
Abstract
Fibrinogen (FG) is a high molecular weight soluble plasma protein responsible for the formation of fibrin. Being an extremely vulnerable target for oxidants, FG undergoes post-translational modifications that disrupt the structure and function of the protein. Thermal denaturation of FG during its induced oxidation by hypochlorite and hydrogen peroxide is studied for the first time using differential scanning calorimetry (DSC). The D and E regions of FG are shown to undergo structural changes in the oxidized protein molecule. The thermodynamic parameters of denaturation—temperature and calorimetric enthalpy—are determined. The consequences of the effect of oxidative modification of FG on its thermal stability and disturbance of the structure of the D and E regions of the protein are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Fibrinogen (FG) is a high molecular weight (MW = 340 kDa) soluble plasma protein, which is mainly responsible for the formation of insoluble fibrin gel in the presence of thrombin. An FG molecule can be approximated by a rod-shaped particle 45 nm in length with the degree of asymmetry close to two [1]. As a homodimer, the FG molecule contains two sets of three nonidentical polypeptide chains, Aα, Bβ, and γ, which are linked together by 29 disulfide bonds. In turn, the polypeptide chains are composed of 610, 461, and 411 amino acid residues, respectively [2]. The FG molecule is divided into several structurally and functionally important regions: the central region E formed by the NH2-terminal sections of all three pairs of polypeptide chains; two peripheral regions D, consisting of COOH-terminal sections of polypeptide chains Bβ and γ, which form the so-called β- and γ-nodules, respectively; two areas BβN, which are composed of NH2-terminal sections of chains Bβ; and two regions of αC, including a significant part of the COOH-terminal regions of the chains Aα (amino acid residues 221–610) [1, 3]. We note that in order to provide the rod-like shape of the FG molecule required for the formation of long fibrin fibrils, regions E and D are significantly distant from each other and are held together by coiled-coil strands. In addition, each of the two αC regions is formed by αC-domains that are in contact with each other and with the central region E and covalently linked to the main part of the FG molecule by flexible αC-connectors [4, 5].
FG occupies a special place among other plasma proteins, due to at least three distinctive properties. First, being a key protein in blood coagulation, FG is simultaneously a marker of the acute phase of diseases [6–8]. Second, the structure of FG is composed of both typical globular domains and includes coiled-coil structures characteristic of fibrillar proteins [1]. Finally, the FG molecule exhibits the maximal vulnerability to oxidation in comparison with other blood plasma proteins [9]. The latter property is of particular importance, since the oxidation of FG in vivo during the development of oxidative stress in the body causes damage to the structure and function of the protein, contributing to the development of a wide range of diseases [10]. All of this is due to the extremely intensive and multidisciplinary studies of the FG structure and the mechanism of its damage.
It is known that the oxidation of proteins is accompanied by the modification of amino acid residues localized both on the surface and inside the protein globule. In the latter case, such modified residues tend to migrate to the surface of the protein. This causes a partial violation of the structure of the protein globule and an increase in its hydrophobicity. Since the oxidation of amino acid residues (with the exception of cysteine) in plasma proteins is an irreversible process, we can consider the irreversible process of denaturation of the protein microstructure. Since FG does not contain any free cysteine group, the oxidation of this protein is a completely irreversible process.
The method of differential scanning calorimetry (DSC) of a protein provides a fairly complete thermodynamic characteristic of the process of its thermal denaturation. However, understanding the energy effects related to the structural disturbances of a protein during its oxidation requires knowledge of oxidative sites in each of the individual structural regions. In recent years, DSC has become a new tool for characterizing the thermodynamic properties of basic serum/plasma proteins and establishing calorimetric markers in various diseases [11].
Using the DSC method, the phenomenon of a decrease in the resistance of many proteins, including albumin and myoglobin, to thermal denaturation under in vitro conditions during induced oxidation [12–14] or in vivo in various diseases related to oxidative stress [15, 16] was revealed.
Earlier, when studying the effect of the oxidative modification of ozonized FG on its thermal stability, a decrease in the enthalpy of denaturation for all structural regions of oxidized FG was shown, which is a consequence of the modification of its molecular structure during oxidation [17]. In this study, the DSC method is used to assess the effects of FG oxidation induced by hydrogen peroxide and hypochlorite on the stability of the structure of its individual regions to thermal denaturation. Together with the obtained results of mass spectrometry on the identification of oxidative sites in a protein during its treatment with the aforementioned oxidants [18, 19], this will allow a deeper understanding of the mechanism of damage to the structure and function of the modified protein.
