
Abstract
The chemical composition of sponge spicules was studied based on the debatable fossil early Cambrian sponge Lenica unica Gorjansky, 1977 from the Siberian Platform. The study has shown that the skeleton of this giant sponge is built of large monaxon megascleres that are a natural carbonate–silica–organic compound with a complicated chemical composition. The data we obtained confirm that the earliest early Cambrian sponges formed skeletal elements of a mixed carbonate–biosilica composition. The authors suggest that this skeleton composition was the initial for all spiculate sponges.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The modern biota includes approximately 8000 species of sponges. More than 75% of them are animals with a siliceous skeleton: most common sponges (Demospongiae) and all known glass sponges (Hexactinellida). Their skeleton consists of individual composite biosilica spicules. Representatives of three classes of sponges (hexactinellids, demosponges, and homoscleromorphs), which form a biosilica skeleton, make a significant contribution to the silica cycle on the Earth. Before the appearance of abundant silica-secreting plankton in the Mesozoic Era, sponges were one of the most important accumulators of dissolved silica [26]. In this process, the role of glass sponges is especially noticeable; up to 95% of the dry weight of animals is concentrated in their biosilica spicules [6]. At present, most hexactinellids are deep-sea animals capable of concentrating significant volumes of silica on a small area of the ocean floor [5, 9, 20]. As an example, there are reefs formed by hexactinellids. The extremly large reef system that was formed by glass sponges and stretched for 7000 km in the northern basins of the Tethys Ocean and the ancient North Atlantic developed in the Late Jurassic Epoch [10, 18, 27].
However, ancient representatives of siliceous sponges remain a mystery in many ways. First, because biogenic silica is quickly replaced by other minerals (for example, carbonates, phosphates and pyrite) when it settles into sediments. Secondly, only a few of the earliest (Cambrian) sponges built skeletons from spicules that are similar to spicules of modern sponges.
Hexactinellids and demosponges appeared at the very beginning of the Cambrian Period approximately 538 million years ago (Ma). Among them, the first reef builders were demosponges of the family Antaspidellidae that possessed a massive skeleton of coalescent dendroclone- and desma-type spicules. In reefs, antaspidellids replaced the aspiculate hypercalcified sponges, that is, archaeocyaths [2, 19]. At present, a great number of fossil hexactinellids and demosponges are known and these remains are not only separate spicules, but also whole skeletons containing biosilica compounds [22]. They constituted an essential part of the fossil biodiversity in the Cambrian paleocommunities and were abundant [16]. Moreover, it was in the early Cambrian time that biogenic siliceous deposits began to form, taking the place of abiogenic deposits; biogenic siliceous deposits, represented exclusively by spiculites, that is, marine sedimentary rocks, are completely built by spicules of siliceous sponges and by silica deposited at their dissolution [8]. In the Ordovician–Permian strata, approximately half of the biogenic sedimentary rocks are formed of spiculites.
In this work, we studied the fossil early Cambrian sponges in order to clarify the taxonomic position of the ancestral forms of silicon sponges and the elemental composition of their spicules.
MATERIALS AND METHODS
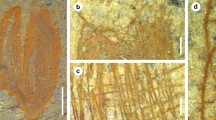
The collection of the material was carried out in the Republic of Sakha (Yakutia) with the assistance of the Administration of the Lena Pillars Natural Park. The studied material is a sample of the lower Cambrian calcareous mudstone with a fragment of a skeleton of the sponge Lenica unica Gorjansky, 1977 (Figs. 1a–1d). The sample belongs to the Sinsk Formation located in the basin of the Buotama River, the right tributary of the Lena River in its middle reaches. The Sinsk Formation was formed in the middle of the Botoman Age of the early Cambrian time (approximately 513–511 Ma) in the relatively deep-water and low-oxygen eastern part of the epicontinental sea that covered the Siberian Platform [30]. The platform was a separate continent, that is, Siberia, which was located at that time in tropical latitudes [25].

The sponge Lenica unica; lower Cambrian, Botoman Stage, Sinsk Formation; the Republic of Sakha (Yakutia). (a) general view of the spiculate/ skeleton, specimen PIN No. 4349/630, the Lena River; (b) fragment of the spicule skeleton, the Buotama River; (c) spicule detail; (d) detail of a spicule showing impregnation of amorphous silica in calcite. Scale: (a), (b), 10 mm; (c), (d), 1 mm.
