
Abstract
We surveyed the vegetation in Deogyusan National Park (conservation areas) and surrounding areas outside the park (Mt. Juksang, Mt. Johang and Mt. Bonghwa). We calculated plant community indices and diversity indices; these identified properties of the surrounding areas that are relevant to conservation efforts in the park. We compared the diversities of plant communities and plant species between areas using field and quadrat surveys to assign the identities of plant communities to an existing un-delineated preliminary vegetation map produced from aerial photographs. We identified 19 associations. The covers of dominant species in all tree and herb layers were markedly higher in the conservation areas than in the peripheral zone. Mean richness (based on plant community richness and species richness indices) was significantly higher in the conservation area, but mean values of community dominance, diversity, and evenness and the Simpson, Shannon diversity, and evenness indices in the surrounding area were significantly higher than those in the conservation area. Ordination by detrended canonical correspondence analyses showed that elevation and slope explained the distributions of quadrats in the Cartesian space of the ordination. The cover of dominant species in the tree, herb, and sub-tree layers and species numbers in quadrats explained the variation in species compositions within the quadrats. Our results demonstrate that disturbances in the areas surrounding a park can have increasing impacts on plant community and diversity indices. Hence, the preservation and management of surrounding areas are essential conservation elements for protecting whole national park areas.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Individual species and species assemblages are indices of forest integrity [1]. They function as indicators of ecosystem conditions. National parks have had important conservation functions since the creation of a worldwide national park system. Human activities have often had destructive impacts on national parks.
Twenty-two areas have been designated national park lands in South Korea, and these occupy 6.7% of the republic’s terrestrial landscape. The Korea national park service, which was established in 1987, is charged with managing 21 of the national parks; the Service does not manage Hallasan national park (http://english.knps.or.kr/). Deogyusan national park became Korea’s tenth national park in Korea in 1975. It is threatened by human activities, such as ski resort development, plantation development, and medicinal plant collection, although they are all prohibited by the Natural Parks Law of Korea.
The area surrounding a national park has an important role in protecting the entire park, including the core area. The transition areas can function as external support zones where limited development is permitted while protecting the core areas, as in the biosphere reserve strategies of the United Nations [2]. The transition areas are outside of the national park protected areas but are subject to environmental scrutiny to mitigate the impacts of surrounding human activities on the national parks themselves [3]. Hence, the ecological conditions in the gradient from the conserved core into the surrounding transition area should be studied to establish the current status and the relationships between the two zones. An understanding of ecological zonation in human-impacted and natural systems can facilitate appropriate park-protection strategies [4]. The term vegetation includes all plant life in a particular area, and is synonymous with “plant community” [5]. Elevation and climatic variables shape the forest community structure of the Singalila national park in the eastern Himalaya of India [6]. Land use influences vegetation structure; a mixture of protected areas and zones open to human use can help promote biodiversity conservation in savanna vegetation [7].
In this study, we compared the vegetation of a national park (conservation area, CA) and the surrounding areas (SA) to inform management planning for the two systems.
MATERIAL AND METHODS
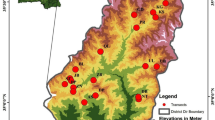
Study site. We compared a sector of the CA within the Deogyusan national park with the surrounding Muju-Jinan mountain zone (Mt. Juksang, Mt. Johang, and Mt. Bonghwa) located at the northwestern boundary of the national park (35°44–36°02′ N, 127°30–127°46′ E) (Fig. 1). Deogyusan, which is at the center of the montane backbone of Korea, was designated a national park in 1975 in Notice no. 25 of the Ministry of Land, Transport, and Maritime Affairs. The park lies at the heart of Baekdudaegan (backbone mountain ridges of Korea), and has an area of 231.650 km2. It houses 1067 plant species and a total biota of 2039 species (http://english.knps.or.kr/ Knp/Deogyusan). The plant biome of the park is in the southern part of the cool temperate zone [8]. The mountain forest vegetation of the Choksangsan sector of the park has been subdivided into the following categories: deciduous broad-leaved forest, valley forest, coniferous forest, plantation, and other vegetation [9]. The summit heights in the zone surrounding the park are 1038 m (Mt. Juksang), 799.3 m (Mt. Johang), and 884.5 m (Mt. Bonghwa).