EXPERIMENTAL
Fibrinogen was isolated from the citrated blood plasma of donors by glycine precipitation [20] and dissolved in a 50 mM phosphate buffer (0.15 M NaCl, pH 7.4). FG oxidation was induced by adding a solution of hydrogen peroxide (Sigma-Aldrich, United States) or sodium hypochlorite (Sigma-Aldrich, United States) [21, 22] at concentrations of 50, 150, and 300 μM. After FG oxidation, electrophoresis of the reduced protein samples and its covalently cross-linked polypeptide chains under the action of the activated coagulation factor XIII (FXIIIa) was performed [21]. Coomassie brilliant blue dye R-250 manufactured by Thermo Fisher Scientific (United States) was used to stain the protein gels.
The thermodynamic parameters of FG denaturation (6.5 mg/mL) in the absence and in the presence of oxidants at different concentrations were determined using a highly sensitive DSC on a DASM-4 microcalorimeter (Pushchino, Russia). The sample volume was 0.5 mL in a closed cell. The measurements were carried out in the temperature range 10–110°C at a constant pressure of 2.5 MPa and a heating rate of 1°C/min. The scale of excess heat capacity for each experiment was calibrated using the Joule–Lenz effect. A 50 mM phosphate buffer was used as the reference solution for the measurement (see above).
The average values of the thermodynamic parameters of denaturation were determined from at least three parallel measurements. The denaturation temperature value corresponded to the maximum of the heat capacity peak on the thermogram. The experimental denaturation enthalpy corresponded to the area under the peak of the excess heat capacity curve as a function of temperature. The molar enthalpy of denaturation (ΔdH) was calculated on 1 mol of FG (340 kDa).
RESULTS AND DISCUSSION
Figure 1 shows the results of electrophoresis of polypeptide chains of native and oxidized FG (either with hydrogen peroxide or hypochlorite), which indicate that, regardless of the chosen concentration of the oxidant, neither protein fragmentation nor the formation of covalent crosslinks of its chains was observed. In the presence of FXIIIa, fibrin polypeptide chains are involved in covalent crosslinking, which manifests itself in the formation of γ–γ dimers and α–α polymers [23, 24]. With an increase in the concentration of the oxidizing agent, the amount of α–α polymers and γ–γ dimers formed decreases, which is also indicated by an increase in the content of the initial Aα- and γ‑chains (Fig. 1). Obviously, this is a consequence of the oxidative modification of the structure of the FG molecule.

Electrophoresis of various FG samples: (a) electrophoresis of FG polypeptide chains in polyacrylamide gel (5%, concentrating gel; 12%, separating gel) in the presence of sodium dodecyl sulfate; (1) unoxidized FG; (2) oxidized with 50 μM H2O2 or HOCl/–OCl; (3) oxidized with 150 μM H2O2 or HOCl/–OCl; (4) oxidized with 300 μM H2O2 or HOCl/–OCl. (b) Polyacrylamide gel electrophoresis of FG crosslinking reaction products with factor XIIIa (4%, concentrating gel; 8%, separating gel) in the presence of sodium dodecyl sulfate: (5) the reaction products of FXIIIa with unoxidized FG; (6) FG, oxidized with 50 μM H2O2 or HOCl/–OCl; (7) 150 μM H2O2 or HOCl/–OCl; and (8) 300 μM H2O2 or HOCl/–OCl.
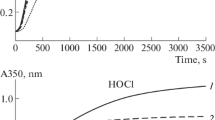
Figure 2 shows thermograms of FG denaturation in the absence and in the presence of sodium hypochlorite and hydrogen peroxide in various concentrations. The values of thermodynamic parameters of FG denaturation both in the absence and in the presence of oxidants (HOCl/–OCl and H2O2) are given in Table 1.

DSC thermograms of FG denaturation (6.5 mg/mL, 50 mM phosphate buffer, 0.15 M NaCl, pH 7.4) in the absence of (1) and in the presence of hypochlorite oxidants (curves 3–5) and hydrogen peroxide (6, 7) at their various concentrations: 50 μM (3, 6), 150 μM (4, 7), 300 μM (5); (2) thermogram of repeated scanning of FG solution (1).