To study the elemental composition, the samples were preliminarily washed with a solution of hydrogen peroxide. An elemental analysis of the material was carried out using the point method: in a certain part of the sample, several points were selected to form a sector. The elemental composition of the material at these points was determined using energy dispersive X-ray spectroscopy (EDX) on a Zeiss Evo 40 scanning electron microscope (SEM) equipped with an X-ray analyzer and the INC software. A quantitative analysis of the elemental composition of the host rock (in the form of a powder) was carried out at Lomonosov Moscow State University (Faculty of Geology) with the use of a vacuum wave-dispersive X-ray fluorescence spectrometer SPECTROSKAN-MAKS-GV equipped with a high-resolution LiF (200) diffraction crystal.
To determine the organic matter and minerals that form spicules, we used the method of Raman spectroscopy, which makes it possible to recognize the structure of the crystal lattice. In our work, Raman spectra were obtained using a Via Reflex Raman microspectrometer (Renishaw, UK) in combination with a Leica DM2500M universal microscope (Leica-Microsystems, Germany) with an Olympus objective (Japan). Excitation was achieved using a diode laser at a wavelength of 532 nm and a power of 1.0 mW at the object level; the exposure time was 10 s. To obtain the total spectrum, measurements were performed in 10–20 replicates.
RESULTS
The studied sample is a calcareous mudstone (calcareous-argillaceous siltstone) containing large monaxial spicules (megascleras-diactins) of the sponge Lenica unica (Figs. 1a–1d). The mudstone matrix, according to X-ray fluorescence spectrometry data, is almost entirely composed of calcium carbonate (70–71%) with an admixture of clay minerals (SiO2, 20–21%; Na2O, 2.55–2.77%; K2O, 1.06–1.08%; Al2O3, 0.76–1.08%; MgO, 0.69–0.96%; Fe2O3, 0.44–0.53%; TiO2, 0.25%); phosphates (P2O5, 0.11–0.23%); and sulfides (S, 0.84–1.15%).
Spicules are composed of crystals of several minerals. A light microscope displays an alternation of light and dark layers interspersed with a dark-colored mineral (Figs. 1b–1d). SEM shows that the dark mineral is tightly embedded into the light layers (Figs. 1c, 1d). X‑ray spectroscopy made it possible to determine that the light layers are formed by calcium carbonate and the dark layers by iron oxyhydroxide. The inclusions (Figs. 2a–2c) are dominated by carbon (12%), oxygen (60%), and silicon (30%). Calcium and iron are found in trace amounts (approximately 1%). The layers of iron oxyhydroxide (Fe, up to 45%; O, approximately 45%) also contain carbon (up to 9%) and calcium (approximately 4%), but there is practically no silicon (approximately 1%).

A spicule of the Lenica unica sponge, SEM (a) and the elemental composition (% of mass) of amorphous silica (b) and calcite (c) according to the results of EDX analysis. Asterisks mark points of analysis (as, amorphous silica; c, calcite). Scale: 100 μm.
X-ray spectroscopy data are confirmed by Raman spectroscopy (Fig. 3). The Raman spectrum without fluorescence background shows peaks in the range of 129–400 cm–1, corresponding to iron- and sulfur-containing compounds (Fig. 3). The well-defined peak at 466 cm–1 corresponds to the amorphous silica band, which is due to the symmetric stretching mode of the Si–O–Si bond, which is the main vibration of quartz (SiO2). The 1087 cm–1 band corresponds to calcium carbonate in the form of aragonite. Two well-pronounced peaks correspond to the peaks for organic matter assigned to the G band (graphite) at approximately 1598 cm–1 and to the D band (defect band) at approximately 1310–1430 cm–1.

Overlapping Raman spectra showing the main bands of amorphous silica of the Lenica unica spicule and its organic matter content. Modes 129–400 cm–1 correspond to iron-containing and sulfur-containing materials; the peak at 466 cm–1 corresponds to the band of amorphous silica; the peak at 1087 cm–1 corresponds to calcium carbonate, the peak at approximately 1598 cm–1 corresponds to organic matter (graphite).
