factors on the small mammal population of the studied biogeocoenoses, and the relative stability of the communities under study.
DISCUSSION
In the course of the census of small mammals conducted in the Nadym Hills in July 2021–2022, 5 species out of the 16 (17 species taking into account E. sibiricus) inhabiting the central part of the north-taiga subzone of Western Siberia were caught, including four species (Cl. rutilus, S. caecutiens, S. araneus, and A. oeconomus) out of the nine that form the core of small mammal communities throughout the north-taiga subzone (Starikov and Vartapetov, 2021). The absence of the remaining 12 small mammal species in the catches can be explained by their ecological and/or biotopic preferences and, accordingly, by the heterogeneity of their spatial distribution within the subzone; for some species, this can be explained by their uneven numerical distribution in the latitudinal and longitudinal directions, as well as the phase of low population size amid the universally insignificant abundance of these species.
All the recorded vole and shrew species were described as part of Pleistocene faunas in the northern part of Western Siberia (Smirnov et al., 1986). E. sibiricus, a typical species inhabiting modern north-taiga Siberian pine and pine forests (Ravkin et al., 1980; Mlekopitayushchie…, 2007), apparently penetrated into the northern regions of the West Siberian Plain later, in the course of the formation of the taiga–forest theriocomplex, a derivative from the tundra–steppe and forest–steppe faunal complexes, in the second half of the Holocene (Smirnov et al., 1986). Siberian and transpalaearctic species predominate among the recorded species, which is consistent with the geographical location of the study area and the history of faunas.
The dominance estimates are consistent with the data (Starikov and Vartapetov, 2021) indicating the universal and joint predominance of Cl. rutilus and S. caecutiens throughout the entire north-taiga subzone of Western Siberia with the maximum proportion of Cl. rutilus in the center of the subzone. However, contrary to the data collected by B.P. Starikov and L.G. Vartapetov (2021), in our study, S. araneus was recorded as a common species only on the upland, and it is not among the dominants.
The spatio–biotopic and numerical distributions of individual small mammal species by Nadym catena localities correspond to their species-specific ecological and biological features. The composition, structure, and parameters of small mammal communities change along the catena following changes in geosystem characteristics: afforestation, wetting, plant species richness, vegetation structure, and microtopography.
Low diversity and sustainability indices in small mammal communities inhabiting the Nadym Hills are determined by their harsh natural and climatic conditions, which is confirmed by the results of landscape and ecological studies (the presence of areas where perennially frozen rocks occur close to the surface, cryogenic heaving, areas with excessive moistening, low species diversity of higher vascular plants; the numerical predominance of a few plant species; etc.). Relatively higher values of diversity indices and the composite well-being index in the floodplain small mammal community correlate with the greater species and structural diversity of floodplain vegetation and a milder microclimate in this part of the landscape. Relatively higher resistant sustainability and, conversely, lower resilient and overall sustainability in communities inhabiting upper parts of the catena can be somewhat explained by the impact of fires.
The average multidimensional icon plot constructed for the entire small mammal community inhabiting all localities of the Nadym catena is symmetrical along both axes and more harmonious in diversity and evenness than multidimensional icon plots constructed for communities inhabiting its individual localities, which can be explained as follows: being an integral system, the landscape is balanced better than its local geosystems at the intralandscape level. The multidimensional icon plot constructed for the small mammal community of the Nadym Hills is more symmetrical and balanced in diversity and evenness when compared to multidimensional icon plots constructed for communities inhabiting western parts of the northern taiga, forest–tundra, and mountain tundra. This is consistent with the known zonal and provincial heat supply, moisture, and biodiversity distribution patterns in Western Siberia (Parfenova, 2004; Ravkin and Bogomolova, 2016, 2018).
CONCLUSIONS
The species composition and population structure of small mammals inhabiting the Nadym Hills correspond to the zonal–subzonal and provincial location of these hills, the formation history of the taiga faunal complex, the composition of biotopes, and their proportions within the study area.
Patterns determining changes in the composition, population structure, and parameters of small mammal communities inhabiting the Nadym Hills along the slope vector (i.e., from the upland to the floodplain) are consistent with changes in a number of other geosystem characteristics.
In all catena localities and throughout the entire study area, small mammal communities feature a generally undisturbed information structure typical for subarctic landscapes, which indicates their sustainability.
Despite traces of fires noted in the soil structure and their probable impact on the stability of small mammal communities in the upper catena localities, the predominance of anthropogenically neutral species in the studied communities and their low anthropogenic adaptation indicate the absence of a significant economic anthropogenic impact on the studied landscape.
REFERENCES
Beruchashvili, N.L. and Zhuchkova, V.K., Metody kompleksnykh fiziko-geograficheskikh issledovanii (Methods of Complex Physical and Geographical Research), Moscow: Mosk. Gos. Univ., 1997.
Bigon, M., Harper, J., and Townsend, C., Ecology: Individuals, Populations and Communities, Sunderland: Sinauer, 1986.