From the DSC thermograms shown in Fig. 2, it can be seen that the melting curves of FG have two peaks: a low-temperature one in the region of 51 to 52°C and the high temperature in the region of 95°C. The presence of two peaks on the DSC thermogram is correlated with the published data [25]. The low-temperature peak is related to the unfolding of the peripheral regions D of the FG molecule, and the high-temperature peak is related to the unfolding of the central region E of the FG molecule. The process of protein denaturation under the studied conditions is irreversible for both transitions, which is reflected in the DSC thermogram of the repeated scanning of the FG solution (Fig. 2, curve 2), where there are no transitions at all. It should be noted that a reversible high-temperature transition of FG was observed in the 50 mM glycine buffer at pH 8.5 [26].
When FG is oxidized by sodium hypochlorite at a concentration of 50 μM, the values of the temperature and enthalpy of the low-temperature transition (i.e., for the D region of FG molecules) are close to the corresponding values for unoxidized FG. With an increase in the concentration of the oxidant to 300 μM, the low-temperature peak broadens, which is accompanied by a decrease in the temperature and enthalpy of the transition. The values of these parameters in the case of a high-temperature peak (i.e., for region E of FG molecules) decrease with an increase in the concentration of the oxidant. It is obvious that the oxidative modification of FG by sodium hypochlorite is accompanied by disturbances in the native structure of the protein, since its oxidized forms are more easily denatured.
When FG is oxidized with hydrogen peroxide at concentrations of 50 and 150 μM, the values of temperature and enthalpy of denaturation for the low-temperature transition decrease in comparison with the corresponding values for unoxidized FG. At the same time, the values of the thermodynamic parameters of denaturation for the low-temperature peak oxidized by hydrogen peroxide of FG are quite close to each other. The value of the enthalpy of denaturation for the high-temperature peak of FG oxidized by hydrogen peroxide decreases with the increasing concentration of the oxidizing agent. It should be noted that hydrogen peroxide at a concentration of 150 μM had a more significant effect on the structure of the central region E of the FG molecule than hypochlorite at the same concentrations, which is reflected in the value of the enthalpy of the high-temperature transition. Thus, the oxidative modification caused by the studied oxidants at these concentrations leads to damage of the native structure of FG.
Earlier in the work concerning the melting of ozonized FG [4], in the unoxidized protein, in addition to two main peaks, minor peaks with melting temperatures of 62.5 and 73.0°C were found, corresponding to the unfolding of the structure of the C- and N‑terminal subdomains of the αC-domain [4, 17, 27]. These peaks are absent on the thermograms (Fig. 2), which is explained in [4] and in this study by the different buffer composition.
It is known that the αC-region of human FG (amino acid residues 221–610) consists of a flexible NH2-terminal connector (221–391) and a compact COOH-terminal of the αC-domain (392–610), which can contain only some part of the regular structure [28]. The cooperative structure in the isolated αC-domain is inherently unstable and extremely sensitive to the concentration and composition of the buffer, and also has low thermal stability and can unfold even at room temperature [29]. It should be added that, according to an alternative point of view, αC-domains are “free-floating regions” devoid of any ordered structure [30], which is confirmed by the X-ray diffraction studies of intact chicken FG, in which αC-domains are not found on electron density maps [31]. Although it is believed that the αC domains of human FG form relatively compact structures [1], their ordered structure is still unidentified. Thus, the question of whether these regions contain a compact ordered structure remains controversial [32].
The studies performed have convincingly shown that the oxidative modification of FG causes rearrangements in the structure of the D and E regions of the protein, an indicator of which, according to the DSC data, is a decrease in its thermal stability. It was previously shown [18, 19] that FG, being a highly vulnerable target for hypochlorite- and peroxide-induced oxidation, undergoes significant modification, affecting multiple amino acid residues that belong to all three polypeptide chains of the protein and are localized in all structural regions of FG, including regions D and E. The electrophoresis data indicate the preservation of the integrity of the polypeptide chains of oxidized FG; however, the decrease in the content of α–α polymers and γ–γ dimers (Fig. 1) indicates a violation of the reactivity of the sites involved in the covalent crosslinking of chains.