Study area. The right map shows the southern sector of the Republic of Korea. The left map enlargement shows Deogyusan National Park (green) and the surrounding area (red).
Deogyusan national park is managed by the Korea national park service to protect the biota and ensure sustainable use. The National park service has three special protection zones in the Chilyeon Ravines and at the foot of Hyangjeokbong Peak inside the park where important wild plant habitats are protected. Most of the human population living in the SA is engaged in agriculture. Vegetation there is strongly influenced by human agricultural activities (including sowing and harvesting), which cause unnatural disturbance and patterns of succession in the vegetation.
The climate parameters at Jangsugun, which is close to the study site, were as follows in 1981–2010: average temperature – 10.5°C; average annual highest temperature – 17.0°C; average annual lowest temperature – 4.9°C; average annual precipitation – 1464.3 mm (http://data.kma.go.kr).
Deogyusan national park is located on typical inland montane terrain where the profile is hilly and the soils are rich in humus and poor in minerals. The most common soils are acidic brown forest soil, lithosols, and red-yellow soil. Most of profile at the study site is steeply sloping. Areas with an easy access slope of ≤10% occupy only 1.5 km2, accounting for only 0.7% of the total area; slopes with a gradient of >30% occupy 77.6% of the total area, i.e., this is rugged mountain terrain [10].
Vegetation sampling. We studied both the CA and SA. The SA included the mountain slopes of Juksang (non-conservation area), Johang, and Bonghwa. The CAs were divided into slopes on Mt. Juksang and Mt. Deogyu. We assigned the plant communities identified by physiognomic indices obtained in direct field surveys to polygons on maps delineating different vegetation types recognized by aerial photographs and digital maps (scale 1 : 25000) provided by the National Institute of Ecology, Republic of Korea (http://www. nie.re.kr/contents/siteMain.do?mu_lang=ENG). We conducted quadrat sampling to determine the plant species composition, plant species cover and dominance (covers of dominant plant species in the tree (T1), sub-tree (T2), shrub (S), and herb (H) layers) and environmental conditions (elevation, aspect, and degree of slope) in the CA and SA. Quadrats measured 10 × 10 m and were sufficiently large to include tree species in the canopy layers of the forests. The quadrat locations were determined randomly within each of the plant community polygons designated in a previous field survey. The polygons included in the quadrat surveys were selected randomly. Plant species cover was quantified using the Braun-Blanquet scale [11]. The class numbers of the scale were transformed into mean values following the procedures of Mueller-Dombois and Ellenberg [12].
Overall, 25, 6, and 17 quadrats were deployed in the Juksang, Johang, and Bonghwa SAs, respectively (total: 48). We deployed 53 quadrats in the Mt. Juksang CA. The nomenclature and classification used for the vascular plants followed Lee [13] and park [14, 15].
Plant community indices. We calculated four plant community indices derived from the conventional diversity index to compare community diversities among the CA and SAs: the richness, dominance, diversity, and evenness indices. For each, we replaced species number (s) with the plant community number (c) and the individual numbers of each species (n) with the individual number per plant community (n). Each segmented polygon on the study maps was defined as a plant community based on our field survey; the polygon compartment was also examined by the National Institute of Ecology, Republic of Korea, using aerial photographs (scale 1 : 25000). Each of the communities was identified and confirmed through actual vegetation sampling.
The plant community richness index was calculated as the number of plant community types in the study area. The plant community richness index indicates the number of plant community types classified physiognomically in study sites which means the diversity of plant communities to be used for checking plant community pattern [16].
The plant community dominance index was calculated as 1–D` where:
D' is the plant community diversity index modified from Simpson’s diversity index [5], c is the plant community number, and n is the individual number of each species per plant community.
Species dominance can be an indicator of community condition [17]. The plant community was identified generally by dominant species physiognomically. Therefore, the plant community dominance index produces the extent both to which species dominates in the plant communities in study areas and then which plant community dominates in study areas.
