
Abstract
Through comparing three survey reports of Caohai since 1986 once a decade in the past thirty years, we confirm that some physical and chemical factors directly (e.g. a decrease of water salinity, pH variability) have provided opportunities for the invasion of Procambarus clarkii (Girard, 1852) in Caohai (a wetland situated in Guizhou, South-West China) in 2010. In addition, some physical and chemical factors (e.g. an increase of organic oxygen consumption and total nitrogen) reflect the richness of herbivorous food (vascular plants residues) for this exotic crayfish. Furthermore, we suggest that the successful invasion of exotic crayfish is related to human activity and the presence of Anatidae waterflows. Direct and indirect effects of crayfish invasion on Caohai have been evaluated by comparing data between before 2010 and after 2010. Although it is an omnivorous species, eating animal food is more conducive to its growth and reproduction. Invasion of red swamp crayfish had a negative effect on submerged plants, Annelida, aquatic insects and amphibians. By contrast, increases in this exotic crayfish provides a richer food source for Ciconiifornis birds. Further research is needed to solve crayfish invasion thoroughly.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
Biotic invasions refer to the not exclusively human-driven phenomena of non-native species (non-indigenous in taxonomic categories and geographic origins) being transferred to new, often distant, ranges where their descendants proliferate, spread, and establish a self-sustained population that gradually develops into a dominant population. The consequence of biotic invasions could be deemed more detrimental generally, with altering dominant species in a community, a physical feature of ecosystem, nutrient cycling, plant productivity, and decaying of biodiversity (Mack et al., 2000). The adverse effects of biotic invasion have been ranked among the most important drivers of biodiversity losses (Sala et al., 2000) and species extinctions in freshwater ecosystems (Foden et al., 2008). Invasion of aquatic alien species is the second largest threat to the reduction or extinction of freshwater fish populations (Foden et al., 2008).
Procambarus clarkii (the red swamp crayfish) is native to Eastern North America, but has been introduced worldwide because of its economic importance and value as a food source (Barbaresi et al., 2000). P. clarkii was introduced from Japan to Nanjing, China in 1929, from where it has spread to almost every province and city. This species has established wild populations in some rivers, ponds, ditches, paddy fields and other water bodies. Multiple features determine the high invasiveness of P. clarkii: firstly, it has a habit of burrowing, as main prerequisite for the invasiveness of non-indigenous species, for feeding, reproductive, and defensive purposes. The burrows allow crayfish to withstand environmental extremes, such as high temperatures and dehydration, and protects them from predators. In addition, they usually live in the cracks of masonry and caves, and use grasses as natural shelters. P. clarkii can adapt to a wide range of environment, with the suitable pH range is 7.5–8.5, the suitable dissolved oxygen amount is not less than 2 mg/L, and the salinity is less than 10 g/L, and temperature from 5°C to 38°. This invasive species can thus withstand a wide range of physical, chemical, and biological conditions, demonstrating great ecological plasticity (Matteo et al., 2011). Further, it is a generalist omnivores and opportunistic crayfish, feeding on vegetal and animal detritus, submerged vegetation, molluscs, amphibians, fish and insect larvae (Matteo et al., 2011; Gherardi et al., 2007). Some adult individuals even show cannibalism. Thirdly, it has high fertility, with P. clarkii can produce more than 700 eggs based on female size, and the complex mother-offspring relationships enhance descendant survival increasing the populations growth (Gherardi, 2006; Aquiloni and Gherardi, 2008). Finally, it has strong resistance to plague of Aphanomyces astaci (Souty-Grosset et al., 2006; Gherardi and Panov, 2009) and parasite of Paragonimus genus (Gherardi, 2006; Gherardi and Panov, 2009), and able to accumulate and transfer heavy metals, pesticides and toxins of cyanobacteria to its consumers, therefore it can withstand extreme environmental conditions and outcompete native species.
Given the P. clarkii is a generalist omnivore (Parkyn et al., 2001), the effect of introducing it into aquatic systems is not restricted to displacement of the autochthonous crayfish but also extends to the rest of the trophic chain levels (Donato et al., 2018). Various types of impact have been documented in response to the introduction of P. clarkii. Invasion of this species is a known cause for the decline of several native freshwater species, such as competitive removal of autochthonous fish species, the disappearance of amphibians due to predation, declines in macroinvertebrates (Correia, 2002), and destruction of the aquatic plant cover (Rodríguez et al., 2003). By contrast, P. clarkii has also become a new and rich food source for a number of vertebrate predators, such as fish and birds, and may thus provide positive change for some native species (Ilhéu et al., 2007).