As shown by us earlier [17], oxidation increases the hydrophobicity of FG. It is known that the oxidation of methionine residues in proteins induced by the reactive oxygen species leads to an increase in their surface hydrophobicity [33]. The mass spectrometry data indicate [18, 19] that the most common modification in FG during its treatment with HOCl/–OCl or H2O2 is the conversion of methionine to methionine sulfoxide (MetSO). Since the formation of MetSO is correlated to an increase in protein hydrophobicity [34], accompanied by partial denaturation of the protein globule and its dehydration [35], it is obvious that these effects cause a decrease in the thermal stability of FG, observed in calorimetric experiments.
Our data indicate a significant effect of oxidation on the thermodynamic characteristics of unfolding of the E region of FG, despite the fact that for both oxidants this region was least susceptible to posttranslational modifications [18, 19]. In this study, it is rather difficult to unambiguously explain the nature of this phenomenon. It can be assumed that this is due to the structural features of region E, which is a complex multidomain complex [3]. It is not yet clear how oxidation can affect the structure and stability of individual domains of region E. In addition, it is possible that due to the high conformational lability of both the FG molecule itself and its individual structures, this effect can be mediated, i.e., caused by oxidation and, as a consequence, damage to protein structures that do not belong to region E. For example, this can be caused by damage to coiled-coil structures (about 17 nm long) formed by the residues of all three, Aα, Bβ, and γ chains, which, on the one hand, according to the mass spectrometry data [18, 19], are actively involved in oxidative modification, and on the other hand, they can affect the structure of the latter, holding together the regions D and E. Of course, this assumption requires a special study.
CONCLUSIONS
In conclusion, we would like to note the followingpoints: DSC is a thermodynamic method for the direct measurements of thermal effects related to intra- and inter-molecular processes using diluted polymer solutions [36]. In accordance with the virial equation for the osmotic pressure of a polymer solution, the criterion for the ideality of a solution is the small value of product A2c compared to the molecular weight of the polymer, i.e., A2c \( \ll \) 1/M [35], where c and M are the concentration and molecular weight of the polymer and A2 is the second virial coefficient. The latter in the case of FG is vNA/2M 2 [37], where v is the excluded volume of the molecule and NA is the Avogadro number. In turn, quantity v can be calculated as v = 0.5πLD2, where L and D are the length and diameter of the rod-shaped molecule. If we take into account the values of the length and diameter of the FG molecule, 45 and 2 nm, respectively [2], we can obtain the value A2c, which is not more than a few percent of the value 1/M. This indicates that the FG solution is diluted. It should also be taken into account that quantity A2 was estimated for an unhydrated particle, while the FG molecule, like other soluble proteins, is covered with a hydration shell. However, the values of the hydrodynamic constants of FG (the coefficients of sedimentation, translational diffusion, intrinsic viscosity [38]) indicate that with an extremely high degree of asymmetry of the molecule, the degree of hydration should be extremely low in order to make significant corrections to the estimate of value A2.