The Raman spectrum of calcium carbonate, in which the biosilica mineral is impregnated, does not differ from that shown in Fig. 3. Since the shift of the Raman combination scattering curve is due to the strong parasitic luminescence of the sample at a given excitation wave, the analysis needs to remove the background illumination (operation of background radiation subtraction). The spectrum has a well-pronounced peak at 1087 cm–1, corresponding to the peak of aragonite, and a very weak signal from silica, the peak of which is at 466 cm–1.
DISCUSSION
Raman spectroscopy is widely used for analysis of an elemental composition of complex-component compounds. The spectra of iron and sulfur-containing compounds were obtained earlier [28]. Amorphous silica and other silicon compounds [4], and calcium carbonate in the form of aragonite [13] have been studied in many publications. The earlier results were also used for identification of organic matter [15].
The presence of iron oxyhydroxide in the spicules of the sponge Lenica unica is probably explained by the formation of this complex mineral from early diagenetic pyrite, that is, iron sulfide, which, in turn, is usually formed as a result of the activity of sulfate-reducing sulfur bacteria [12]. Conditions favorable to this formation could have developed both in the thin sediment enriched with organic matter, now represented by the mudstone of the Sinsk Formation, and in the anoxic bottom layers of the Siberian Sea Basin, where it formed [21]. The dark color of the rock originates from finely dispersed organic matter [1]. The data of X-ray structural analysis and Raman spectroscopy confirm the high carbon content in iron oxyhydroxide and in amorphous silica of spicules.
The sponge L. unica was discovered in a section of the lower Cambrian Sinsk Formation of the Lena River basin and was referred by the author to the class of six-rayed sponges. Ivantsov et al. found in 2005 additional specimens of this species in the same strata, these specimens contained spicules with an axial canal including a trihedral organic filament [3]. This shape of the filament, which serves as the template for the spicule formation [23], indicates that L. unica belongs to demosponges, regardless of whether the axial canal was enlarged again due to the dissolution of the inner layers of the spicule.
It has been found that sponges of this genus from the lower Cambrian Hetang Formation of Anhui Province (South China) have biminerallic spicules and consist of an inner silica core, an organic outer envelope and a middle probably calcium carbonate layer (the elemental composition was determined using EDX analysis). These data indicate that this sponge does not belong to any of the recent classes but is a transitional form between siliceous sponges (Silicea: Hexactinellida + Demospongiae) and calcareous sponges (Calcarea) and so occupies a basal position in relation to these two main branches in the evolution of the phylum Porifera [7].
Our data on the chemical composition of L. unica spicules partly confirm the earlier conclusions. However, unlike Lenica sp. from the Hetang Formation, the Siberian type species of this genus had not simply biminerallic, but biosilica spicules. Moreover, a significant amount of carbon, oxygen, and silicon in inclusions indicates that the spicules are partially composed not solely of amorphous silica, but of a biosilica composite. The main part of the spicule was formed by calcium carbonate, which has now turned into calcite. However, such a mosaic calcite is a diagenetic mineral and its precursor is mostly often aragonite, whose crystallographic structure differs from that of calcite [29]. The peak at 1087 cm–1 in the Raman spectrum also corresponds to aragonite [13]. Aragonite is typical of recent hypercalcified and some horny Demospongiae, while spicules of calcareous sponges are composed of magnesian calcite [24].
The data in [7] and our results on the study of the composition of L. unica spicules show that very different Cambrian sponges built their skeletons from spicules of mixed carbonate–biosilica composition. Heteractinide sponges, which were previously considered extinct calcareous sponges (Calcarea), and siliceous sponges (Silicea), to which L. unica belongs, are among them. Moreover, the spicules of L. unica have also displayed a significant organic component. It is interesting that a similar skeletal composition is characteristic of recent aspiculate demosponges of the order Verongida, the chitinous fibers of their skeleton are covered with crystalline aragonite and amorphous silica [11].
