
Abstract
The differences in morphometric features of the canopy-gap structure of the three dominant forest types in the Valuevsky Forest Park were investigated with high-resolution and detailed-resolution remote sensing data. The forest-community groups (deciduous forest with a predominance of lime, deciduous forest with a predominance of birch or aspen, and coniferous forest with a predominance of spruce or pine) were classified according to the random forest method based on Sentinel-2/MSI multispectral satellite images. The better classificaton accuracy was 0.96 (k = 0.88). The Sentinel-2 data were used to create a layer of segments: spectrally homogeneous forest parcels. The forest gaps were obtained via cluster analysis with Resurs-P1 Geoton panchromatic data and visual interpretation of the clusters. Eight morphometric parameters were calculated for each gap. The differences were analyzed at the segment level (the Mann–Whitney U-test) and for all gap sets of each forest-community group (the Kruskal–Wallis H-test). The highest U-test values for the average values of morphometric features at the level of forest-community segments were obtained for the gap area (U = 24), gap perimeter (U = 19.3), gap-shape complexity index (U = 19.0), and the ratio of the perimeter to the gap area (U = 18.7). The highest values of the H-test at the level of individual gaps were calculated for the fractal dimension of the gap (H = 2229.2), the ratio of the perimeter to gap area (H = 2064.9), and the gap area (H = 1718.4). Analysis of the calculated and published data made it possible to find the possible reasons for the differences in the gap structure and gap parameters of coniferous, small-leaved, and lime communities of the model territory.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Forest ecosystems that develop spontaneously in the absence of catastrophic external influences are characterized by a mosaic structure represented by “renewal gaps” at different stages of development (Schliemann, Bockheim, 2011; Muscolo et al., 2014). “Gaps” refer to the gaps in the canopy of an adult stand (treefall gaps) a result of the death of one or more large trees. The gap mosaic of an adult stand determines the spatial heterogeneity of the forest phytocoenosis, which is associated with many important ecological structures and processes at different spatial scales. Gaps play a key role in reforestation, maintaining the taxonomic and structural biodiversity of forest communities (Shirokov, 2005). The appearance of gaps is found in 40- to 60-year-old stands, when trees of early successive species begin to die off (Smirnova et al., 2001). Once formed, the gaps do not remain static; they exist at various stages of “filling.” In the consideration of the entire cycle of gap development, it is appropriate to use the term “age parcel” proposed by N.V. Dylis (1978) from the moment of their appearance to the decay of a new generation of trees. Tall grasses and shrub species predominate in the first 15 years in the gaps in coniferous-deciduous forests of the European part of Russia; after the undergrowth canopy closes, they are replaced by forest nemoral and boreal plant species. The original structure of the lower layers is restored 35–40 years after the gap was formed (Shirokov, 2005).
In this work, we consider the gaps of the first developmental stages, from the moment of formation to the closing (gap closure). L.E. Tyrrell and T.R. Crow (1994) gave three criteria for gap closure: trees in a gap are one-half to two-thirds of the height of the surrounding canopy; trees in the gap have a diameter (at chest level) of more than 25 cm; the canopy is so dense that the original gap is difficult to identify.
The upper and lower limits of the gap size are another important parameter that must be considered in the examination of the gap structure of the upper canopy of the stand. By definition, gaps are small breaks in the canopy that form when one or more trees die off. J. McCarthy (2001) considers it necessary to distinguish the concepts of “gap” and “patch.” This is necessary, because areas formed by large-scale catastrophic disturbances (fires, insect outbreaks, and massive windfalls) are very different from small gaps in terms of the microclimate, regeneration time, illumination, accessibility to light-demanding flora, and other parameters. The lower limit for gap size is determined by the area from the fall of a single tree, which usually ranges from 50 to 200 m2 (Drobyshev, 1999; McCarthy, 2001).