REFERENCES
J. W. Weisel and R. I. Litvinov, Subcell. Biochem. 82, 405 (2017).
B. Blombäck, Thromb. Res. 83, 1 (1996).
L. Medved and J. W. Weisel, J. Thromb. Haemost. 7, 355 (2009).
R. I. Litvinov, S. Yakovlev, G. Tsurupa, et al., Biochemistry 46, 9133 (2007).
L. V. Medved, O. V. Gorkun, and P. L. Privalov, FEBS Lett. 160, 291 (1983).
C. Cray, Prog. Mol. Biol. Transl. Sci. 105, 113 (2012).
L. Zuliani-Alvarez and K. S. Midwood, Adv. Wound Care (New Rochelle) 4, 273 (2015).
M. A. Petersen, J. K. Ryu, and K. Akassoglou, Nat. Rev. Neurosci. 19, 283 (2018).
E. Shacter, J. A. Williams, M. Lim, et al., Free Radic. Biol. Med. 17, 429 (1994).
M. Martinez, J. W. Weisel, and H. Ischiropoulos, Free Radic. Biol. Med. 65, 411 (2013).
S. Todinova, S. Krumova, A. Danailova, et al., Eur. Biophys. J. 47, 549 (2018).
M. Anraku, K. Yamasaki, T. Maruyama, et al., Pharm. Res. 18, 632 (2001).
M. G. Gorobets, L. A. Wasserman, A. V. Bychkova, et al., Chem. Phys. 523, 34 (2019).
Y. Thiansilakul, S. Benjakul, and M. P. Benjakul, Food Chem. 124, 254 (2011).
M. Moezzi, I. Zapf, T. Fekecs, et al., J. Therm. Anal. Calorim. 123, 2037 (2016).
I. Zapf, M. Moezzi, T. Fekecs, et al., Therm. Anal. Calorim. 123, 2029 (2016).
M. G. Gorobets, L. A. Wasserman, A. V. Bychkova, M. L. Konstantinova, I. G. Plaschina, and M. A. Rosenfeld, Dokl. Biochem. Biophys. 480, 146 (2018).
L. V. Yurina, A. D. Vasilyeva, V. L. Kononenko, A. E. Bugrova, M. I. Indeykina, A. S. Kononikhin, E. N. Nikolaev, and M. A. Rosenfeld, Dokl. Biochem. Biophys. 492, 130 (2020).
L. V. Yurina, A. D. Vasilyeva, A. E. Bugrova, M. I. Indeykina, A. S. Kononikhin, E. N. Nikolaev, and M. A. Rosenfeld, Dokl. Biochem. Biophys. 484, 37 (2019).
N. J. White, Y. Wang, X. Fu, et al., Free Radic. Biol. Med. 96, 181 (2016).
A. D. Vasilyeva, L. V. Yurina, V. B. Leonova, D. Yu. Azarova, A. E. Bugrova, T. S. Konstantinova, M. I. Indeykina, A. S. Kononikhin, E. N. Nikolaev and M. A. Rosenfeld, Russ. J. Phys. Chem. B 14, 468 (2020).
A. N. Shchegolikhin, A. D. Vasilyeva, L. V. Yurina, and M. A. Rosenfeld, Russ. J. Phys. Chem. B 15, 123 (2021).
R. Chen and R. F. Doolittle, Biochemistry 10, 4487 (1971).
J. H. Sobel and M. A. Gawinowicz, J. Biol. Chem. 271, 19288 (1996).
P. L. Privalov and L. V. Medved, J. Mol. Biol. 159, 665 (1982).
Y. Chen, H. Mao, X. Zhang, et al., Int. J. Biol. Macromol. 26, 129 (1999).
J. W. Weisel and L. Medved, Ann. (N.Y.) Acad. Sci. 936, 312 (2001).
R. A. Burton, G. Tsurupa, L. Medved, et al., Biochemistry 45, 2257 (2006).
G. Tsurupa, L. Tsonev, and L. Medved, Biochemistry 41, 6449 (2002).
R. F. Doolittle, Ann. Rev. Biochem. 53, 195 (1984).
Z. Yang, J. M. Kollman, L. Pandi, et al., Biochemistry 40, 12515 (2001).
R. F. Doolittle, Blood Rev. 17, 33 (2003).
J. E. Zull, S. K. Smith, and R. Wiltshire, J. Biol. Chem. 265, 5671 (1990).
C. C. Chao, Y. S. Ma, and E. R. Stadtman, Proc. Natl. Acad. Sci. U. S. A. 94, 2969 (1997).
L. Wang, L. Li, H. Wang, et al., Biochem. J. 473, 4373 (2016).
P. L. Privalov and A. I. Dragan, Biophys. Chem. 126, 16 (2007).
Ch. Tenford, Physical Chemistry of Macromolecules (Wiley, New York, 1961).
M. A. Rozenfel’d, K. B. Gershkovich, D. V. Kuznetsov, et al., Mo. Biol. 20, 1098 (1986).
Funding
We used the equipment of the Center for Collective Use of the Institute of Biochemical Physics , Russian Academy of Sciences.
The study was carried out as part of a state-funded assignment (topics 0084-2014-0001 and 0084-2014-0005) and with financial support from the Russian Foundation for Basic Research grant no. 18-04-01313.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wasserman, L.A., Yurina, L.V., Vasilieva, A.D. et al. Effect of Hypochlorite- and Hydrogen Peroxide-Induced Oxidation of Fibrinogen on its Thermal Denaturation. Russ. J. Phys. Chem. B 15, 1036–1040 (2021). https://doi.org/10.1134/S1990793121060105
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1990793121060105