As studies of the early Cambrian hexactines and pentactines (typical of the Hexactinellida) have shown, these elements had also dense organic envelopes, which are preserved even at complete dissolution of the mineral components of the spicule [14]. Taken together, these data suggest that at the beginning of their evolution, sponges might have had a three-component skeleton, which has been maintained in some extant forms. However, judging by the findings of specific spicules in strata of 538–520 Ma [2], as early as in the first half of the early Cambrian, sponges split into two branches,“calcareous” and “siliceous,” from which all recent classes of sponges developed by the middle of the Cambrian Period.
One of the modern scientific directions in the field of extreme biomimetics pays much attention to the study of composite biosilica materials created on the basis of biominerals [11, 17]. However, modern glass sponges are benthic animals that form large settlements mainly at great depths of seas and oceans; their species diversity accounts only for 7% of the total diversity of sponges [24]. Therefore, the search for the most ancient siliceous sponges and their accumulations is also important for elucidating the possibility of using fossil spiculites and ancient sponge reefs as raw materials with a high silica content.
REFERENCES
Bakhturov, S.F., Evtushenko, V.M., and Pereladov, V.S., Kuonamskaya bituminoznaya karbonatno-slantsevaya formatsiya (The Kuonamka Bituminous Carbonate–Shale Formation), Tr. Inst. Geol. Geofiz., Sib. Otd., Akad. Nauk SSSR, Novosibirsk: Nauka, 1988.
Zhuravlev, A.Yu., Early history of Metazoa: The view of a paleontologist, Zh. Obsh. Biol., 2014, vol. 75, no. 6, pp. 411–465.
Ivantsov, A.Yu., Zhuravlev, A.Yu., Krasilov, V.A., et al., Unique Sinsk locations of Early Cambrian organisms (Siberian Platform), Tr. Palentol. Inst. Ross. Akad. Nauk, 2005, vol. 284, pp. 1–143.
Arasuna, A., Kigawa, M., Fujii, S., et al., Structural characterization of the body frame and spicules of a glass sponge, Minerals, 2018, vol. 8, no. 3, pp. 88. https://doi.org/10.3390/min8030088
Archer, S.K, Dennison, G., Tryon, L., et al., Invertebrate settlement and diversity on a glass sponge reef, Can. Field-Nat., 2020, vol. 134, pp. 1–15.
Barthel, D., Tissue composition of sponges from the Weddell Sea, Antarctica: Not much meat on the bones, Mar. Ecol.: Prog. Ser., 1995, vol. 123, pp. 149−153.
Botting, J.P., Muir, L.A., Xiao, S., Li, X., and Lin, J.P., Evidence for spicule homology in calcareous and siliceous sponges: Biminerallic spicules in Lenica sp. from the Early Cambrian of South China, Lethaia, 2012, vol. 45, no. 4, pp. 463–475.
Chang, S., Zhang, L., Clausen, S., et al., Source of silica and silicification of the lowermost Cambrian Yanjiahe Formation in the Three Gorges area, South China, Palaeogeogr. Palaeoclimatol. Palaeoecol., 2020, vol. 548, pp. 109697. http://doi: https://doi.org/10.1016/j.palaeo.2020.109697.
Chu, J.W.F., Maldonado, M., Yahel, G., and Leys, S.P., Glass sponge reefs as a silicon sink, Mar. Ecol.: Prog. Ser., 2011, vol. 441, pp. 1–14.
Conway, K., Barrie, J., and Krautter, M., Geomorphology of unique reefs on the western Canadian shelf: Sponge reefs mapped by multibeam bathymetry, Geo-Mar. Lett., 2005, vol. 25, no. 4, pp. 205–213.
Ehrlich, H. et al., Insights into chemistry of biological materials: Newly discovered silica–aragonite–chitin biocomposites in demosponges, Chem. Mater., 2010, vol. 22, pp. 1462–1471.
Fürsich, F.T. and Pan, Y., Diagenesis of bivalves from Jurassic and Lower Cretaceous lacustrine deposits of northeastern China, Geol. Mag., 2016, vol. 153, pp. 17–37.
Gunasekaran, S., Anbalagan, G., and Pandi, S., Raman and infrared spectra of carbonates of calcite structure, J. Raman Spectrosc., 2006, vol. 37, no. 9, pp. 892–899.
