
Abstract
A revision of the species composition of the genus Agarum from the Far Eastern seas is based on specimens collected personally by the authors, those deposited at the LE and SAP herbaria, and also obtained during the expeditions of the Zhirmunsky Institute of Marine Biology FEB RAS and SakhalinNIRO to the Kuril Islands. A multigene analysis using the rbcL + cox1 and ITS2 + rbcL+cox1 sequences has shown that the genus Agarum comprises three species. Two of them, A. clathratum Dumortier and A. turneri Postels et Ruprecht, were previously reported from the Asian algal flora. The third species, A. undulatum sp. nov., is here described as new to science. It differs from A. clathratum in ITS2 + rbcL + cox1 by 5.51%; from A. turneri, by 7.5%. The new species, growing in habitats with depths of 15–43 m, is widely distributed in waters along the middle Kuril Islands and the Lesser Kuril Chain. In the phylogenetic tree, A. clathratum splits into two clades, of which one includes specimens belonging to the type form of this species, and the other includes specimens having traits of A. yakishiriense. The difference between the two clades in ITS2 + rbcL + cox1 constitutes only 1.7%. This allows considering A. yakishiriense as a subspecies of A. clathratum. In waters of the Russian Far Eastern seas, it is found in the Sea of Japan and the southern Sea of Okhotsk.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The present report continues the series of our publications on the taxonomy of laminariacean algae from the Far East [11–13, etc.]. Here we consider the Asian members of the genus Agarum Dumortier, 1822. In the Far Eastern region, this genus is distributed almost ubiquitously and plays a major role in the formation of underwater kelp beds [4, 10, 16]. Representatives of the genus Agarum have repeatedly become objects of ecological and taxonomic research based on analyses of phenotypic [2, 4, 10, 15, 17, 20, 21] and genotypic [7–9] characters.
The genus under study is one of the few in the order Laminariales that has a wide Arctic–Pacific–Atlantic range. In the flora of the West Atlantic and all along the Arctic coast, it is represented by a single species, A. clathratum Dumortier, 1822. In the Pacific flora, five species have been known for this genus until recently: A. clathratum, A. fimbriatum Harvey, 1862, A. oharaense Yamada, 1961, A. yakishiriense Yamada, 1962, and A. turneri Postels et Ruprecht, 1840. The latest taxonomic revision of the genus [9], based on data of a multigene analysis, has shown that A. oharaense and A. fimbriatum differ markedly from other members of the genus and form a separate clade, which is more closely related in genetic terms to members of the genus Dictyoneurum Ruprecht, 1852 and Costaria costata (C. Agardh) De A.Saunders, 1895. For this reason, the genus Agarum has been divided into Agarum and Neoagarum H. Kawai et T. Hanyuda, 2017.
For the Far Eastern seas of Russia, two species have been reported to date: A. clathratum and A. turneri. The former was suggested to inhabit the Bering Sea, Sea of Okhotsk, and Sea of Japan. The latter, previously known only from off Kamchatka and the Commander Islands, was found by us much further south [3]. When re-examining the members of the genus Agarum, collected from the middle Kuril Islands, we noted several unusual plants. They differed sharply from other representatives of this genus by a number of specific morphological traits. A multigene analysis using three concatenated sequences (rbcL + cox1 + ITS2) revealed some features of molecular organization of these plants unique enough to describe them as a species new to science, which we named as A. undulatum.
A revision of the materials from the Sea of Japan and the southern Sea of Okhotsk showed that, besides typical representatives of A. clathratum and A. turneri, specimens of Agarum that matched the description of A. yakishiriense were also present in them. Information about findings of such plants in Posyet Bay (southern Primorsky Krai) was first provided in the work on laminariacean algae from the Japanese Archipelago [10]. Then, on the basis of our collections, they were mentioned for Sakhalin and the Sea of Okhotsk as A. clathratum subsp. yakishiriense [7]. However, in subsequent publications of Russian authors with reviews on the algal flora of Peter the Great Bay [1, 6] and commercially valuable and potentially commercial algae of the Russian Far East [5], this taxon was not reported.