Bobretsov, A.V., Populyatsionnaya ekologiya melkikh mlekopitayushchikh ravninnykh i gornykh landshaftov Severo-Vostoka evropeiskoi chasti Rossii (Population Ecology of Small Mammals of the Plain and Mountain Landscapes of the Northeast of European Russia), Moscow: KMK, 2016.
Brown, J.H., Macroecology, Chicago: Univ. Chicago Press, 1995.
Chernykh, D.V., Zolotov, D.V., Biryukov, R.Yu., Kolesnikov, R.A., and Pechkin, A.S., Catenary differentiation of the Nadym Hills geosystems (Yamal-Nenets Autonomous District), Nauchn. Vestn. Yamalo-Nentskogo Avtonomnogo Okruga, 2022, vol. 3, no. 216, pp. 80–96.
Chibilyov, A.A., Parshina, V.P., and Pavkejchik, V.M., Landscape catenas as regional models for studying biodiversity, in Geneticheskie rastitel’nye resursy Rossii i sopredel’nykh gosudarstv (Genetic Plant Resources of Russia and NeighbouringCountries), Orenburg: Dimur, 1999, p. 59.
Ferronsky, I.V., About collection of papers “Modern global change of environment”, Vodn. Resur., 2013, vol. 40, no. 6, p. 633.
Gashev, S.N., Mlekopitayushchie v sisteme ekologicheskogo monitoringa (na primere Tyumenskoi oblasti) (Mammals in the System of Ecological Monitoring: The Example of Tyumen Oblast), Tyumen: Tumensk. Gos. Univ., 2000.
Gashev, S.N., RF Patent 2011615336, 2011.
Gashev, S.N., Soroking, N.V., and Khritan’ko, O.A., RF Patent 2013620056, 2013.
Gataullin, V.N., The Western Yamal – Marresale formation – deltaic deposits of the Pra-Ob, Byull. Kom. Izuch. Chetvertichn. Perioda, Akad. Nauk SSSR, 1991, no. 60, pp. 53–61.
Gosudarstvennaya geologicheskaya karta Rossiiskoi Federatsii. Tret’e pokolenie. Q-43 (Novyi Urengoi) 1 : 1 000 000. Karta pliotsen-chetvertichnykh obrazovanii (State Geological Map of the Russian Federation, Third Generation. Q-43 (Novy Urengoy) 1 : 1 000 000. Map of Pliocene-Quaternary Formations), VSEGEI, 2015.
Karaseva, E.V., Telitsyna, A.Yu., and Zhigalsky, O.A., Metody izucheniya gryzunov v polevykh usloviyakh (Methods for the Study of Rodents in the Field), Moscow: LCI, 2008.
Kryštufek, B., Tesakov, A.S., Lebedev, V.S., Bannikova, A.A., Abramson, N.I., and Shenbrot, G., Back to the future: The proper name for red-backed voles is Clethrionomys Tilesius and not Myodes Pallas, Mammalia, 2019, vol. 84, no. 2, pp. 214–217.
Lisovsky, A.A., Sheftel, B.I., Savelyev, A.P., Ermakov, O.A., Kozlov, Yu.A., Smirnov, D.G., Staheev, V.V., and Glazov, D.M., Mammals of Russia: List of types and applied aspects, in Sbornik trudov Zoologicheskogo muzeya Moskovskogo Gosudarstvennogo Universiteta (Collection of Works of the Zoological Museum of Moscow State University), Moscow: KMK, 2019, vol. 56.
Litvinov, Ju.N., Influence of factors of various nature on the diversity of small mammal communities, Usp. Sovrem. Biol., 2004, vol. 124, no. 6, pp. 612–621.
Litviniov, Ju.N., Microprocesses of communities’ evolution (small mammal communities case study), Vestn. Irkutsk. Gos. S-kh. Akad., 2008, no. 30, pp. 29–46.
Magurran, A.E., Ecological Diversity and Its Measurement, Princeton Univ. Press, 1988.
Mergelov, N.S., Post-pirogenic transformation of soils and soil carbon stocks in sub-tundra woodlands of Kolyma Lowland: A cascading effect and feedbacks, Izv. Ross. Akad. Nauk, Ser. Geogr., 2015, no. 3, pp. 129–140.
Mlekopitayushchie Polyarnogo Urala (Mammals of the Polar Urals), Berdyukina, K.I., Ed., Yekaterinburg: Ural. Gos. Univ., 2007.
Mordkovich, V.G., Shatokhina, N.G., and Titlyanova, A.A., Stepnye kateny (Steppe Catenas), Novosibirsk: Nauka, 1985.
Moskalenko, N.G., Cryogenic landscape changes in the West Siberian northern taiga in the conditions of climate change and human-induced disturbances, Kriosfera Zemli, 2012, vol. XVI, no. 2, pp. 38–42.
Mokhov, I.I., Eliseev, A.V., and Guryanov, V.V., Model estimates of global and regional climate changes in the Holocene, Dokl. Earth Sci., 2020, vol. 490, no. 1, pp. 27–32.
