
Abstract
Aim to study the molecular mechanisms of apoptotic death of mouse testicular teratocarcinoma cells (line F-9) under exposure to the widely used selenium-containing compounds with antitumor activity, sodium selenite and methylseleninic acid. Methods fluorescence microscopy, MTT assay, Western blotting. Results It was shown that sodium selenite at a concentration of 10 μM and methylseleninic acid at concentrations of 1 and 10 μM cause apoptosis-dependent death of F-9 cells, excluding necrotic death. Western blotting showed an increase in the expression of XBP1s when treating F-9 cells with 1 μM methylseleninic acid. Conclusions 10 μM methylseleninic acid leads to cell apoptosis, most likely by activation of the IRE1 signaling pathway under prolonged stress of the endoplasmic reticulum.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Sodium selenite (SS) and methylseleninic acid (MSA) are widely used potential anticancer selenium-containing compounds, which was repeatedly confirmed in a number of studies [1–11]. It is known that the cell death under the influence of SS and MSA is associated with ER-stress [1, 2, 11, 12], which is related to the signaling pathways united by a common trigger mechanism and represented by the triad of transmembrane proteins: PERK, IRE1, and ATF6. The purpose of this study was to investigate the changes in the expression of the key members ATF4 and XBP1s of two signaling pathways, PERK and IRE1, respectively, by Western blotting analysis after the treatment of F-9 cancer cells (mouse testicular teratocarcinoma) with SS and MSA at concentrations of 1 and 10 μM.
We have previously found that the treatment of F-9 cells with 1 and 10 μM SS decreased their viability [12]. However, no similar experiments with MSA have been performed. To establish the pathway of death (apoptosis/necrosis) of the studied cells after their 24-h exposure to SS and MSA at concentrations of 1 and 10 μM, we used the Apoptosis/Necrosis Detection Kit (Abcam, United States) in tandem with fluorescence microscopy. This fluorescence analysis makes it possible to detect live cells (blue, stained with CytoCalcein Violet 450), apoptotic cells (green, stained with Apopxin Green), and necrotic cells (red, stained with 7-AAD). Before treatment with the selenium-containing compounds, F-9 cells were seeded in a 96-well plate in DMEM medium containing 10% serum and grown in a CO2 incubator to a density of 5 × 105 cells/well for 24 h, after which SS and MSA were added to the cells at appropriate concentrations. After incubation for 24 h, the cells were resuspended in the assay buffer according to the manufacturer’s protocol. The apoptotic cells were detected using an Apopxin Green Indicator and visualized using the FITC channel (Ex/Em = 490/525 nm). The healthy cells were detected using CytoCalcein 450 (Ex/Em = 405/450 nm), and the necrotic cells were detected using 7-aminoactinomycin D (Ex/Em = 550/650 nm).
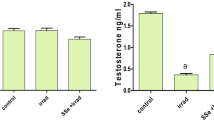
Figure 1 shows the results of qualitative analysis, which provide information about the presence/absence of apoptosis or necrosis in the cancer cells studied. They show that, after a 24-h treatment with 10 μM SS and 10 μM MSA, apoptosis was observed in all visualized cells (100%). After the treatment of cells with 1 μM SS, the majority of cells did not lose their adhesive properties and retained viability, although their morphology changed (cells acquired spherical shape). Interestingly, when cells were treated with 1 μM MSA, approximately 50% of the cells underwent apoptosis, whereas the remaining 50% remained viable.

Fluorescent analysis of the viability of F-9 cancer cells after their 24-h exposure to SS and MSA at concentrations of 1 and 10 μM. The live cells were stained with CytoCalcein Violet 450 (blue), and the apoptotic cells were stained with Apopxin Green Indicator (green).
To establish the signaling pathways that are activated as a result of the treatment of cells with SS and MSA in each case, we tested how the expression of their key components changed by Western blotting analysis. For this purpose, cells were exposed to 1 and 10 μM SS and MSA for 24 h, after which they were lysed with the buffer PBS 1 (pH 7.4) containing 1 mM PMSF. The total protein content in each sample was measured colorimetrically by the method of Bradford. Proteins were separated by SDS-PAGE in 12% polyacrylamide gel. The content of proteins in each sample (cell lysate) was 50 μg. We used the commercial primary antibodies against the transcription factor ATF4, XBP1 (X-box binding protein 1), and the reference protein GAPDH at a dilution of 1 : 200 and the secondary antibodies conjugated to horseradish peroxidase (Abcam, United States) at a dilution of 1 : 2000 (Fig. 2).

Western blot analysis of protein expression (XBP1s, ATF4 and GAPDH) in F-9 cancer cell lysates after 24-h exposure to SS and MSA at concentrations of 1 and 10 μM by immunoblotting: (1) protein content in intact cells and protein content after exposure of cells to (2) 1 μM SS, (3) 10 μM SS, (4) 1 μM MSA, and (5) 10 μM MSA.
Proteins were quantitated using the Fusion Solo chemiluminescence documentation system (Viber, France). This experiment was repeated three times (Fig. 3). It was found that, when cells were treated with 1 μM MSA, the expression of the spliced form XBP1s significantly increased, which may indicate the activation of the IRE1 signaling pathway. The XBP1 transcription factor is involved in the expression regulation of the genes that are required for the immune system functioning and in the regulation of response to ER-stress. Activated IRE1 (inositol-requiring enzyme 1) catalyses the elimination of the noncanonical intron (26 nucleotides) from the ubiquitously expressed XBP1u mRNA (unspliced XBP1, 29.5 kDa), which causes a frame shift in the coding sequence XBP1 and translational of the XBP1s isoforms (spliced XBP1, 55 kDa).