The taxonomic position of A. yakishiriense also remained unresolved. Various authors referred to it as an independent species, as a form, or as a subspecies of A. clathratum. In the last revision of this genus [9], it has been mentioned again as a valid species. Due to this uncertainty, we found it relevant to make an extended description of the species and clarify its taxonomic status.
MATERIALS AND METHODS
In this study, we used our materials collected personally from off Sakhalin (in 2018–2019), the southern Kuril Islands (in 2015), and southern Primorsky Krai (in 2020). In addition, we used specimens of Agarum from Ushishir, Simushir, Iturup, Yankich, and Tanfiliev islands, kindly provided to us by members of the Kuril expeditions of the Zhirmunsky Institute of Marine Biology, Far Eastern Branch, Russian Academy of Sciences (in 1971 and 1986) and the Sakhalin Research Institute of Fisheries and Oceanography (in 1967 and 1989). We had an opportunity to examine the type specimens of A. clathratum and A. turneri at the LE herbarium (Komarov Botanical Institute, St. Petersburg), and also the type specimen of A. yakishiriense at the SAP herbarium (Hokkaido University, Sapporo).
To extract DNA, we used pieces of juvenile blades of Agarum specimens, preliminarily rinsed in sterile seawater, frozen in liquid nitrogen, and homogenized for cell destruction. DNA was extracted using an Invisorb Spin Plant Mini Kit (Invitek Molecular, Berlin, Germany). Nuclear ITS was amplified with YB1-LB2 primers (ITS2 region) [18]; the mitochondrial cox1, with GazF2-GazR2 primers [14]. To amplify the rbcL plastid gene, we used RS1-RS2 and KR3-KR4 primers.
A thermally stable enzyme TaKaRa Ex Taq (TaKaRa Bio Inc., Japan) was used for polymerase chain reaction (PCR). Sanger sequencing was performed by a commercial company (Cosmogenetech, Ltd., Korea). The obtained sequences and the sequences for comparative analysis accessed from GenBank were aligned using the Geneious v. 11.0.4 software (Biomatters Ltd., New Zealand) and fitted visually. Phylogenetic trees were generated using the Bayesian analyses (MrBayes 3.2.2) [22] based on the General Time Reversible (GTR) model for nucleotide substitutions, 2 000 000 generations, with every 1000 generations sub-sampled, and a burn-in length of 200 000 generations. For a maximum likelihood analysis, we used RAxML 8.2.11 [19] with the GTR+Gamma substitution model. For a molecular genetic comparison of the Agarum specimens from the Kuril Islands with known representatives of this genus, we carried out a multigene analysis, which showed high efficiency in studies of intrafamilial differentiation in the family Agaraceae [7, 9]. The nucleotide sequences obtained from a representative specimen of the new species are deposited in GenBank under the accession nos. MZ333127 (rbcL), MZ333128 (ITS2), and MZ333129 (cox1).
RESULTS
The multigene analysis of the A. undulatum specimens collected off Ushishir Island, based on the combinations of the rbcL + cox1 and ITS2 + rbcL + cox1 sequences, showed that this species cannot be associated with any of the previously known species of the genus Agarum (Fig. 1). Figure 1a represents the results of a comparative analysis of all known Agarum species and subspecies. In the tree in Fig. 1b, inferred from the ITS2 + rbcL + cox1 combination, A. turneri is absent, since no ITS2 data are currently available for it. The genetic differences between the analyzed representatives of different species are sufficient to classify the unusual specimens from the Kuril Islands as a species new to science. In morphological terms, it also differs from other members of the genus by a number of unique traits.

Phylogenetic trees inferred from concatenated sequences, rbcL+cox1 (a) and ITS2+rbcL+cox1 (b), for different Agarum species. The numerals at the branches are values of statistical bootstrap/a posteriori probability of similarity (with values only above 75 taken into account). The specimens that we examined are indicated by asterisk (*).
Below are the descriptions of the morphological organization, ecology, and distribution of A. undulatum sp. nov. and A. clathratum subsp. yakishiriense from the Russian Far East.