It is not known exactly when and how the P. clarkii entered the Caohai Lake, but we learned, from local residents and experts, that crayfish first appeared in Caohai in approximately 2010. Since that time, it has widespread throughout whole Caohai conservation reserve, becoming the dominant species in some sewers, rivers and river channels. This study reviews the existing three times comprehensive survey reports on Caohai wetland, comparing the survey data from its early recovery stage in 1985 (Xiang, 1986), before the introduction of crayfish in 2005 (Zhang et al., 2007) with later one after crayfish successful invasion in 2017, we outline hypotheses on the causes and consequences of this crayfish invasion, which could help to estimate its potential future distribution area and identify effective management practices to contain its spread. At the same time, this information may provide evidence of the effects of predation by alien crayfish on the trophic web of Caohai wetland.
Study area. Caohai wetland is known for its high conservation value and is important for several populations of migratory birds, especially Grus nigricollis, plateau cranes endemic to China. Caohai National Nature Reserve is located in the southwest of Weining Yi and Hui Autonomous County, Guizhou Province, China. It is a typical Karst shallow lake (1980 hm2; maximum depth 5.0 m), and its geographic coordinates are from 26°47′35″ N to 26°52′10″ N, and from 104°9′23″ E to 104°20′10″ E. The Caohai was traditionally used as an irrigation pool for agriculture, the inhabitants of the area have traditionally used it for uncontrolled hunting of birds, amphibians and fish. In the early 1970s, in order to get a little more land, a devastating human intervention “released water to make land” caused the plateau freshwater lake ecosystem that has been running for centuries to collapse and disintegrate. In years, this intervention resulted in ecological imbalances, abnormal climate, bird migration, soil desertification, as well as crop diseases and insect’s plague. These consequences of this change demonstrate the important function of this lake ecosystem in maintaining and regulating the local ecological environment. Not only rich in biological resources, but also a large number of paleontological fossils and human cultural relics, it has long attracted the attention of domestic and international scholars. The Guizhou government decided to restore the Caohai area and has been regularly monitored since 1980 onwards.
MATERIALS AND METHODS
Physical and chemical water data from Caohai, and the aquatic plant and fauna inventories in periods before and after exotic crayfish introduction, were extracted from the two comprehensive survey reports on Caohai wetland in 1985 and 2005 (Xiang, 1986; Zhang et al., 2007), as well as the later survey in 2017. Due to the fact that these studies partially overlap, the inventory with the largest record of species was used to establish the number of species affected and the decrease. Due to the lack of data on Zoobenthos after 2007, the related data of Zoobenthos after crayfish invasion was extracted from the published article (Chen et al., 2015). However, due to their short survey time and small survey scope, the statistically obtained species of Zoobenthos were not complete.
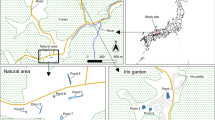
Due to the lack of data referring to the P. clarkii for Caohai wetland, we estimated the Caohai crayfish densities in October of 2018 and 2019. Due to the high conservation value of Caohai, the local government has banned vessels from entering Caohai core waters. Therefore, six sites of marginal water (Jiangjia Wan, Yangguan Mountain, Baijia Zui, Dengjia Yard, Louluo Moutain and Gujia Dixia) (Fig. 1) were selected within the Caohai. Three sampling stations were selected within each site, making a total of eighteen sampling stations. At each station, three non-baited fyke traps were set up for a period of 24 h. The abundance and proportion of capture were estimated on the basis of the presence and absence of crayfish in each fyke-trap (Meineri et al., 2014).

Map of the sampling sites in the Caohai Lake. The pushpin shows the location of six sampling sites (Yangguan Mountain, Louluo Mountain, Gujia Dixia, Baijia Zui, Dengjia Yard and Jiangjia Wan).
Statistical analysis. The captured abundance and proportion of P. clarkii and other fauna were calculated as means ± standard deviation (SD), and evaluated by one-way analysis of variance (ANOVA) using GraphPad Prism 5 version software. Significance in the mean values was set at p < 0.05.