Quantification of XBP1s and ATF4 proteins with respect to GAPDH in F-9 tumor cell lysates after their 24-h exposure to SS and MSA: (1) protein content in intact cells and protein content after exposure of cells to (2) 1 μM SS, (3) 10 μM SS, (4) 1 μM MSA, and (5) 10 μM MSA.
In all other samples of cells treated with both SS and MSA, the expression level of XBP1u and XBP1s was approximately the same. In the case of activation of the PERK signaling pathway, detection of an increase in the expression of the ATF4 transcription factor induced by phosphorylation of eIF2α, the key component of protein translation, could be expected. However, after the treatment of cells with both SS and MSA at the studied concentrations, the expression level of ATF4 did not differ from the control (intact F-9 cancer cells). Possibly, the apoptosis of cancer cells, in which no increase in the expression of either ATF4 or XBP1s was observed, was caused by the activation of the third signaling pathway ATF6. This requires a more detailed analysis and confirmation in further studies.
REFERENCES
Shigemi, Z., Manabe, K., Hara, N., et al., Methylseleninic acid and sodium selenite induce severe ER stress and subsequent apoptosis through UPR activation in PEL cells, Chem. Biol. Interact., 2017, vol. 266, pp. 28–37. https://doi.org/10.1016/j.cbi.2017.01.027
Han, B., Ren, Y., Guan, L., et al., Sodium selenite induces apoptosis in acute promyelocytic leukemia-derived NB4 cells through mitochondria-dependent pathway, Oncol. Res., 2009, vol. 17, no. 8, pp. 373–381.
Ramoutar, R.R. and Brumaghim, J.L., Effects of inorganic selenium compounds on oxidative DNA damage, J. Inorg. Biochem., 2007, vol. 101, no. 7, pp. 1028–1035. https://doi.org/10.1016/j.jinorgbio.2007.03.016
Battin, E.E., Zimmerman, M.T., Ramoutar, R.R., et al., Preventing metal-mediated oxidative DNA damage with selenium compounds, Metallomics, 2011, vol. 3, no. 5, pp. 503–512. https://doi.org/10.1039/c0mt00063a
Okuno, T., Honda, E., Arakawa, T., et al., Glutathione-dependent cell cycle G1 arrest and apoptosis induction in human lung cancer A549 cells caused by methylseleninic acid: comparison with sodium selenite, Biol. Pharm. Bull., 2014, vol. 37, no. 11, pp. 1831–1837. https://doi.org/10.1248/bpb.b14-00453
Husbeck, B., Bhattacharyya, R.S., Feldman, D., et al., Inhibition of androgen receptor signaling by selenite and methylseleninic acid in prostate cancer cells: two distinct mechanisms of action, Mol. Cancer Ther., 2006, vol. 5, no. 8, pp. 2078–2085. https://doi.org/10.1158/1535-7163
Zeng, H. and Wu, M., The inhibitory efficacy of methylseleninic acid against colon cancer xenografts in C57BL/6 mice, Nutr. Cancer, 2015, vol. 67, no. 5, pp. 831–838. https://doi.org/10.1080/01635581.2015.1042547
Tarrado-Castellarnau, M., Cortés, R., Zanuy, M., et al., Methylseleninic acid promotes antitumour effects via nuclear FOXO3a translocation through Akt inhibition, Pharmacol. Res., 2015, vol. 102, pp. 218–234. https://doi.org/10.1016/j.phrs.2015.09.009
Varlamova, E.G., Goltyaev, M.V., and Kuznetsova, Yu.P., Effect of sodium selenite on gene expression of SELF, SELW, and TGR selenoproteins in adenocarcinoma cells of the human prostate, Mol. Biol. (Moscow), 2018, vol. 52, no. 3, pp. 446–452. https://doi.org/10.1134/S0026893318030147
Varlamova, E.G., Goltyaev, M.V., Novoselov, V.I., et al., Cloning, intracellular localization, and expression of the mammalian selenocysteine-containing protein SELENOI (SelI) in tumor cell lines, Dokl. Biochem. Biophys., 2017, vol. 476, pp. 320–322. https://doi.org/10.1134/S160767291705012X
Varlamova, E.G. and Goltyaev, M.V., The effect of sodium selenite on the expression of genes of endoplasmic reticulum-resident selenoproteins in human fibrosarcoma cells, Biophysics (Moscow), 2018, vol. 63, no. 5, pp. 700–705. https://doi.org/10.1134/S000635091805024X
Kuznetsova, Yu.P., Goltyaev, M.V., Gorbacheva, O.S., et al., Influence of sodium selenite on the mRNA expression of the mammalian selenocysteine-containing protein genes in testicle and prostate cancer cells, Dokl. Biochem. Biophys., 2018, vol. 480, pp. 131–134. https://doi.org/10.1134/S1607672918030018
Funding
This work was supported by the Russian Foundation for Basic Research (project no. 18-34-00118 mol_a) and the program of the Presidium of the Russian Academy of Sciences “Postgenomic Technologies and Innovative Solutions in Biomedicine.”
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
Translated by M. Batrukova
Rights and permissions
About this article

Cite this article
Goltyaev, M.V., Varlamova, E.G., Novoselov, S.V. et al. Activation of Signal Pathways of Apoptosis under Conditions of Prolonged ER-Stress Caused by Exposure of Mouse Testicular Teratoma Cells to Selenium-Containing Compounds. Dokl Biochem Biophys 490, 9–11 (2020). https://doi.org/10.1134/S160767292001007X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S160767292001007X