Agarum undulatum N. Klochkova, Klimova, T. Klochkova et Yotsukura sp. nov. (Figs. 2a–2e)

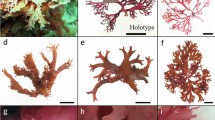
Morphology of Agarum undulatum sp. nov. The type specimen collected off Yankich Island (a), variously aged herbarium specimens from Simushir Island (b) and Ushishir Islands (c), and enlarged fragments of a blade of the type specimen: marginal fringe (d) and ribbed midrib (e). Scale bars: (a, b) 10 cm and (c–e) 1 cm.
Sporophyte consists of blade, stipe, and rhizoids. Blades wide-oval, up to 52 cm in length, with flattened central midrib bearing small longitudinal ribs up to 7 mm wide. Undulations adjacent to each other, present on both sides of blade surface. Blade margins have large outgrowths resembling fringe. Stipe slightly flattened, with longitudinal ribs; holdfast whorled, forming loose bundle. Sporangial sori bilateral, developing as narrow stripe along blade margins.
This species differs from other representatives of the genus by the fringed margin of blades and the abundant undulations on both surfaces of the blade; other differences are in ITS2, rbcL, and cox1 sequences.
Holotype (designated here: Figs. 2a, 2d, 2e) was collected at a depth of 15 m in August 1986 and deposited at the Komarov Botanical Institute RAS (St. Petersburg, LE).
Type locality: Middle Kuril Islands, Ushishir Islands (Yankich Island).
Collector: L.V. Romeiko.
Etymology: The species epithet indicates the undulations on the blade surface as a characteristic feature of its morphology.
Thallus has a blade, a stipe, and a rhizoidal holdfast. Mature blades are 52 cm long, 40 cm wide, 450–700 μm thick, dense, leathery, brown or grayish-brown in color due to the abundant diatom fouling. The shape of blades is wide-oval; the surface is smooth or coarsely folded; the base is rounded (Fig. 2a). Deep folds of the blade, running down to its base, overlap each other and conceal the site of its connection with the stipe. A characteristic feature of the blade structure is the crenate margin all along its circumference. The marginal outgrowths of the blades are spaced from one another, up to 2–4 cm in length. They form a kind of fringe and, in turn, have their own thin, undulating margins. Their structure is shown close-up in Fig. 2d. In some of the plants that we examined, the margins of the blades are frayed and, therefore, they lack regular crenate outgrowths. Blade perforations in this species are extremely rare. These resemble rather irregular holes that appear after puncture. Instead of perforations that are typical for members of the genus Agarum, various-sized bullations develop all over the blade part of thallus (Fig. 2). Some of them are convex on its ventral side, and others are on the dorsal side. Small holes may appear exactly at their apices, but mostly absent. The stipe of this species is rather short, rigid, and terete, with longitudinal ribs. The midrib is slightly flattened, ribbed (Fig. 2e). In the most adult plants, it is up to 7 mm in width. The holdfast rhizoids extend from the stipe base all along its circumference and are clustered into rather loose haptera. Rhizoids are fibrous, multiple-branched. In juvenile specimens, blades are thin, greenish-olive, elongated oval, up to 4 cm in length. The youngest of them lack perforations and bullations. In plants that have reached 10 cm in height, these are already well expressed, have a narrow oval shape, and are longitudinally elongated. Sporangial sori in this species form a narrow, intermittent stripe along the blade margin. It has no distinct boundaries due to the chaotic arrangement of sporiferous patches having hieroglyph-like shapes.
A. undulatum grows on pebbly/sandy sediments at depths of 10–43 m. The ecological optimum for the species is, apparently, a depth range of 15–25 m. Unfavorable habitat conditions lead to inhibition of plant development. In this case, mature plants do not grow higher than 25 cm. Judging by the differences in size and morphology of mature plants, this species is perennial, occurring rarely.