The plant community diversity index (H ') was calculated as:
H ' is the plant community diversity index and the plant community diversity index was transformed from the Shannon-Wiener index [5] on the different condition that species information is displaced by plant community information. In this case, the plant community diversity index demonstrates how diverse plant communities are in study area.
The plant community evenness index (E) was calculated as:
E = eH '/c,
The plant community evenness index was transformed from the Pielou’s evenness index [18] on the different condition that species information is displaced by plant community information. In this case, the plant community evenness index demonstrates how even plant communities are in study area.
We obtained only one value for each index for the whole study site; accordingly, statistical significance was verified using the bootstrap procedure [19]. The statistic estimated from the original sample was t. The simulated estimates from R bootstrap replicates were \(t_{1}^{*},t_{2}^{*}, \ldots ,t_{R}^{*}\). To calculate 95% confidence intervals, we set a one-tailed error α of 0.025. Among three different bootstrap methods, we selected the basic procedure and calculated the basic bootstrapped confidence interval (A) as:
t is the statistic produced from the original sample, R is an optional integer value such as 999, 9999 or 99999 and α is 0.025 at a one-tailed error under 95% confidence intervals. All plant community indices and bootstraps were calculated using PAST 3.22 version software [19, 20]. Species richness, the Simpson index, the Shannon index, and the evenness index were calculated from plant species identities and their individual numbers within each quadrat in the CA and SAs. We pooled the data for the three SAs before calculating the plant community and diversity indices [21, 22].
Ordination. To analyze the differences in vegetation structure between CA and SAs and to identify significant correlations with environmental variables, we ordinated the samples using detrended canonical correspondence analysis (DCCA). The relative covers of herb (H) and woody species (T1, T2, S) layers were ordinated in relation to seven environmental variables (e.g., elevation, aspect, slope). Our premise was that plots in the CA would cluster together and separately from clustered plots from the SAs assuming homogeneity of vegetation types within areas. All of ordination analyses were performed using CANOCO 4.55 software [23].
Statistical analyses. The plant community indices were calculated with bootstrapping 95% confidence intervals using a specified number of bootstrap replicates. To verify and compare the significance of the mean values of the indices, we used the Kruskal-Wallis test to examine data from Mt. Juksang, Mt. Bonghwa, Mt. Johang, and Mt. Juksang for overall comparisons over the entire study site (p < 0.05). We found a significant difference in plant community indices between CA and SAs (Kruskal-Wallis test; p < 0.05); therefore, we compared the means of the indices for each study area using the Mann-Whitney test (p < 0.05). The cover of dominant species in each vegetation layer and the plant community indices of the tree and herb layers were examined statistically using the Mann-Whitney test (p < 0.05). All statistical analyses were performed using SPSS for Windows 22.0 software (IBM, 2013).
RESULTS
Characteristics of vegetation. We found 190 plant species in the CA (53 quadrats) and 169 plant species in the SAs (48 quadrats). We detected a total of 211 plant communities in the CA (192) and SAs (39); 20 plant communities were common to both areas. Among plant associations, the Pinus densiflora – Quercus variabilis community had the highest rank among vegetation conservation indices (I: climax community; II: community restored by secondary succession; III: disturbed community; IV: afforestation; V: orchard or arable lands; https://egis.me.go.kr/ main.do). The Pinus densiflora – Quercus variabilis community was recorded along the periphery of Mt. JukSang (Deogyusan National Park). The degraded areas within forests of the peripheral zone were produced by clear cutting, plantation development after clear cutting, and agricultural cultivation. Private forest property owners continued to clear cut; plant horticultural tree species such as Pinus densiflora, Pinus rigida, Liriodendron tulipifera, and Thuja orientalis; and grow forest products with permission of the Korea Forest Service. Two exotic species (Paspalum dilatatum and Robinia pseudoacacia) in the SA occurred in 23 of the 48 quadrats. Pinus densiflora, Quercus mongolica, Quercus serrata, Quercus variabilis, Fraxinus mandshurica, and Cornus controversa were dominant in the canopy layers in the vegetation of the CA.
The covers of dominant species differed significantly in all vegetation layers (tree, sub-tree, and herb layers) between the CA and SAs (Table 1, p < 0.05).