RESULTS
Abundance and proportion of P. clarkii and other fauna in Caohai. The abundance of fauna ranged from five to eight species in one trap. The abundance of fauna at Jiangjia Wan reached the peak (Fig. 2). From Palaemon sinensis, Pseudoperilampus lighti, Procambarus clarkii, Pelteobagrus fulvidraco, Bellamya aeruginosa, Rhodeus ocellatus, Ctenogobius giurinus, Pseudorasbora parva, Carassius auratus, Parafossarulus striatulus, to Misgurnus anguillicaudatus, the abundance was decreased. The proportion of P. sinensis at all the sites was significantly higher than other species (Fig. 3). P. clarkii relative proportion varied, but on the whole the proportion of crayfish at sampling sites near Weining Country (Figs. 3a, 3c, 3d) was significantly higher than other sampling sites (Figs. 3a, 3e, 3f), human activities are presumed to play a major role in influencing the crayfish abundance and proportion, and enrich the local species.

Captured species abundance in one trap at different sampling sites. For each species, the upper and lower transverse lines represent the maximum and lowest abundance of the captured species in one site, respectively. The mean values of every species are plotted as a middle longer transverse line. (a) Jiangjia Wan; (b) Yangguan Mountain; (c) Baijia Zui; (d) Dengjia Yard; (e) Louluo Mountain; (f) Gujia Dixia. Here and Fig. 3 letters (a–e) in common indicate no significant difference (p < 0.05).

Captured species proportion in the one trap at different sampling sites. (a) Jiangjia Wan; (b) Yangguan Mountain; (c) Baijia Zui; (d) Dengjia Yard; (e) Louluo Mountain; (f) Gujia Dixia. Vertical segment on the top of each bar represents the standard error.
Changes in physical and chemical variables. Water and air temperature both were significantly increased from 1986 to 2007, may resulting in the decrease of water dissolved oxygen and oxygen saturation from 2007 to 2017. After the arrival of this crayfish, air temperature decreased by 41% and water temperature by 23%, respectively. The same trend was also reflected in pH, silicon and total ferrum. There was not significant change of pH before and after crayfish arrived, but the water body is always in a weak alkaline state, which is conducive to the survival of crayfish. To the opposite, the water depth and total nitrogen first decreased and then increased. These physical and chemical characterizations showed a wavy change. From 1986 to 2017, the organic oxygen consumption presents a continuous upward trend, whereas the total phosphorus and water salinity decline constantly, especially from 1986 to 2007, the salinity dropped by 67%. As necessary nutrient elements, Cl–, Mg2+ and Na+ also increased constantly, while Ca2+ and \({\text{SO}}_{4}^{{2 - }}\) showed the opposite trend (Suppl. Table S1).
Phytoplankton. Total biomass of phytoplankton decreased after crayfish invasion, although almost every species amount of phytoplankton increased. Before crayfish invasion, the ratio of Chlorophyta coverage and biomass were the highest of all phytoplankton. After the crayfish invasion, the ratio of Chlorophyta coverage decreased but still the highest (Suppl. Table S2). The ratio of biomass declined by 52% significantly, with the Pyrrophyta biomass reached the highest of all phytoplankton. Followed by, the ratio of coverage of Cyanophyta was decreased by 54%, but with biomass increasing by 56%. The ratio of coverage of Bacilariophyta increased by 96%, but with the ratio of biomass decreased by 69%. Density defined as the biomass divided by coverage, the density of Cyanophyta and Pyrrophyta increased by 80 and 68%, respectively. The others phytoplankton density decreased by 90–98%, except for a 46% reduction in Chlorophyta.
Zooplankton. After crayfish invasion, there were five genera and four species in total missing in Zooplankton, which were Trichoderma, Solanum, Phytophthora, Pyrus and Cucurbita, and the added genera were Amoeba, Epistylis, Vorticella and Euplotes. Rotifera missing genera are Argonotholca, Collotheea, Hexarthra, Squatinella and Ploesoma, and those added were Pompholyx, Proales, and Testudinella. As for Cladocera, the missing genera were Dadaya, Eurycerus and Polyphemus, those added were Bosminopsis, Chydorus, Rhynchotalona and Simocephalus. Copepoda missing genera were Eodiaptomus and Neodiaptomus, the added were Canthocamptus, Acanthocyclops and Limnoithona. Compared the survey between before and after crayfish invasion, the dominant species in Zooplankton are Arcella vulgaris and Lecane luna, there was no significant change for the composition of Zooplankton species from 2007 to 2017 (Suppl. Table S3).