Examined specimens. Middle Kuril Islands. Ushishir Islands: Yankich Island, depth 25 m, three specimens, July 8, 1971; Yankich Island, Kraternaya Cove, depth 15 m, one specimen, collected by L.V. Romeiko; Yankich Island, southern side, depth 43 m, Aug. 1986, six specimens, collected by L.V. Romeiko (used for DNA analysis); depth 15 m, one specimen, Aug. 1986, collected by L.V. Romeiko, (holotype specimen, Figs. 2a, 2d, 2e). Simushir Island: Malaya Cove, depth 19 m, Sept. 6, 1967, two specimens, collected by I.S. Gusarova (Fig. 2b). Southern Kuril Islands: Iturup Island, Zolotaya Cove, depth 10 m, one specimen, July 13, 1989, collected by V.G. Sizov. Lesser Kuril Chain: Tanfiliev Island, Groznaya Cove, depth 15 m, coarse pebbles, sand with shell debris, Aug. 26, 1967, two specimens, collected by I.S. Gusarova.
Agarum clathratum subsp. yakishiriense (Fig. 3).

Morphology of Agarum clathratum subsp. yakishiriense. A specimen from Aniva Bay, Sakhalin Island (a), additional rhizoids forming along the flat stipe in the plants from Peter the Great Bay (b) and Aniva Bay (c), and juvenile specimens from the Gulf of Patience (d, e). Scale bars: (a, d, e) 10 cm and (b, c) 1 cm.
Thallus consists of a blade, a stipe, and a rhizoidal holdfast. Blade is thin, up to 200 μm in juvenile plants, 430 μm in adults, rounded or wide oval, smooth or coarsely folded, 28–40 cm in greater diameter. Its base is cordate in shape. The lower margins of the blade in plants that grew in a eutrophic habitat exposed to low surf impact proliferate and overlap each other. Mature blades have a lacework appearance due to numerous closely located perforations with turned-out scalloped edges. The perforations are rounded, with the largest of them being up to 13 mm in diameter. In the lower half of the blade near the midrib, some of them are elongated oval, transversely stretched. In juvenile plants, these are numerous, very small, forming an even row at the very edge of the blade. The midrib is flattened, from 2.5 to 3.8 mm wide in the largest specimens. On one side of the blade it is slightly convex; on the other side, concave. Thus, the plant has a distinct dorsoventral structure. The margins of the blade are finely folded, especially at the base. The width of the folded band in juvenile plants is up to 1–2 mm; in adults, 1.2 cm. The stipe in this species is short. In juvenile plants, it is terete or flattened; in mature plants, flat, up to 4–5 mm wide. Rhizoidal holdfast develops from the cone-shaped base of the stipe. Additional rhizoids are formed upper on the stipe, on both its sides. Together, they form a loose, multilayered bundle. The texture of the rhizoids is fibrous. Rhizoidal outgrowths are once or twice branched, mainly in the lower third. Sporangial sori are initially formed as small, shapeless patches. As the soporiferous tissue matures, they merge and cover a significant part of the blade.
Examined specimens. Sea of Japan, southern Primorsky Krai: Posyet Bay, Troitsa Cove, depth 12 m, three specimens, Oct. 4, 2020, collected by N.G. Klochkova; Ussuri Bay, Shamora Cove, Feb. 16, 1971, beach casts, collected by T. Glushchenko; Peter the Great Bay, Bolshoi Pelis Island, beach casts, Aug. 13, 1987, two specimens, collected by N.G. Klochkova; Naumov Island, depth 8 m, May 18, 1966, one specimen, collected by M.V. Sukhoveeva (Fig. 3b). Tatar Strait: Frideriks Cove, beach casts, Aug. 29, 1973, one specimen, collected by N.G. Klochkova. Sea of Okhotsk: Sakhalin Island, Gulf of Patience, vicinities of Poronaysk Town, beach casts, July 28, 1993, nine specimens, collected by E.A. Khaliman; Gulf of Patience, Cape Pyata, depth 10 m, stony sediments, July 29, 1984, two specimens, collected by L.A. Chumakova (Figs. 3d, 3e); Aniva Bay, Busse Lagoon, depth 3 m, Aug. 8, 1993, two specimens, collected by E.A. Khaliman; Aniva Bay, depth 21 m, July 7, 1991, one specimen, collected by E.A. Khaliman (Figs. 3a, 3c). Shantar Islands, Feklistov Island, depth 19 m, two specimens, Sept. 2, 1986, collected by A.F. Zhmakin. Kuril Islands: Kunashir Island, Izmeny Bay, depth 7 m, coarse pebbles, sand, May 30, 1971, one specimen, collected by L.T. Prudnikova.