Various ground-based methods are traditionally used for spatial analysis of the gap structure: (1) transects (Shirokov, 2005); (2) dendroecological analysis (Hobi et al., 2015); and (3) photography with a hemispherical camera (Nyamgeroh et al., 2018). However, ground-based methods require a significant investment of time and labor to carry out measurements and to prepare schemes for the spatial arrangement of gaps in the stand canopy (Hobi et al., 2015; Nyamgeroh et al., 2018). Therefore, their application is limited to small areas (up to several hectares).
The potential use of aerospace survey data to map and study the gap structure of forests has been studied in a number of works (Runkle, 1992; Fox et al., 2000; Hobi et al., 2015; Barton et al., 2017; Bagaram et al., 2018; Nyamgeroh et al., 2018; White et al., 2018). J.R. Runkle (1992) studied the potential mapping tree canopy gaps with aerial imagery data. T.J. Fox (2000) compared the accuracy of gap maps generated from a ground survey of the forest canopy with the results of visual interpretation of aerial photographs with a high spatial resolution (1 : 15 000). The aerial maps were shown to be more accurate, since they had a much lower omission rate (4.7%) for gap mapping than ground surveys (25.6%). Although aerial photography allows more accurate identification of gaps in the stand canopy, it is not possible to describe in detail the characteristics of vegetation in the gaps, which require ground-based methods.
A separate issue is the influence of the trees' own shadows on the accuracy of the determination of the boundaries and areas of the upper canopy gaps. Due to differences in tree heights, some crowns are shaded and are included in the gaps during mapping (Zielewska-Büttner et al., 2016; Barton et al., 2017; Nyamgeroh et al., 2018). B.B. Nyamgeroh et al. (2018) showed that the identification of gaps from images (Object Based Image Analysis (OBIA) and Image Texture Based Analysis (ITBA)) overestimates their area (especially for small gaps) in comparison with ground-based measurements. The gaps are detected worst of all in the canopy of broad-leaved species.
The advantage of satellite measurements is that they make it possible to study the gap structure of forests in hard-to-reach regions and, with repeated surveys of the same territories, to analyze the dynamics of the gap area and, accordingly, changes in the species-age structure of forests (Hobi et al., 2015; Nyamgeroh et al., 2018). The gap structure of the tree canopy differs in forest communities of different types and successional status. This fact can be used as an indirect decryption sign for their mapping. M.L. Hobi et al. (2015) used stereo pairs of WorldView-2 satellite images to study the parameters of the canopy gaps of old-growth beech forests of the Ukrainian Carpathians. Satellite images showed that the gap size in the canopy of a beech forest does not exceed 200 m2 and that the proportion of disturbances caused by hurricane winds and snow breaks is less than 1% of the study area. Verification of the satellite maps of the canopy gaps based on ground-based data proved to be challenging, since not all plots could be accurately spatially referenced during the ground survey. On the whole, however, the estimates of the gap sizes provided by ground-based and remote sensing methods are in good agreement. Thus, the published data analysis shows the complexity of the task and the prospects for the development and application of remote sensing methods to map the gap structure of forests and to monitor them in large forest areas.
The purpose of this investigation is to assess the differences of the parameters of the gap structure in different groups of forest communities based on satellite images with a detailed resolution. The following tasks were set: (1) to assess the informative value of the spectral channels and to classify the main groups of forest communities with Sentinel-2b/MSI satellite data (10–20 m); (2) to obtain a gap mask for the stand canopy with the Resurs-P1/Geoton panchromatic image (0.8 m); and (3) to calculate the morphometric features of the gap structure of the canopy and to statistically evaluate the differences in the values of morphometric features for groups of forest communities.