Aquatic macrophytes. According to vegetation morphological characteristics and ecological habits, the aquatic macrophytes of Caohai are divided into emergent plants, floating-leaved plants, submerged plants and floating plants. After the crayfish invasion, the total biomass increased, with a 61% in fresh weight and a 52% in dry weight. And the amount of species had increased significantly, especially the emergent plants has increased by 31%, but the coverage ratio of different aquatic plants types almost decreased except emergent plants, with gone down from emergent, submerged, floating and floating-leaved plants (Table 1). Before and after the crayfish invasion, the changes in dominant species were mainly reflected in emergent and submerged plants, the added species were Phragmites australis and Typha angustifolia of emergent plants, and Potamogeton perfoliatus, Myriophyllum spicstum and Potamogeton pectinatus of submerged plants (Suppl. Table S4). The Sparganium stoloniferum of emergent plants and Trapa maximowiczii of floating-leaved plants, listed in 1986 survey report, were not appeared in the dominant species of 2007 and 2017 survey reports (Suppl. Table S4).
Zoobenthos. It shows that species amount had decreased, especially Annelida and Aquatic insect, but increased in Mollusc after this exotic crayfish invasion (Suppl. Table S5). In addition, the dominant species also changed, mainly in Gastropods and Crustacea. Before crayfish invasion, the dominant species including Cipangopaluding cathayensis, Radix lagotis, Neocaridina dentioulata, Pantala flavescens and Tendipus sp. There was the appearance of Sinodina gregoriana, but not in the dominant species list. After crayfish invasion, there were many Gastropods added in dominant species, including Parafossarulus striatulus, Bellamya aeruginosa, Lymnaea stagnalis, Bellamya purificata, Radix ovata and Tricula. As for Crustacea, the Caohai native species N. dentioulata had not been found, and S. gregoriana had become the dominant species. In addition, two exotic crustacean species P. sinensis and P. clarkii had been listed in the dominant species (Table 2).
Amphibian. The findings have shown that crayfish will prey on the eggs and larvae of amphibians, leading to a decline in quantity and species of amphibians (Kats and Ferrer, 2003; Cruz and Rebelo, 2005; Rodríguez et al., 2005). Before crayfish invasion, fourteen species of amphibians were found in Caohai Lake, but after appearance of crayfish, an alien species (Lithobates catesbeiana) of amphibian was found. Native species Tylototriton kwcichowensis and Psewdorana weiningensis have disappeared (Suppl. Table S6).
Birds. Among the composition of bird resources in Caohai, the water bird group is the most distinctive. The water birds, especially Anatidae, were the most abundant and speciose birds in Caohai. The Caohai’s bird resources can be classified into migrant, summer migrant, passing migrant, winter migrant, migrant status unclear and waterfowl species. Before the crayfish invasion, most species were migrants, while waterfowl is a little less. Following the P. clarkii invasion, however, waterfowl species increased significantly more than other species, and became the largest group birds in Caohai (Table 3). After 2010, although the dominant species changed significantly, only including the Anas Penelope, Anas strepera, Tadorna ferruginea, Fulica atra, Gallinula chloropus, Anser indicus, Grus nigricolis, Grus grus and Egretta garzetta, there increased 25 newly bird species (Table 4).
DISCUSSION
Here, we reviewed and compared the past three 10-year surveys of Caohai wetland to summarize and analyze the potential losses in biodiversity following the invasion of P. clarkii in this region. In Fig. 4, we attempt to summarize these findings and thereby lay a foundation for future monitoring this exotic species invasion. While we can summarize these correlative patterns in the environment and species composition, quantitative determination of the causal relationships among these factors are only possible through future research.

Outline of putative direct and indirect causal factors and consequences of exotic crayfish invasion in Caohai ecosystem. Red box indicates the increased biomass, species amount, or value of physical and chemical characterisation of Caohai water body after crayfish invasion, the green box stands for the decreased ones. “+” and “–” shows the positive and negative effect, respectively.
Considering crayfish habitat, our analysis showed that changes of physical conditions, such as salinity, pH and water temperature, may explain the invasion of crayfish successfully a decade ago. The pH (7.7–10.2) and water temperature (20–29°C) around 2010 were near optimal crayfish growth and reproduction. Huner and Barr study has demonstrated that P. clarkii adult individuals are found in water up to 10 g/L salinity and that reproduction is limited to water less than 5 g/L salinity (Huner and Barr, 1991). The recent study that traps were placed in water bodies that had a salinity level above 5 g/L captured with no crayfish, further confirms that low salinity can increase the occurrence, the abundance and the reproduction success of P. clarkii (Meineri et al., 2014). Caohai water salinity is not only less than this threshold limit (5 g/L) since record in 1986, but also constantly decreasing from 1986 to 2017. The crayfish abundance from sampling sites near the residence area is higher than others (Fig. 2). This may result from more domestic water pouring into the lake leading to lower salinity. It speculated that the increase in freshwater inputs and crayfish introduction from other places for unknown reasons lead to its invasion.