Harvey, T.H.P., Carbonaceous preservation of Cambrian hexactinellid sponge spicules, Biol. Lett., 2010, vol. 6, no. 6, pp. 834–837.
Hodkiewicz, J., Characterizing carbon materials with Raman spectroscopy, Thermo Fisher Sci. Appl. Note 51901, 2010, pp. 1–5.
Ivantsov, A.Yu., Zhuravlev, A.Yu., Leguta, A.V., et al., Palaeoecology of the Early Cambrian Sinsk biota from the Siberian Platform, Palaeogeogr. Palaeoclimatol. Palaeoecol., 2005, vol. 220, nos. 1–2, pp. 69–88.
Karpenko, A.A. and Drozdov, A.L., Skeletons of glass sponges as a substrate for creating functional composite organosilicon material, J. Mater. Appl., 2019, vol. 8, pp. 20–27.
Krautter, M., Conway, K., Barrie, J.V., et al., Discovery of a “living dinosaur”: Globally unique modern hexactinellid sponge reefs off British Columbia, Canada, Facies, 2001, vol. 44, pp. 265–282.
Kruse, P.D. and Zhuravlev, A.Yu., Middle-Late Cambrian Rankenella-Girvanella reefs of the Mila Formation, northern Iran, Can. J. Earth Sci., 2008, vol. 45, pp. 619–639.
Leys, S.P., Wilson, K., Holeton, H.M., et al., Patterns of glass sponge (Porifera, Hexactinellida) distribution in coastal waters of British Columbia, Canada, Mar. Ecol.: Prog. Ser., 2004, vol. 283, pp. 133–149.
Parfenova, T.M., Korovnikov, I.V., Eder, V.G., et al., Organic geochemistry of the Lower Cambrian Sinyaya Formation (northern slope of the Aldan anteclise), Russ. Geol. Geophys., 2017, vol. 58, pp. 586–599.
Rigby, J.K., Sponges of the Burgess Shale (Middle Cambrian), British Columbia, Palaeontographica Canadiana, 1986, vol. 2, pp. 1–105.
Sandford, F., Physical and chemical analysis of the siliceous skeletons in six sponges of two groups (Demospongiae and Hexactinellida), Microsc. Res. Tech., 2003, vol. 62, pp. 336–355.
Systema Porifera: A Guide to the Classification of Sponges, Hooper, J.N.A. and van Soest, R.W.M., Eds., New York: Kluwer/Plenum, 2002.
Torsvik, T.H. and Cocks, L.R.M., New global palaeogeographical reconstructions for the Early Palaeozoic and their generation, Early Palaeozoic biogeography and palaeobiogeography, Geol. Soc. London Mem., 2013, vol. 38, no. 1, pp. 5–24.
Tréguer, P.J. and De La Rocha, C.L., The World Ocean silica cycle, Annu. Rev. Mar. Sci., 2013, vol. 5, pp. 477–501.
Whitney, F., Conway, K., Thomson, R., et al., Oceanographic habitat of sponge reefs on the Western Canadian Continental Shelf, Cont. Shelf Res., 2005, vol. 25, pp. 211–226.
Yuan, X. and Zheng, H., In situ Raman spectroscopic studies of FeS2 pyrite up to 675 K and 2100 MPa using a hydrothermal diamond anvil cell, Mineral. Mag., 2015, vol. 79, pp. 1–10.
Zhuravlev, A.Yu. and Wood, R., Eve of biomineralization: controls on skeletal mineralogy, Geology, 2008, vol. 36, pp. 923–926.
Zhuravlev, A.Yu. and Wood, R., Dynamic and synchronous changes in metazoan body size during the Cambrian Explosion, Sci. Rep., 2020, vol. 10, pp. 6784. https://doi.org/10.1038/s41598-020-63774-2
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. This article does not describe any research using humans and animals as objects.
Additional information
Translated by I. Barsegova
Rights and permissions
About this article

Cite this article
Drozdov, A.L., Zhuravlev, A.Y., Karpenko, A.A. et al. The Morphology and Chemical Composition of Siliceous Spicules of the Early Cambrian Sponge Lenica unica Gorjansky, 1977. Russ J Mar Biol 48, 33–38 (2022). https://doi.org/10.1134/S1063074022010059
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1063074022010059