The examination of the Agarum specimens deposited in the herbaria of LE and SAP and other research organizations has shown that in the Far Eastern part of the range the species of this genus differ in their distribution patterns. A. undulatum has a very limited insular range, as can be seen in Fig. 4. A. turneri has a wider insular range. Until recent years, the latter has been considered as a cold temperate Asian-American species. In the recently published work of Klochkova and Klimova [3], it is reported also as distributed all along the Kuril Chain. The margin of its range covers the Nemuro Province coast, northeastern Hokkaido.

Distribution of the members of the genus Agarum in the Far Eastern seas. Black symbols indicate the type habitats of the species.
DISCUSSION
According to the latest taxonomic revision of the genus Agarum [9], it comprises the species A. clathratum, A. turneri, and A. yakishiriense. The type species of the genus Agarum is A. clathratum. The independence of A. turneri was long questioned, and it was considered a synonym of A. clathratum [4, 17] until a molecular genetic study has shown that these species significantly differ in rbcL and ITS2 [8].
In the reconstructed molecular phylogenetic tree, A. turneri forms a separate branch (Fig. 1a). It differs from A. clathratum by more than 4.5% (Table 1). This quite evident difference in the rbcL and cox1 nucleotide sequences between A. turneri and other members of the genus indicates its taxonomic independence. It is worth noting that in our extensive collection of specimens of this species, obtained from different sites of its distribution range, there are a few plants that, like A. clathratum subsp. yakishiriense, have blade perforations with scalloped edges slightly bent towards the ventral side. The factors causing their formation— habitat conditions or genetic differences—still remain unclear.
The issue of independence of A. yakishiriense has been addressed in different ways to date. A. yakishiriense was first described as a distinct species by Y. Yamada based on materials collected from off small islands (Yakishiri and Teuri) near Hokkaido [20]. Subsequently, I. Yamada [21] referred to it as one of the forms of A. cribrosum (=A. clathratum). In a monographic review of laminariacean algae of Japan, Kawashima [10] shared the opinion of I. Yamada and indicated A. yakishiriense as a form of the species A. clathratum.
In 2011, the status of this form was changed to the subspecies level after a comparative study of ITS2 + cox1 + cox3 in Agaraceae, including A. yakishiriense [7]. In this work of Boo and co-authors [7], the analyzed specimens of A. clathratum formed two clades. One of them, morphologically close to the type form of the species, comprised plants collected from off Hokkaido, the Kuril Islands, Kamchatka, Alaska, and the state of Minnesota. The other clade included specimens with a morphology characteristic of the form A. clathratum f. yakishiriense, collected from off the Korean Peninsula, Hokkaido Island, and Sakhalin Island.
From the viewpoint of phenotypic systematics, it should be recognized that the specimens described as A. yakishiriense are, indeed, characterized by a set of morphological traits that distinguish them from the other known members of the genus. Nevertheless, Kawashima [10] noted their close relationship with f. rishiriense and f. rugosum, which are the forms of the species described by I. Yamada [21] for A. cribrosum (=A. clathratum), and pointed out that all the forms distinguished by I. Yamada, including f. yakishiriense, occur in the same coastal waters off Japan.
In a study conducted by Kawai with co-authors [9], A. yakishiriense was not included in the multigene analysis, but was repeatedly mentioned as an independent species in the discussion of the study results and the phylogeny of Agaraceae. Nevertheless, the authors of the article noted that, in morphological terms, the species holds an intermediate position between the Agarum and Neoagarum species, since it is close to A. clathratum f. rugosum and A. clathratum f. rishiriense in the blade structure and to N. oharaense and N. fimbriatum in the stipe structure. The authors also pointed out that further research is needed to clarify the taxonomic position of A. yakishiriense [9].