P. clarkii is a generalist and opportunistic omnivore (Parkyn et al., 2001), the effect of its introducing into aquatic system is not restricted to displacement of the autochthonous macroinvertebrates, such as the disappearance of N. dentioulata after its invasion (Table 2). But can also extends to the rest of the trophic chain levels through predating, grazing and burrowing. P. clarkii diet includes submerged macrophytes, invertebrates, algae and detritus. However, in preference experiments, it shown that crayfish prefer animal food over detritus or plants (Gherardi and Barbaresi, 2007). Animal protein is a chief substance promoting crayfish rapid growth, experimental studies show that crayfish survival and growth increase when fed animal food as compared to a diet based on plants or detritus (Gherardi and Barbaresi, 2007). In our study, we saw many remains of predated P. sinensis in containers during we captured and shipment of crayfish back to the lab, and our results (Figs. 2 and 3) showed that P. sinensis could be captured at every sampling site and its biomass and proportion were the highest. P. sinensis is another exotic crustacean and its body length is from 20–40 mm, preferring to live in freshwater lakes or ponds with aquatic plants, has become the dominant species in Caohai wetland. The data of relationship between P. sinensis and P. clarkii are consistent with the results given in the previously studies, with P. clarkii change their diet from plant food and detritus to an animal diet in relation to the availability of macroinvertebrates (Correia, 2002). It is uncertain which invaded the Caohai Lake earlier as for P. clarkii and P. sinensis, but given the abundance and wide distribution of P. sinensis providing high occurrence of crayfish large-scale expansion. Our analyzed data further demonstrated that this species is selecting its animal prey as the animal food is abundance, it prefers on the most profitable prey items available with more energy and less consumption, resulting in the decrease of aquatic insect and Crustacean, increase of Gastropods and Oligochaeta in dominant species of Zoobenthos (Table 2). The increase of Oligochaeta may be due to its living in sediment, P. clarkii must be exhausted from burrowing the sediment to obtain food. Gastropods are protected by their hard shells.
The impacts of exotic crayfish invasion also on the decrease of Amphibian through direct predation on its eggs and larvae, e.g. on Taricha torosa (Kats and Ferrer, 2003), decreasing its larvae activities (Cruz and Rebelo, 2005), preventing its successful mating and spawning, and destroying its spawning grounds by destroying the aquatic plants. Three native amphibian species disappeared, and one exotic amphibian species appeared after P. clarkii been introducing in Caohai wetland. Disappearance of these species may be due to the alternative predation of P. clarkii on eggs and larvae of amphibians, or disturb on its spawning and mating. Furthermore, it is the result of another exotic amphibian species invasion, e.g. L. catesbeianus. L. catesbeianus, native to the eastern United States, is considered one of the 100 most invasive species in the world (Lowe et al., 2000), it can threaten native amphibians through direct predation (Jankowski and Orchard, 2013), competition (Snow and Witmer, 2011), sexual interference (Pearl et al., 2005) and transmission of pathogens (Martel et al., 2013). Among L. catesbeianus food, P. clarkii is not only preyed upon by bullfrog, but also probably as bullfrog first source of food (Vannini et al., 2015). The phenomenon that the areas where P. clarkii and L. catesbeianus live in sympatry observed in Caohai also confirm this point. So another possible explanation for the decrease in amphibian species observed is related to the invasion of P. clarkii inducing an expansion of invasive L. catesbeianus and indirectly leading to the disappearance of native amphibian species in Caohai.
In addition to human activities, birds may be another vector for dispersal of P. clarkii in Caohai wetland except human activities. The characteristics of Caohai bird distribution before crayfish invasion (Tables 3 and 4) is consistent with the evidence given that Anatidae are of particular importance for the dispersal of exotic crayfish due to their abundance, widespread distribution, and the capacity for long-distance movements (Green et al., 2002). Furthermore, the evidence given in the study of P. clarkii was readily preyed upon by species of Ciconiiform birds (Correia, 2001) providing some explanations on the distribution traits of dominant bird species in Caohai after crayfish invasion (Table 4).