The covers of dominant species in all tree and herb layers were markedly higher in the CAs, but cover in the shrub layers did not vary between the CA and SAs. The proportions of vegetation types also differed significantly between the CA and SAs. The proportion of plantation area was nearly 3- to 4-fold higher in the SAs than in the CA. The proportions of other vegetation types were also clearly different between the areas. These trends were apparent across the entire CA.
Plant community indices. Most of the plant community indices were higher in the SAs than in the CA whereas the plant community richness was higher in the CA than in the SAs (Fig. 2). The quadrat data were subjected to Mann-Whitney test analyses (p < 0.05). We found statistically significant differences between the CA and SAs in all diversity indices (Fig. 3). The mean species richness (alpha diversity) was significantly higher in the CA (Fig. 3). However, the mean Simpson, Shannon diversity, and evenness indices were significantly higher in the SA (Fig. 3). The diversity indices for tree and herb layers were all significantly different between the CA and SAs (p < 0.05).

Mean confidence intervals of plant community indices for different sites based on plant community numbers (95% bootstrap confidence interval, n = 9999; Surrounding areas: (A) Mt. Juksang, (B) Mt. Johang, (C) Mt. Bonghwa, (D) Entire surrounding area; Conservation area: (E) Mt. Juksang, (F) Deogyusan National Park).

Mean confidence intervals of diversity indices for different sites based on plant species and numbers (Surrounding area: (A) Mt. Juksang, (B) Mt. Johang, (C) Mt. Bonghwa, (D) Entire surrounding area; Conservation area: (E) Mt. Juksang). Different italic letters like as a and b identify statistically different values (Mann-Whitney test, p < 0.05) and ab represents statistically similar values to a and b.
Relationship between vegetation and environmental factors. We performed DCCA ordination analyses that included the vegetation and environmental variables in the two areas based on relative species cover (Fig. 4; first axis length: 4.22; cumulative percentage variance in the species–environment relationship: 31.8%; second axis length: 3.19; variance explained by the relationships: 44.0%). The ordination was based on 254 species in 101 quadrats (53 quadrats in the CA and 48 quadrats in the SAs) and eight environmental factors (aspect, slope, elevation, covers of dominant species in T1, T2, S, and H layers, number of species). A Monte-Carlo permutation test indicated that the first canonical axes were significantly different (F = 4.555, p = 0.002, n = 499).

Detrended canonical correspondence analysis (DCCA) ordination of study sites. Correlations with environmental factors are shown for cases where r > 0.241 (Axis 1: slope, r = 0.4318; elevation, r = –0.9581; cover of dominant species in the T1 layer, r = –0.7075; cover of dominant species in the T2 layer, r = –0.5433; cover of dominant species in the H layer, r = –0.7023; number of species, r = –0.5246; Axis 2: elevation, r = 0.2600; cover of dominant species in the T1 layer, r = –0.5199; cover of dominant species in the S layer, r = –0.4645). (a): ⚪—conservation area, ◻—surrounding area. (b) Plant community types: ⚪—Quercus mongolica, ◻—Pinus densiflora,  —Quercus variabilis,
—Quercus variabilis,  —Quercus serrata,
—Quercus serrata,  —Carpinus laxiflora,
—Carpinus laxiflora,  —Fraxinus mandshurica,
—Fraxinus mandshurica,  —others.
—others.
The means of six environmental factors in the CA and SA were significantly different (p < 0.05), but the covers of dominant species in the shrub layer and aspect were not. The significance level (r > 0.241) was set with critical values from the correlation coefficients table [24]. The first axis of the ordination was highly correlated with the angle of slope (CA = 21.04°, SA = 29.02°), elevation (CA = 740.85 m, SA = 385.40 m), the cover of dominant species in the T1 layer (CA = 91.68%, SA = 66.65%), the cover of dominant species in the T2 layer (CA = 39.43%, SA = 30.74%), the cover of dominant species in the H layer (CA = 39.25%, SA = 20.65%), and the number of species (CA = 27.00, SA = 17.06). The second axis was highly correlated with elevation (CA = 740.85 m, SA = 385.40 m), the cover of dominant species in the T1 layer (CA = 91.68%, SA = 66.65%), and the cover of dominant species in the S layer (CA = 32.92%, SA = 31.15%). There was little correlation between aspect (CA = 150.21°N, SA = 184.49°N) and the DCCA axes (Fig. 4a).