EXPERIMENTAL
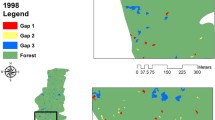
The studies were carried out on the model territory of the Valuevsky Forest Park (Novomoskovsky administrative district of Moscow) (Fig. 1). The total study area is 1156.33 ha (Publichnaya…, 2020). The Valuevsky Forest Park lies within the Aprelevsko–Kuntsevsky landscape, where moraine plains are the dominants (absolute heights of 190–210 m). The soil cover is predominated by sod-podzolic soils on cover loam. The parent rock is characterized by a medium-heavy, loamy granulometric composition (Akkumulyatsiya…, 2018). According to the taxonomic materials of the last forest inventory, forests are represented by broad-leaved (predominated by lime) and broad-leaved-spruce communities aged 60–110 years (Materialy lesoustroistva…, 2005). Secondary, small-leaved forests at different stages of postfelling succession are very widespread. Spruce and pine forest plantations occupy insignificant areas. The paper discusses three dominant groups of forest communities: (1) a deciduous forest with dominant lime; (2) a deciduous forest with birch or aspen as the dominants; (3) a coniferous forest with spruce or pine as the dominants.

Scheme of forest stands of the Valuevsky Forest Park by dominant species. 1, location of ground plots in 2016–2019. Forest formations: 2, Picea abies; 3, Betula pendula and Populus tremula; 4, Tilia cordata; 5, Pinus sylvestris; 6, other species.
The forest-community groups were selected via analysis of the materials from the latest forest inventory and data from selective ground geobotanical and taxonomic surveys.
Multi-seasonal multispectral images of the Sentinel-2 satellites, which launched in 2015 and 2017 as part of the Copernicus scientific space program of the European Commission, were used to assess the spatial distribution of forest communities in Valuevsky Forest Park. The multispectral instrument (MSI) of each satellite allows imaging of the Earth’s surface with a wide swath of 290 km with a reimaging period of 5 days in 13 channels of visible, near, and middle infrared (IR) ranges at various spatial resolutions (10, 20, and 60 m) (Sentinel-2…, 2020).
A time series of cloudless Sentinel-2 images with a Level-2A processing (Level-2A Algorithm Overview, 2020) for 2017, 2018, and 2019 was collected for the research. Table 1 shows the parameters of five channels and the characteristics of the ranges of values (minimum, maximum, average) of the spectral brightness of the forest-park vegetation in the indicated periods of the survey.
A geometric processing was carried out with the standard image processing algorithm “resampling” with the bicubic interpolation function (ERDAS Imagine…, 2014) for spectral channels with a spatial resolution of 20 m to obtain a uniform spatial resolution of all spectral channels.
A panchromatic image of the Resurs-P1 satellite with a spatial resolution of about 1 m was used for spatial identification of the gaps in the tree canopy. Resurs-P1 is a satellite with onboard optical instruments designed to monitor natural and anthropogenic objects of the Earth (Cosmic…, 2020).
In addition to satellite information, digital forest inventory data were prepared based on forest inventory materials in 2005. In the period from 2016 to 2019, ground-based surveys of the territory were carried out, 53 relevés were carried out on 20 × 20 m plots, and 15 tree taxation was carried out on 50 × 50 m plots (Fig. 1).
Figure 2 shows diagram of the sequence of research stages. The set of standards for the classification of forest-community groups was based on the materials of the forest inventory. An allotment became a candidate for standards if the stand contained seven or more units of a certain tree species.

Scheme of research stages.
When the proportion of species in the stand was less than the threshold determined in the study (seven units), the species of one group (pine and spruce, birch and aspen) were summed and compared again with the threshold value. The control over the compliance of the allotments with their taxonomic characteristics was carried out via visual analysis of synthesized satellite images and data from ground-based sites. To obtain spectrally homogeneous standards within the blocks, multiseason Sentinel-2 multispectral images were segmented with the Full Lambda Schedule (FLS) method (Redding et al., 1999) in the ERDAS Imagine satellite image processing program. The segments were further used for the statistical analysis of morphometric characters within their boundaries. On the one hand, this step made it possible to minimize the influence of errors in the classification of forest-community groups based on Sentinel-2 images, and on the other hand, it enabled assessment of the level of distinguishability of the morphometric trait values averaged per segment in relation to their initial values at the level of a separate gap.