The impacts of P. clarkii invasion impacts on flora are less pronounced than those on frauna. Phytoplankton, as the primary producer, is the basis of the material cycle and energy flow in the entire aquatic ecosystem. Due to its respond quickly to changes in the nutrition status of the water body, phytoplankton is usually a common indicator for evaluating eutrophication. Silicon, the main component of diatom cells, is very important for the total production of organisms in water. It has shown that diatoms cannot reproduce below 0.13 mg/L. From 1986 to 2017, Caohai silicon content is always higher than 0.13 mg/L, so it will not likely affect diatom reproduction. Ferrum, an important trace element of phytoplankton, can promote the normal production of chlorophyll, and involved in respiration as the main component of animal heme, cytochrome oxidase, catalase and peroxidase. The optimal concentration of ferrum for phytoplankton growth is 0.14–1.47 mg/L. From 2007 to 2017, the ferrum content decreased from 0.5 mg/L to 0.06 mg/L, which is not conducive to the growth of phytoplankton. Phosphorus is an essential nutrient element for all phytoplankton. Carbohydrates and phosphoric acids (directly related to photosynthesis) form phosphate esters, which are the main cellular components and involved in energy transmission of respiration and fermentation process. That total phosphorus presented continuous decline from 1986 to 2017 maybe a factor to inducing the decrease of phytoplankton (Goldyn et al., 2007). Na+ mainly comes from human activities and is discharged into the water body as domestic sewage. Cl–, Ca2+, Mg2+ and \({\text{SO}}_{4}^{{2 - }}\) are necessary for phytoplankton metabolism. As the components of chlorophyll, Ca2+ has important effects on the synthesis and metabolism of proteins, the conversion of carbohydrates, and the absorption and transformation of nitrogen and phosphorus, and Mg2+ is mainly involved in the metabolism of carbohydrates. When there are more Ca2+ and Mg2+ in the water body, the organisms absorb them less. Therefore, the increase of Mg2+ in water body indicated that the amount of Mg2+ used by phytoplankton in the water is reduced, and it is speculated that the increase of Mg2+ is caused by the reduction of phytoplankton. However, as for Ca2+ decrease, it may be the result of the rose of pH, inducing the increase of \({\text{CO}}_{3}^{{2 - }}\), increasing likelihood of Ca2+ present as precipitates of CaCO3. Sulfur is an essential nutrient element for algae, which absorbs and utilizes \({\text{SO}}_{4}^{{2 - }}\) to meet the needs of sulfur. \({\text{SO}}_{4}^{{2 - }}\) is one of the main anions in natural waters, which is high in content and generally not lacking. In total, increase of Cl- and Na+, decrease of Ca2+, \({\text{SO}}_{4}^{{2 - }}\), total ferrum and total phosphorus, and the appearance of exotic crayfish may together have reduced phytoplankton abundance, which would contribute to the higher water transparency and [Mg2+].
Higher water transparency is beneficial to the growth of submerged plants. However, the compared data show that the submerged plants biomass seems stable before and after crayfish invasion. Thus, our analysis is consistent with many other studies have shown that even relatively low crayfish densities (<1 ind/m2) can completely remove submerged plants from shallow lakes (Rodríguez et al., 2003). However, success invasion of crayfish plays different consequences on macrophytes (Fernández-Aláez et al., 2002). Another significant change of aquatic macrophytes is the increase of emergent plants, one of the vascular plants (Table 4). Examination of stomach contents emphasizes indigestible residues, such as plant fragments and detritus, has confirmed that vascular plants residues is one of the herbaceous food sources for this species. The increase of emergent plants providing enough food for exotic crayfish, while the burrow behavior of crayfish boosts the inorganic nutrient resolve in favor of emergent plants growth. The amount of oxygen consumption by organisms (called organic oxygen consumption) in Caohai is mainly determined by the amount of aquatic vascular plant residues, which consumes a lot of oxygen when being decomposed by microorganisms. The more organic matter deposited on the lake bottom, the greater the oxygen consumption potential. The continuous upward trend of organic oxygen consumption strongly manifests the richness of aquatic vascular plant residues on the bottom, improving the fertility of the water body and facilitating the growth of aquatic macrophytes. The increase in total nitrogen after the occurrence of crayfish may be also the results of increasing in aquatic vascular plant residues.
In conclusion, some observed factors (e.g. decrease of water salinity, rich vascular plants residues, vast Anatidae waterflows and human activity) together likely facilitated the successful invasion of the red swamp crayfish in Caohai. Because of its omnivorous diet and burrowing behavior, P. clarkii invasion appears to have direct and indirect effects on the Caohai wetland ecosystem through a trophic cascade. Ultimately, the invasion of this species may be driving declines in Amphibians, Zoobenthos and aquatic macrophyte, and facilitating invasion of some Ciconiifornis bird species, and stablization of submerged plants (Fig. 4). In order to control the P. clarkii invasion more reasonably and effectively, however, the quantitative relationships among these factors and species is in urgent need of further study.