In the ordination plot, the quadrats from the CA and SAs clustered separately (Fig. 4a). Within the ordination coordinate space, quadrats were classified and clustered by six types of plant communities (Quercus monglica, Pinus densiflora, Quercus variabilis, Quercus serrata, Carpinus laxiflora, and Fraxinus mandshurica) (Fig. 4b).
DISCUSSION
Vegetation of the conservation and surrounding areas. The cover of dominant tree species in the CA was significantly higher than that in the SAs but this was not the case for the shrub layers. The cover of dominant shrubs was similar under the canopies of the sub-tree and tree layers. Shrub development was limited in both the CA and SAs. The cover of dominant herbaceous species was significantly higher in the CA largely because two invasive exotic species, Paspalum dilatatum and Robinia pseudoacacia, thrived in the SAs, where they had competitive pressure on native species, thereby reducing native species’ cover values. Invasive species are common in harvested and disturbed sites in the managed portions of the boreal forest of eastern North America [25]. Intensive human activities, such as agriculture, tree thinning, and medicinal plant overharvesting, can have negative impacts on the growth of plants in the herb layers. The small and large-scale disturbances can be critical determinants of forest dynamics [26].
The greater area under plantation forests in the SAs may have disturbed the park ecosystem. Land-use patterns and intense anthropization may change the surrounding vegetation beyond national park boundaries [7, 27]. The vegetation information such as vegetation maps revealed intensive anthropogenic disturbances in forested, savanna, and semi-arid areas subjected to intensive agricultural use outside of the Park boundaries like this study [28].
Plant community indices in the conservation and surrounding areas. Plant community indices can be used to broadly analyze regional diversity, and they extend the physical range of conventional diversity indices. We compared the overall regional trends using both the new index and the conventional diversity indices. The trends were similar for the two index types (Figs. 2, 3). The values of the conventional species diversity indices for the study sites were similar or a little higher than those for other sites in Korea and in other countries (Shannon diversity index national range: 1.2202–1.3428 [29, 30]; Shannon-Wiener index of the upper reaches: 0.1–1.2 [31]). The Shannon diversity indices that we calculated for the SAs and CAs were similar to those of the temperate vegetation zones in other nations [32]. And the Shannon diversity indices CAs covered the range of those of the urban ecosystems in East China [33].
The numbers of plant communities and plant species were higher in the CA than in the SAs, whereas the plant community dominance index and the Simpson dominance index were higher in SAs. This is indicative of dominance by some of the plant community species. Our calculations showed that the SAs had greater diversities of plant communities and plant species than the CA, likely because the long-lived dominant species in the CA suppressed diversity in the underlying vegetation. The distribution of the numbers of plant community and plant species were more even in the SAs than in the CA.
The Shannon-Wiener index has been widely used to analyze the different effects of stress and disturbance on the diversity of plant communities. We found that the plant community indices can also be effective indicators of conditions in plant community; values for these indices were similar to those of conventional diversity indices (Simpson dominance, Shannon-Wiener index and evenness index). The plant community indices could be used to compare similarities and dissimilarities between ecosystems.
Ordination analyses. Within the coordinates of the ordination plot, the quadrats from the CA and SAs clustered separately. Among environmental variables, elevation and slope had the greatest correlation with vegetation parameters. These nonbiological variables contributed to explaining the distribution of quadrats in the ordination plot. Elevation and slope influence temperature and soil conditions, such as water and nutrient contents and have marked impact on vegetation growth [34]. The covers of dominant species in the tree, herb, and sub-tree layers, and species numbers also contributed substantially to explaining the distribution of quadrats in the ordination. The vertical structure of tree species in an old-growth temperate forest can play an important role in population regeneration and community dynamics [35]. Hence, the dominance of tree species in the vertical community structure of the forests in the CA and SAs may have influenced species distributions in ordination space. The dominance of some plant species can influence the establishment and distribution of other plant species. The variation in the distributions of quadrats in the ordination space was reliably related to the distributions of plant community types. Thus, the distribution of quadrats in ordination space can help explain differences in plant community types.