In the above-presented molecular phylogenetic trees (Fig. 1), inferred from combined data of both rbcL + cox1 and ITS2 + rbcL + cox1, A. clathratum clearly splits into two clades: A. clathratum subsp. clathratum and A. clathratum subsp. yakishiriense. The former consists of two groups of specimens. One of them includes representatives of American populations from the Atlantic and North Pacific, and also a specimen collected off southeastern Kamchatka; the other is comprised of specimens collected from off the middle Kuril Islands and Hokkaido. The maximum difference in nucleotide sequences between the plants of the type form is 0.23% (Table 1).
The latter clade of A. clathratum subsp. yakishiriense is formed by plants collected in South Korea and off the coasts of Hokkaido and Sakhalin. The differences in ITS2 + rbcL + cox1 between the representatives of this group reach 0.9% due to the difference in ITS2. The maximum difference between the two subspecies of A. clathratum constitutes 1.69%. Due to the high internal variability in the subspecies yakishiriense, we suggest this difference is insufficient to consider it as an independent species. Another obstacle is also the high overlap of morphological characters between representatives of different subspecies. Thus, the specimens of A. clathratum subsp. yakishiriense from Hokkaido, Sakhalin, and southern Primorsky Krai have stipes with additional rhizoids always flat and very short, whereas its representatives from Chikhachev Bay and off the Shantar Islands have more elongated and terete stipes with less fibrous rhizoids. The belonging of the plants collected from the latter of the above-mentioned sites to the subspecies yakishiriense is confirmed by the study of Boo with co-authors [7].
When comparing the morphological traits characteristic of the considered taxa of the genus Agarum, it becomes clear that some of them overlap in several pairs of species (Table 2). Furthermore, each of them has its unique features of morphological organization. Thus, both subspecies of A. clathratum differ from A. turneri by a narrower, convex midrib, a thinner and lighter blade, and smaller sizes of perforations. The species A. turneri differs from A. clathratum subsp. yakishiriense in the stipe structure. In the former, stipe is terete and always quite long.
A. undulatum is distinguished by the most unusual morphology in the genus Agarum (Fig. 2). It is the only species with the ribbed stipe, the crenate blade margin, and bullations instead of perforations developed on blade. Merged with one another, bullations cover the entire surface of blade on both sides. No other species of the family Agaraceae has such morphological traits.
The genetic differences between A. undulatum and other species are also well expressed. In the maximum likelihood trees (Fig. 1), it clearly forms a separate branch. Thus, its differences in ITS2+rbcL+cox1 from A. clathratum constitute 5.51%; from A. turneri, 7.5%; and the difference from N. fimbriatum, a species of the genus Neoagarum, reaches the highest value, 9.58%. Thus, the phenotypic and genotypic traits of A. undulatum distinguish it well from the other representatives of the genera Agarum and Neoagarum. This gives us reason to describe this species as new to science.
REFERENCES
Belous, O.S., Titlyanova, T.V., and Titlyanov, E.A., An inventory of benthic flora in the intertidal and upper subtidal zones of Troitsa Bay (Peter the Great Bay, Sea of Japan), Russ. J. Mar. Biol., 2020, vol. 46, no. 6, pp. 472–484.
Klochkova, N.G., Koroleva, T.N., and Kusidi, A.E., Atlas vodoroslei-makrofitov prikamchatskikh vod (Atlas of Macrophytic Algae from Waters off Kamchatka), Petropavlovsk-Kamchatsky: KamchatNIRO, 2009, vol. 1.
Klochkova, T.A. and Klimova, A.V., The genus Agarum (Laminariaceae, Phaeophyceae) in the Far Eastern seas: Clarification of the species composition and distribution, in Materialy XIII mezhdunardnoi nauchnoi konferentsii “Sokhraneniye bioraznoobraziya Kamchatki i prilegayushchikh morei” (Conservation of the Biodiversity of Kamchatka and Adjacent Seas, Proc. XIII Int. Sci. Conf.), Petropavlovsk-Kamchatsky: Kamchatpress, 2020, pp. 214–217.
Petrov, Yu.E., Laminariales and Fucales algae from the seas of the Sobiet Union: Morphology, ecology, phylogeny, and systematics, Extended Abstract of Cand. Sci. (Biol.) Dissertation, Leningrad: Komarov Bot. Inst., USSR Acad. Sci., 1975.