Parfenova, E.I., Tchebakova, N.M., and Vlasenko, V.I., Relationships between climatic factors and biodiversity of vegetation cover in Central Siberia at different levels of organization, Contemp. Probl. Ecol., 2004, vol. 11, no. 5, pp. 725–734.
Pavlinov, I.Ya. and Lissovsky, A.A., Mlekopitayushchie Rossii: sistematiko-geograficheskii spravochnik (The Mammals of Russia: A Taxonomic and Geographic Reference), Moscow: KMK, 2012.
Pavlov, A.V. and Malkov, G.V., Dynamic of permafrost zone of Russia under changing climate conditions, Prir. Protsessy Din. Geosist., 2010, no. 5, pp. 44–51.
Pesenko, Yu.A., Printsipy i metody kolichestvennogo analiza v faunisticheskikh issledovaniyakh (Principles and Methods of Quantitative Analysis in Faunal Research), Moscow: Nauka, 1982.
Ravkin, Yu.S. and Bogomolova, I.N., Ecological organization of spatial-typological diversity of ornithocomplexes of the West-Siberian plain, Vestn. Tomsk. Gos. Univ., Biol., 2016, vol. 4, no. 36, pp. 147–163.
Ravkin, Yu.S. and Bogomolova, I.N., Environmental organization of spatial-typological diversity of the communities of terrestrial vertebrates of the West-Siberian plain, Byull. Mosk. O-va Ispyt. Prir., Otd. Biol., 2018, vol. 123, no. 1, pp. 3–14.
Ravkin, Yu.S. and Livanov, S.G., Faktornaya zoogeografiya: printsipy, metody i teoreticheskie predstavleniya (Factor Zoogeography: Principles, Methods, and Theoretical Concepts), Novosibirsk: Nauka, 2008.
Ravkin, Yu.S. and Luk’yanova, I.V., Geografiya pozvonochnykh yuzhnoi taigi Zapadnoi Sibiri (Geography of Vertebrates of Southern Taiga of Western Siberia), Novosibirsk: Nauka, 1976.
Ravkin, Yu.S., Vartapetov, L.G., Fomin, B.N., and Ravkin, E.S., Peculiarities of Siberian chipmunk distribution in Western and Middle Siberia forest zone, in Fauna Sibiri (Fauna of Siberia), Yudin, B.S., Ed., Novosibirsk, 1980, pp. 148–157.
Shvarts, S.S., Smirnov, V.S., and Dobrinsky, L.N., The method of morphophysiological indicators in the ecology of terrestrial vertebrates, in Trudy Instituta ekologii rastenii i zhivotnykh (Proceedings of the Institute of Plant and Animal Ecology), Sverdlovsk: Akad. Nauk SSSR, 1968, vol. 58.
Smirnov, N.G., Bolshakov, V.N., and Borodin, A.V., Pleistotsenovye gryzuny Severa Zapadnoi Sibiri (Pleistocene Rodents of the Northern Part of Western Siberia), Moscow: Nauka, 1986
Starikov, V.P. and Vartapetov, L.G., Geographic ecological analysis of small mammals of the northern taiga of Western Siberia, Contemp. Probl. Ecol., 2021, vol. 14, no. 1, pp. 49–61.
Tishkov, A.A., The “Arctic Vector” in the preservation of terrestrial ecosystems and biodiversity, Arktika: Ekol. Ekon., 2012, vol. 2, no. 6, pp. 28–43.
Vinogradov, B.S. and Gromov, I.M., Gryzuny fauny SSSR (Rodents of the USSR Fauna), Moscow: Akad. Nauk SSSR, 1952.
Volpert, Y.L. and Shadrina, E.G., Melkie mlekopitayushchie severo-vostoka Sibiri (Small Mammals of the North-East Siberia), Novisibirsk: Nauka, 2002.
Yudin, B.S., Nasekomoyadnye mlekopitayushchie Sibiri (Insectivorous Mammals of Siberia), Novosibirsk: Nauka, 1989.
Funding
This study was supported by the Government of the Yamalo-Nenets Autonomous Okrug as part of the project “Modern Climatic Changes and Their Impact on the Landscape Structure in the Yamalo-Nenets Autonomous Okrug.” The original catenary landscape differentiation model was developed as part of the State Task of the Institute for Water and Environmental Problems, Siberian Branch, Russian Academy of Sciences, project no. FUFZ-2021-0007.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest. The authors declare that they have no conflicts of interest.
Statement on the welfare of animals. Animal-related experiments were approved by the Institutional Animal Ethical Committee on Biomedical Ethics of Siberian Federal University, Russia (Approval Code: State Standard of Russian Federation No. 33215-2014; Approval Date: 2016-07-01).
Additional information
Translated by L. Emeliyanov
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Levykh, A.Y., Chernykh, D.V., Zolotov, D.V. et al. Small Mammal Communities of the Nadym Hills and Changes They Undergo along a Landscape Catena. Contemp. Probl. Ecol. 16, 763–775 (2023). https://doi.org/10.1134/S1995425523060148
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1995425523060148