Effects of management protocols in the surrounding area on the vegetation in the conservation zone. The properties of SAs are related to the vegetation structure and function in CAs. Organisms, materials, and nutrients can be exchanged across park boundaries in spite of fragmentation by roads and other artificial barriers. According to meta-community concepts, some of the source communities in the SAs and CA may contribute biological and physical elements to sink communities. We found that plant communities in the national park had stable configurations with low values for plant community indices and diversity indices, which is indicative of late stages in forest succession. The high plant species diversity in the national park may act as a source contributing taxa to the SAs and to the core of the park itself.
CONCLUSIONS
Most of the plant community and diversity indices were significantly higher in the SAs than in the CA. This is likely related to elevated levels of disturbance and species introductions in the SAs. The covers of dominant species in all of the tree and herb layers were markedly higher in the CA than in the SAs, demonstrating the dominance of a few species in the vertical structure of the CA.
DCCA ordination showed that the distribution of quadrats in Cartesian space was strongly influenced by elevation, cover of dominant species in the tree (T1) layer, cover of dominant species in the herb (H) layers, cover of dominant species in the sub-tree (T2) layers, the number of species and the slope (in decreasing rank order of statistical significance) on the first DCCA axis. This highlights the importance of CAs, such as national parks, for the development of stable plant communities that function as a source of species that can contribute to sink communities.
REFERENCES
Sanders, S. and Grochowski, J., Alternative metrics for evaluating forest integrity and assessing change at four northern-tier U.S. National parks, Am Midl. Nat., 2014, vol. 171, pp. 185–203.
Withgott, J. and Laposata, M., Essential Environment: The Science Behind the Stories, New York: Pearson Education, Inc., 2012.
Xie, Y., Protected Area, Amsterdam, in: Encyclopedia of Ecology, 2nd ed., Faith, B.D., Ed., Amsterdam: Elsevier, 2019, pp. 451–457.
Young, K.R., National park protection in relation to the ecological zonation of a neighboring human community: An example from Northern Peru, Mt. Res. Dev., 1993, vol. 13, pp. 267–280.
Barbour, M.G., Burk, J.H., Pitts, W.D., et al., Terrestrial Plant Ecology, Menlo Park, NJ: Addison Wesley Longman, Inc., 1999.
Sinha, S., Badola, H.K., Chhetri, B., et al., Effect of altitude and climate in shaping the forest composition of Singalila National Park in Khangchendzonga landscape, Eastern Himalaya, India, J. Asia-Pac. Biodivers., 2018, vol. 11, pp. 267–275.
Nacoulma, B.M.I., Schmann, K., Traoré, S., et al., Impacts of land-use on West African savanna vegetation: A comparison between protected and communal area in Burkina Faso, Biodivers. Conserv., 2011, vol. 20, pp. 3341–3362. https://doi.org/10.1007/s10531-011-0114-0
Yim, Y.J. and Kira, T., Distribution of forest vegetation and climate in Korean Peninsula: 2. Distribution of climatic humidity/aridity, Jap. J. Ecol., 1976, vol. 26, pp. 157–164.
Choi, Y.E., Kim, C.H. and Oh, J.G., Community distribution on mountain forest vegetation on the Choksangsan area in the Deogyusan National Park, Korea, Kor. J. Ecol. Environ, 2013, vol. 46, pp. 460–470.
Song, S.K., Study on variation of vegetation in Mt. Deogyu, M.Sci. Dissertation, Changwon: Changwon National Univ., 2009.
Braun-Blanquet, J., Plant Sociology: The Study of Plant Communities, New York: McGraw Hill, 1932.
Mueller-Dombois, D. and Ellenberg, H., Aims and Methods of Vegetation Ecology, New York: Wiley, 1974.
Lee, T.B., Illustrated Flora of Korea, Seoul: Hyangmunsa, 1985.
Park, S.H., Colored Illustrations of Naturalized Plants of Korea, Seoul: Ilchokak, 1995.
Park, S.H., Colored Illustrations of Naturalized Plants of Korea (Appendix), Seoul: Ilchokak, 2001.