Reference sampling was used to assess the informativeness of multispectral channels (features) and to adjust the classification model with “random-forest” training (Breiman, 2001). The choice of the “random-forest” algorithm is explained by its higher forests classification accuracy as compared to other methods of thematic image processing (Maxwell et al., 2018).
The results of the classification of forest-community groups were checked based on data from ground-based surveys. The dominant group of forest communities was determined for each site. The sites were then spatially combined with a thematic map of forest communities, and pixels with classes that fell within the sites were extracted. The accuracy of the thematic map of forest communities was assessed based on an analysis of the error matrix with an estimate of the Cohen’s kappa index (k) (Kramer,1975).
As part of the work, we take a minimum gap size equal to 20 m2, which is consistent with the average size of spruce and linden crowns in the Moscow region (15–27 m2) (Abaturov and Melankholin, 2004). Due to the absence of large disturbance areas in the study area (more than 1 ha) (McCarthy, 2001), we did not establish an upper limit for the gap size.
At the next stage of the research, cluster analysis of Resurs-P1/Geoton satellite images with detailed spatial resolution was carried out to identify the gap structure of the tree canopy of the model territory. The gap clusters include pixels with low brightness values that correspond not only to the stand gaps but also to the shaded areas of the tree crowns of the upper and subordinate tiers. Since the use of threshold methods is impossible due to the high variation in the gap brightness values for forest communities, we used an unmanaged, redundant classification based on the K-means method (Mirkes, 2011), followed by visual interpretation of the classes for each forest-community group separately and with the combination of groups into two classes, “gap” and “forest.”
The raster layers of segments and the masks of tree-canopy gaps were then converted into vector format with GIS and spatially intersected with the PostGIS extension functions st_intersects (https://postgis. net/docs/ST_Intersects.html) and st_within (https:// postgis.net/docs/ST_Within.html). The following morphometric characteristics were calculated for the vector layer of gaps: the area (S), the gap perimeter (P), their ratio (P : S), the circularity (1), and a number of fractal indices (2)–(5), namely (Getzin et al., 2012):
where CIRC is the gap circylarity, S is the gap square, and P is the gap perimeter;
where GSCI is the index of the gap-shape complex, S is the gap square, and P is the gap perimeter;
where PFD is the gap fractal dimension, S is the gap square, and P is the gap perimeter;
where FDI is the gap fractal dimension index, S is the gap square, and P is the gap perimeter; and
where FD is the fractal dimension, S is the gap square, and P is the gap perimeter.
The nonparametric Kruskal−Wallis H-test (Kruskal and Wallis, 1952) and its parametric analog, the Mann−Whitney U-test (Mann and Whitney, 1947), were used to calculate the differences in the values of the morphometric characters of the three forest-community groups. The morphometric characters were analyzed based on two data sets. The H-test was used at the level of individual gaps belonging to forest-community groups. In this case, each polygon in the vector layer was considered a separate gap belonging to one of the forest-community groups. The U-test was used at the segment level. The second test was carried out for a sample of averaged and weighted values of morphometric characters within the boundaries of a segment belonging to one of three forest-community groups.
RESULTS AND DISCUSSION
Analysis of the informational value of the Sentinel-2/MSI spectral channels showed that an increase in their number affects the results of the classification of forest communities. However, the contribution varies and depends on the phenological period of the forest survey and the spectral range (wavelength). The largest contribution to the recognition of three forest-community groups of 35 spectral channels (features) obtained in different growing seasons is made by the mid-IR ranges B11 (1565–1655 nm) for May, September, and December 2017, and B12 (2100–2280 nm) for August 2017 and May 2018. Next in importance are the near-infrared range B8, which was obtained by shooting in May and December 2018, and the green B3 and red B4 ranges for April 2017 and October 2018, respectively. The classification of forest-community groups with the “random forest” algorithm was carried out according to three sets of features, the 15, 21, and 35 spectral ranges.