REFERENCES
Aquiloni, L. and Gherardi, F., Extended mother-offspring relationships in crayfish: the return behavior of juvenile Procambarus clarkii, Ethology, 2008, vol. 114, p. 946.
Barbaresi, S. and Gherardi, F., The invasion of the alien crayfish Procambarus clarkii in Europe, with particular reference to Italy, Biol. Invasions, 2000, vol. 2, p. 259.
Chen, F.Y., Liang, Z.Q., He, T.R., et al., Community structure of macrozoobenthos and biological water quality assessment in Caohai wetlands in Guizhou, Hubei Agric. Sci., 2015, vol. 54, p. 4446.
Correia, A.M., Seasonal and interspecific evaluation of predation by mammals and birds on the introduced red swamp crayfish Procambarus clarkii (Crustacea, Cambaridae) in a freshwater marsh (Portugal), J. Zool., 2001, vol. 255, p. 533.
Correia, A.M., Niche breadth and trophic diversity: feeding behavior of the red swamp crayfish (Procambarus clarkii) towards environmental availability of aquatic macroinvertebrates in a rice field (Portugal), Acta Oecol., 2002, vol. 23, p. 421.
Cruz, M.J. and Rebelo, R., Vulnerability of Southwest Iberian amphibians to an introduced crayfish, Procambarus clarkii, Amphibia-Reptilia, 2005, vol. 26, p. 293.
Donato, R., Rollandin, M., Favaro, F., et al., Habitat use and population structure of the invasive red swamp crayfish Procambarus clarkii (Girard, 1852) in a protected area in northern Italy, Knowl. Manage. Aquat. Ecosyst., 2018, vol. 419, p. 12.
Fernández-Aláez, M., Fernández-Aláez, C., and Rodríguez, S., Seasonal changes in biomass of charophytes in shallow lakes in the northwest of Spain, Aquat. Bot., 2002, vol. 72, p. 335.
Foden, W., Mace, G., Vié, J., et al., Wildlife in a Changing World: An Analysis of the 2008 IUCN Red List of Threatened Species, Gland: Int. Union Conserv. Nat., 2008.
Gherardi, F., Crayfish invading Europe: the case study of Procambarus clarkii, Mar. Freshwater Behav. Physiol., 2006, vol. 39, p. 175.
Gherardi, F. and Barbaresi, S., Feeding preferences of the invasive crayfish, Procambarus clarkii, Bull. Fr. Peche Piscicult., 2007, vol. 387, p. 7.
Gherardi, F. and Panov, V.E., Procambarus clarkii (Girard), red swamp crayfish/crawfish (Cambaridae, Crustacea), in Handbook of Alien Species in Europe, Dordrecht: Springer-Verlag, 2009.
Goldyn, R., Podsiadlowski, S., Kowalczewska-Madura, K., et al., Lake restoration using iron treatment: preliminary results, Proc. 12th World Lake Conf. (Taal 2007), Jaipur, 2007, p. 2237.
Green, A.J., Figuerola, J., and Sánchez, M.I., Implications of waterbird ecology for the dispersal of aquatic organisms, Acta Oecol., 2002, vol. 23, p. 177.
Huner, J.V. and Barr, J.E., Red Swamp Crawfish: Biology and Exploitation, Baton Rouge, LA: Louisiana State Univ., 1991.
Ilhéu, M., Bernardo, J.M., and Fernandes, S., Predation of invasive crayfish on aquatic vertebrates: the effect of Procambarus clarkii on fish assemblages in Mediterranean temporary streams, in Biological Invaders in Inland Waters: Profiles, Distribution, and Threats, Dordrecht: Springer-Verlag, 2007, p. 543.
Jankowski, K. and Orchard, S.A., Stomach contents from invasive American bullfrogs Rana catebeiana (=Lithobates catesbeianus) on southern Vancouver Island, British Columbia, Canada, NeoBiota, 2013, vol. 16, p. 17.
Kats, L.B. and Ferrer, R.P., Alien predators and amphibian declines: review of two decades of science and the transition to conservation, Diversity Distrib., 2003, vol. 9, p. 99.
Lowe, S.J., Browne, M., Boudjelas, S., and De Poorter, M., 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database, Auckland: IUCN/SSC Invasive Species Spec. Group. 2000.