Li, D. and Waller, D., Long-term shifts in the patterns and underlying processes of plant associations in Wisconsin forests, Glob. Ecol. Biogeogr., 2016, vol. 25, pp. 516–526.
Frieswyk, C.B., Johnston, C.A. and Zedler, J.B., Identifying and characterizing dominant plants as an indicator of community condition, J Great Lakes Res., 2007, vol. 33, pp. 125–135.
Odum, E.P., Basic Ecology, Philadelphia: Saunders, 1983.
Hammer, Ø., Paleontological Statistics. Oslo: Univ. of Oslo Press, 2018.
Hammer, Ø., Harper, D.A.T. and Ryan, P.D., PAST: Paleontological statistics software package for education and data analysis, Palaeo Electronica, 2001, vol. 4, pp. 1–9.
Whittaker, R.H., Evolution and measurement of species diversity, Taxon, 1972, vol. 8, pp. 11–22.
Pielou, E.C., Ecological Diversity, New York: Wiley, 1975.
Ter Braak, C.J.F. and Smilauer, P., CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5), Ithaca, NY, USA: Microcomputer Power, 2002.
Rohlf, F.J. and Sokal, R.R., Statistical Tables, 3rd ed., New York: Freeman, 1995, pp. 123–125.
Jean, M., Lafleur, B., Fenton, N.J., et al., Influence of fire and harvest severity on understory plant communities, For Ecol. Manag., 2019, vol. 436, pp. 88–104.
Caners, R.T. and Kenkel, N.C., Forest stand structure and dynamics at Riding Mountain National Park, Manitoba, Canada. Commun. Ecol., 2003, vol. 4, pp. 185–204.
Squeo, F.A., Loayza, A.P., López, R.P. and Gutiérrez, J.R., Vegetation of Bosque Fray Jorge National Park and its surrounding matrix in the coastal desert of north-central Chile, J. Arid Environ., 2016, vol. 26, pp. 12–22.
Funch, R.R., Harley, R.M. and Funch, L.S., Mapping and evaluation of the state of conservation of the vegetation in and surrounding the Chapada Diamantina National Park, NE Brazil, Biota Neotrop. 2009, vol. 9, pp. 21–30.
Kim, J.Y. and Lee, K.J., Vegetation structure and the density of thinning for the inducement of the ecological succession in artificial forest, national parks – in case of Chiaksan, Songnisan, Deogyusan, and Naejangsan, Kor. J. Env. Ecol, 2012, vol. 26, pp. 604–619.
Kim, C.H., Oh, J.G., Choi, Y.E., et al., Study on the distribution of plant community in the Deogyusan National Park, Korea, Kor. J. Ecol. Environ., 2013, vol. 46, pp. 570–580.
Zhao, J., Gong, L. and Chen, X. Relationship between ecological stoichiometry and plant community diversity in the upper reaches of Tarim River, northwestern China, J. Arid Land., 2020, vol. 12, pp. 227–238.
Rad, J.E., Manthey, M. and Mataji, A., Comparison of plant species diversity with different plant communities in deciduous forests, Int. J. Environ. Sci. Tech., 2009, vol. 6, pp. 389–394.
Wang, C., Wu, B., Jiang, K., et al., Canada goldenrod invasion affect taxonomic and functional diversity of plant communities in heterogeneous landscapes in urban ecosystems in East China, Urban For. Urban Green., 2019, vol. 38, pp. 145–156.
Wang, B., Xu, G., Li, P., et al., Vegetation dynamics and their relationships with climatic factors in the Qinling Mountains of China, Ecol. Indic., 2020, vol. 108, pp. 1–11.
Hao, Z., Zhang, J., Song, B., et al., Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest, For. Ecol. Manag., 2007, vol. 252, pp. 1–11.
ACKNOWLEDGMENTS
This study was financially supported by the National Institute of Ecology, Korea. The authors declare that they have no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jeong, S.C., Lee, I.W., Lee, J.Y. et al. Differences in the Vegetation within Deogyusan National Park and the Surrounding Landscape: Implications for National Park Conservation. Russ J Ecol 52, 455–462 (2021). https://doi.org/10.1134/S1067413621060059
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1067413621060059