The integral classification accuracy of forest communities with 15 features was 0.80 (kappa k = 0.64), 0.89 for 21 features (k = 0.84), and for 0.96 for 35 features (k = 0.88). Figure 3a shows the best result of the classification of forest-community groups according to Sentinel-2 satellite images.

Satellite image processing results: (a) map of forest-community groups with segment boundaries; (b) forest gaps with selection boundaries. 1, with a predominance of lime; 2, small-leaved; 3, with a predominance of conifers.
Classification with the Resurs-P1/Geoton panchromatic image resulted in 200 classes, of which classes with low values of the image pixels were visually combined into a group of classes characterizing the “gap.” All other classes have been assigned to the “canopy” group. The resulting binary gap mask was used to study the differences in the morphometric parameters of the gap structure of three forest-community groups (Fig. 3b).
The use of two tests for the testing of differences in morphometric features made it possible to identify the most informative of them for both levels (Table 2). The highest values of the H-test at the level of individual gaps of forest communities are the gap fractal dimension (PFD) (H = 2229.2), the ratio of the perimeter to the gap area (P : S) (H = 2064.9), and the gap area (S) (H = 1718.4).
The highest U-test values for the mean values of morphometric traits at the level of forest community segments are obtained for S (U = 24.0), P (U = 19.3), the gap-shape complex index (GSCI) (U = 19.0), and P : S (U = 18.7). Thus, P : S and S retain their differences in the forest-community groups even when they are averaged at the segment level. Statistical analysis also showed that the average gap area in coniferous stands and the proportion of the gap area in a segment exceeds the same values for small-leaved and broad-leaved species (Table 3, Fig. 4).

Distribution of the number of segments of coniferous, small-leaved, and broad-leaved (lime) communities by the mean value of the gap area (a) and the proportion of gap area in the segment (b). 1, small-leaved; 2, lime; 3, spruce.
Analysis of the published data shows that the gap structure of the canopy of forest communities is determined by the simultaneous processes of the formation and closing of gaps (Kenderes et al., 2009). The gap-structure parameters are specific for different types of forest communities and depend on many factors that can be combined into two groups corresponding to the aforementioned dynamic processes. The first group includes factors associated with the rate of death (mortality) of trees: the age and size of trees, the wind rate, various pathogens, and the position in the landscape. The second group combines factors that determine the rate of gap closure, such as lateral growth, preliminary undergrowth, etc.
S.E. Dahir and C.G. Lorimer (1996) studied the gap structure in deciduous forests at different developmental stages (young, mature, and old-aged) in the northern United States, establishing the dependence of gap size on the age and size of trees. The average gap area in young forests was four times smaller, and the proportion of gaps in the total plantation area was two times lower than that in old-aged forests. Similar results were obtained for Douglas fir (pseudo-tree) forests (Spies and Franklin, 1989). Various pathogens have been shown to affect the gap dynamics (Kneeshaw and Bergeron, 1998; Pham et al., 2004; Messier et al., 2005). The windblownness of trees, which depends on the species (Kenderes et al., 2009), the tree age, and the landscape position, is an important factor determining the intensity of gap formation (Mirin, 2010).
The rate at which gaps are closed is primarily determined by their size. Small gaps (the diameter of which does not exceed the height of the surrounding trees) close much faster than large ones, usually as a result of the lateral growth of tree branches (Ulanova, 2000; McCarthy, 2001; Kucbel et al., 2010). The lateral growth rate of broadleaf trees is estimated at 4–26 cm per year (Runkle, 1985, cited in McCarthy, 2001). However, conifers with monopodial growth have a limited capacity to fill the vacated space through flexible branching changes (Kuuluvainen, 1994; Pham et al., 2004), so gaps in coniferous forests can be more durable. The rate of gap closure can be influenced by a well-developed, subordinate (second) sublayer of the stand, where the trees are able to fill quickly the open areas that form in the canopy (Kenderes et al., 2009).