Mack, R.N., Simberloff, D., Lonsdale, W.M., et al., Biotic invasions: causes, epidemiology, global consequences, and control, Ecol. Appl., 2000, vol. 10, no. 3, p. 689.
Martel, A., Adriaensen, C., Sharifian-Fard, M., et al., The novel “Candidatus Amphibiichlamydia ranarum” is highly prevalent in invasive exotic bullfrogs (Lithobates catesbeianus), Environ. Microbiol. Rep., 2013, vol. 5, p. 105.
Matteo, E.S., Raoul, M., Emilio, P.S., et al., Spatial autocorrelation and the analysis of invasion processes from distribution data: a study with the crayfish Procambarus clarkii, Biol. Invasions, 2011, vol. 13, p. 2147.
Meineri, E., Rodriguez-Perez, H., Hilaire, S., and Mesleard, F., Distribution and reproduction of Procambarus clarkii in relation to water management, salinity and habitat type in the Camargue, Aquat. Conserv.: Mar., Freshwater Ecosyst., 2014, vol. 24, p. 312.
Nyström, P., Axelsson, E., Sidenmark, J., and Brönmark, C., Crayfish predation on amphibian eggs and larvae, Amphibia-Reptilia, 1997, vol. 18, p. 217.
Parkyn, S.M., Collier, K.J., and Hicks, B.J., New Zealand stream crayfish: functional omnivores but trophic predators? Freshwater Biol., 2001, vol. 46, p. 641.
Pearl, C.A., Hayes, M.P., Haycock, R., et al., Observation of interspecific amplexus between Western North American ranid frogs and the introduced American bullfrog (Rana catesbeiana) and an hypothesis concerning breeding interference, Am. Midl. Nat., 2005, vol. 154, p. 126.
Rodríguez, C.F., Bécares, E., and Fernández-Aláez, M., Shift from clear to turbid phase in Lake Chozas (NW Spain) due to the introduction of American red swamp crayfish (Procambarus clarkii), Hydrobiologia, 2003, vol. 506, p. 421.
Rodríguez, C.F., Bécares, E., Fernández-aláez, M., and Fernández-aláez, C., Loss of diversity and degradation of wetlands as a result of introducing exotic crayfish, Biol. Invasions, 2005, vol. 7, p. 75.
Sala, O.E., Chapin, F.S., Armesto, J.J., et al., Biodiversity-global biodiversity scenarios for the year 2100, Science, 2000, vol. 287, p. 1770.
Snow, N.P. and Witmer, G., A field evaluation of a trap for invasive American bullfrogs, Pac. Conserv. Biol., 2011, vol. 17, p. 285.
Souty-Grosset, C., Holdich, D.M., Noel, P.Y., et al., Atlas of Crayfish in Europe, Paris: Mus. Natl. Hist. Nat., 2006, p. 190.
Vannini, A., Bruni, G., Cantini, M., et al., Updating the distribution of the American bullfrog Lithobates catesbeianus (Anura: Ranidae) in Tuscany (Central Italy), with a note on predatory interactions with red swamp crayfish Procambarus clarkii (Decapoda: Cambaridae), Ital. J. Zool., 2015, vol. 82, no. 3, p. 416.
Xiang, Y.H., Caohai Scientific Investigation Report, Guiyang: Guizhou People Press, 1986.
Zhang, H.H., Li, M. J., and Yao, S. L., Caohai Research, Guiyang: Guizhou Science and Technology Press, 2007.
ACKNOWLEDGMENTS
Thanks to the expert Huiming Chen from the Guizhou Institute of Biology for the classification and identification of the captured specimens. Thanks to the strong support from Jingcheng Ran, head of the Guizhou Wildlife Management Station, and the Guizhou Caohai National Nature Reserve Administration.
Funding
This Work was supported by grants from the Science and Technology Project of Guizhou ([2018]1054) and Guizhou High-level Innovative Talent Project ([2021]11).
This work was supported by the Key Discipline of Ecology of Guizhou Province (ZDXK[2013]11) and Guizhou Province Key laboratory of Ecological Protection and Restoration of Typical Plateau Wetlands ([2020]2002). We thank Heath A. MacMillan for constructive criticism on an earlier draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Supplementary Information
Rights and permissions
About this article

Cite this article
Qing Li, He, B., Chen, Y. et al. Factors Inducing the Crayfish Procambarus clarkii Invasion and Loss of Diversity in Caohai Wetland. Inland Water Biol 15, 446–457 (2022). https://doi.org/10.1134/S199508292204040X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S199508292204040X