In our study, the largest values of the average gap size and the proportion of gap area in a segment were obtained for the group of coniferous communities. This result can be explained by several factors: the coniferous forests in the study area are the oldest; the spruce and pine trees reach the highest height (over 30 m); spruce communities were severely affected by the outbreak of the bark beetle in 2010–2014, and spruce with the shallow root system is more susceptible to windblows than deciduous trees. The age of small-leaved stands (80–100 years) is approaching the longevity limit of birch and aspen. Many trees have stem rot, which determines their windiness and wind breaking. The loss of trees in the upper layer of small leaved communities is quite active, but the gap-formation process in these communities is not as pronounced, because a high second sublayer of spruce and, to a lesser extent, lime and maple. Lime communities are characterized by the smallest values of the average size and proportion of gaps due to the capacity of lindens for lateral crown growth, their greater durability as compared to small-leaved species, and their lower wind susceptibility due to the relatively low tree height (up to 25 m) and deep root system.
CONCLUSIONS
Research has been carried out to assess the differences in the parameters of the gap structure of three forest-community groups in the Valuevsky Forest Park. To complete this task, a map of forest communities was created based on 35 spectral features of Sentinel-2/MSI multi-season images (10–20 m). The Sentinel-2 average IR channels for May, August, September, and December 2017 and May 2018 showed the most informational value for the classification process of forest-community groups. A mask was created for the stand canopy gaps via cluster analysis and visual interpretation of a Resurs-P1/Geoton panchromatic image (0.8 m), and eight morphometric features were calculated based on the published data. The values of the area, the perimeter, and their ratios, as well as the fractal dimension of the gap, demonstrate statistically significant differences for each of the three groups at the level of individual gaps of forest communities. The area and perimeter of the gap, their ratio, and the complex index of the gap shape are statistically significant features for the average values of morphometric features at the segment level. Differences were found in the values of the average gap area and its share in the segment for coniferous and deciduous communities. This is explained by the different light regimes affecting the restoration of undergrowth in communities, as well as the structure of the crowns of coniferous and deciduous species, the latter of which are able to occupy vacated space due to the active growth of lateral shoots.
REFERENCES
Abaturov, A.V. and Melankholin, P.N., Estestvennaya dinamika lesa na postoyannykh probnykh ploshchadyakh v Podmoskov’e (Natural Dynamics of Forest on Permanent Test Plots in Moscow Region), Tula: Grif i K, 2004.
Akkumulyatsiya ugleroda v lesnykh pochvakh i suktsessionnyi status lesov (Carbon Accumulation in Forest Soils and Succession Status of Forests), Lukina, N.V., Ed., Moscow: KMK, 2018.
Bagaram, M.B., Giuliarelli, D., Chirici, G., Giannetti, F., and Barbati, A., UAV remote sensing for biodiversity monitoring: are forest canopy gaps good covariates? Remote Sens., 2018, vol. 10, p. 1397.
Barton, I., Király, G., Czimber, K., Hollaus, M., and Pfeifer, N., Treefall gap mapping using Sentinel-2 images, Forests, 2017, vol. 8, no. 11, p. 426.
Breiman, L. Random forests, Mach. Learn., 2001, vol. 45, no. 1, pp. 5–32.
Cramér, H., Mathematical Methods of Statistics, Princeton: Princeton Univ. Press, 1946.
Dahir, S.E. and Lorimer, C.G., Variation in canopy gap formation among developmental stages of northern hardwood stands, Can. J. For. Res., 1996, vol. 26, no. 10, pp. 1875–1892.
Drobyshev, I.V., Regeneration of Norway spruce in canopy gaps in Sphagnum–Myrtillus old-growth forests, For. Ecol. Manage., 1999, vol. 115, no. 1, pp. 71–83.
Dylis, N.V., Osnovy biogeotsenologii (Fundamentals of Biogeocenology), Moscow: Nauka, 1978.
ERDAS IMAGINE 2014 product description. https://ru. scribd.com/document/254309118/ERDAS-IMAGINE-2014-Product-Description-sflb. Cited April 23, 2020.
Fox, T.J., Knutson, M.G., and Hines, R.K., Mapping forest canopy gaps using air-photo interpretation and ground surveys, Wildl. Soc. Bull., 2000, vol. 28, no. 4, pp. 882–889.
Getzin, S., Wiegand, K., and Schöning, I., Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles, Methods Ecol. Evol., 2012, vol. 3, no. 2, pp. 397–404.
Hobi, M.L., Ginzler, C., Commarmot, B., and Bugmann, H., Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing, Ecosphere, 2015, vol. 6, no. 5, pp. 1–15.
Kenderes, K., Král, K., Vrška, T., and Standovár, T., Natural gap dynamics in a Central European mixed beech—spruce—fir old-growth forest, Ecoscience, 2009, vol. 16, pp. 39–47.
Kramer, G., Matematicheskie metody statistiki (Mathematical Methods of Statistics), Moscow: Mir, 1975
Kneeshaw, D. and Bergeron, Y., Canopy gap characteristics and tree replacement in the southeastern boreal forest, Ecology, 1998, vol. 79, no. 3, pp. 783–794.
Kruskal, W.H. and Wallis, W.A., Use of ranks in one-criterion variance analysis, J. Am. Stat. Assoc., 1952, vol. 47, no. 260, pp. 583–621.
Kucbel, S., Jaloviar, P., Saniga, M., Vencurik, J., and Klimaš, V., Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians, Eur. J. For. Res., 2010, vol. 129, pp. 249–259.
Kuuluvainen, T., Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: a review, Ann. Zool. Fenn., 1994, vol. 31, no. 1, pp. 35–51.
Level-2A algorithm overview. https://earth.esa.int/ web/sentinel/technical-guides/sentinel-2-msi/level-2a/ algorithm. Cited April 23, 2020.
Mann, H.B. and Whitney, D.R., On a test of whether one of two random variables is stochastically larger than the other, Ann. Math. Stat., 1947, no. 18, pp. 50–60.
Materialy lesoustroistva leskhoza Eksperimental’nyi Moskvoretskii, Taksatsionnoe opisanie (po sostoyaniyu na 01.01.05 goda) (Forest Inventory Materials of the Moskvoretskii Experimental Forestry, Taxation Description (by January 1, 2005)), Moscow, 2005.
Maxwell, A.E., Warner, T.A., and Fang, F., Implementation of machine-learning classification in remote sensing: an applied review, Int. J. Remote Sens., 2018, vol. 39, no. 9, pp. 2784–2817.
McCarthy, J., Gap dynamics of forest trees: a review with particular attention to boreal forests, Environ. Rev., 2001, vol. 9, no. 1, pp. 1–59.
Messier, J., Kneeshaw, D., Bouchard, M., and de Römer, A., A comparison of gap characteristics in mixed wood old-growth forests in eastern and western Quebec, Can. J. For. Res., 2005, vol. 35, pp. 2510–2514.
Mirin, D.M., Reasons and consequences of high windfalls in spruce forests, Uch. Zap. Ross. Gos. Gidrometeorol. Univ., 2010, no. 13, pp. 111–120.
Mirkes, E.M., K-means and K-methods applet, University of Leicester, 2011. http://www.math.le.ac.uk/people/ ag153/homepage/KmeansKmedoids/Kmeans_Kmedoids. html. Cited January 28, 2021.
Muscolo, A., Bagnato, S., Sidari, M., and Mercurio, R., A review of the roles of forest canopy gaps, J. For. Res., 2014, vol. 25, no. 4, pp. 725−736.
Nyamgeroh, B.B., Groen, T.A., Weir, M.J.C., Dimov, P., and Zlatanov, T., Detection of forest canopy gaps from very high resolution aerial images, Ecol. Indic., 2018, vol. 95, pp. 629–636.
Public cadastral map of Moscow oblast. https://публичная-кадастровая-карта.рф/московская-область/. Cited January 21, 2020.
Pham, A.T., De Grandpré, L., Gauthier, S., and Bergeron, Y., Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec, Can. J. For. Res., 2004, vol. 34, pp. 353–364.
Redding, N.J., Crisp, D.J., Tang, D.H., and Newsam, G.N., An efficient algorithm for Mumford-Shah segmentation and its application to SAR imagery, Proc. Fifth Conf. on Digital Image Computing: Techniques and Applications (DICTA-99), Perth, 1999, pp. 35–41.
Runkle, J.R., Guidelines and Sample Protocol for Sampling Forest Gaps: General Technical Report PNW-GTR-283, Portland, OR: US Dep. Agric., 1992.
Schliemann, S.A. and Bockheim, J.G., Methods for studying treefall gaps: a review, For. Ecol. Manage., 2011, vol. 261, pp. 1143–1151.
Sentinel-2 MSI instrument. https://sentinels.copernicus.eu/ web/sentinel/technical-guides/sentinel-2-msi/msi-instrument. Cited January 29, 2020.
Shirokov, A.I., Use of the parcel analysis to assess the structural biodiversity of forest communities, Lesovedenie, 2005, no. 1, pp. 19–27.
Smirnova, O.V., Bobrovskii, M.V., and Khanina, L.G., Assessment and forecast of succession processes in forest cenoses based on demographic methods, Byull. Mosk. O-va. Ispyt. Prir., Otd. Biol., 2001, vol. 106, no. 5, pp. 25–33.
Spacecrafts of the Resurs-P type (in operation). http://www. ntsomz.ru/ks_dzz/satellites/resurs_p. Cited April 23, 2020.
Spies, T.A. and Franklin, J.F., Gap characteristics and vegetation response in coniferous forests of the Pacific Northwest, Ecology, 1989, vol. 70, pp. 543–545.
Tyrrell, L.E. and Crow, T.R., Structural characteristics of old-growth hemlock–hardwood forests in relation to age, Ecology, 1994, vol. 75, pp. 370–386.
Ulanova, N.G., The effects of windthrow on forests at different spatial scales: a review, For. Ecol. Manage., 2000, vol. 135, pp. 155–167.
White, J.C., Tompalski, P., Coops, N.C., and Wulder, M.A., Comparison of airborne laser scanning and digital stereo imagery for characterizing forest canopy gaps in coastal temperate rainforests, Remote Sens. Environ., 2018, vol. 208, pp. 1–14.
Zielewska-Büttner, K., Adler, P., Ehmann, M., and Braunisch, V., Automated detection of forest gaps in spruce dominated stands using canopy height models derived from stereo aerial imagery, Remote Sens., 2016, vol. 8, no. 3, p. 175.
Funding
The research was carried out within the framework of the state contract with Center for Forest Ecology and Productivity, Russian Academy of Sciences, no. АААА-А18-118052590019-7, and the field studies were financed by the Russian Science Foundation, project no. 16-17-10284.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest. This article does not.
Additional information
Translated by M. Shulskaya
Rights and permissions
About this article

Cite this article
Komarov, A.V., Ershov, D.V. & Tikhonova, E.V. Informativeness of the Spectral and Morphometric Characteristics of the Canopy-Gap Structure Based on Remote Sensing Data. Contemp. Probl. Ecol. 14, 733–742 (2021). https://doi.org/10.1134/S1995425521070076
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1995425521070076