Resursy i ratsional’noye ispol’zovaniye morskikh vodoroslei i trav dal’nevostochnykh morei Rossii (Resources and Rational Use of Marine Algae and Seaweeds from the Far Eastern Seas of Russia), Vladivostok: TINRO, 2020.
Skriptsova, A.V., Seaweeds of the Peter the Great Bay, Sea of Japan, Biota Sreda Zapov. Territ., 2019, no. 3, pp. 14–52.
Boo, G.H., Lindstrom, S.C., Klochkova, N.G., Yotsukura, N., Yang, E.C., Kim, H.G., Waaland, J.R., Cho, G.Y., Miller, K.A., and Boo, S.M., Taxonomy and biogeography of Agarum and Thalassiophyllum (Laminariaceae, Phaeophyceae) based on sequences of nuclear, mitochondrial, and plastid markers, Taxon, 2011, vol. 60, pp. 831–840.
Boo, S.-M. and Yoon, H.-S., Molecular relationship of giant kelp (Phaeophyceae), Algae, 2000, vol. 15, no. 1, pp. 13–16.
Kawai, H., Hanyuda, T., Gao, X., Terauchi, M., Miyata, M., Lindstrom, S.C., Klochkova, N.G., and Miller, K.A., Taxonomic revision of the Agaraceae with a description of Neoagarum gen. nov. and reinstatement of Thalassiophyllum, J. Phycol., 2017, vol. 53, no. 2, pp. 261–270.
Kawashima, S., Laminariaceaen Algae of Japan, Muroran, Japan: Hokkaido Univ. Publ., 1993.
Klimova, A.V., Klochkova, N.G., Klochkova, T.A., and Kim, G.H., Morphological and molecular identification of Alaria paradisea (Phaeophyceae, Laminariaceae) from the Kurile Islands, Algae, 2018, vol. 33, no. 1, pp. 37–48.
Klochkova, T.A., Kim, G.H., Belij, M.N., and Klochkova, N.G., Morphology and phytogeography of Laminaria appressirhiza and L. inclinatorhiza (Phaeophyceae) from the Sea of Okhotsk, Algae, 2012, vol. 27, no. 3, pp. 139–153.
Klochkova, T.A., Klochkova, N.G., Yotsukura, N., and Kim, G.H., Morphological, molecular and chromosomal identification of dwarf haploid parthenosporophytes of Tauya basicrassa (Phaeophyceae, Laminariaceae) from the Sea of Okhotsk, Algae, 2017, vol. 32, no. 1, pp. 15–28.
Lane, C.E., Lindstrom, S.C., and Saunders, G.W., A molecular assessment of northeast Pacific Alaria species (Laminariaceae, Phaeophyceae) with reference to the utility of DNA barcoding, Mol. Phylogenet. Evol., 2007, vol. 44, pp. 634–648.
Lindeberg, M.R. and Lindstrom, S.C., Field Guide to Seaweeds of Alaska, Fairbanks, Alaska: Alaska Sea Grant College Program, 2021. http://www.seaweedsofalaska.com. Accessed February 25, 2021.
Myabe, K. and Nagai, M., Laminariaceae of the Kurile Islands, Trans. Sapporo Nat. Hist. Soc., 1933, vol. 13, no. 2, pp. 85–102.
Silva, P.C., Nomenclatural remarks on Agarum (Laminariaceae, Phaeophyceae), Jpn. J. Phycol., 1991, vol. 39, no. 3, pp. 217–221.
Saunders, G.W. and Druehl, L.D., Nucleotide sequences of the internal transcribed spacers and 5.8S rRNA genes from Alaria marginata and Postelsia palmaeformis (Phaeophyta: Laminariaceae), Mar. Biol., 1993, vol. 115, pp. 347–352.
Stamatakis, A., RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies, Bioinformatics, 2014, no. 30, pp. 1312–1313.
Yamada, Y., On the species of Agarum, Acta Phytotaxon. Geobot., 1962, vol. 20, pp. 275–279.
Yamada, I., Local variation in Agarum cribrosum Bory (Phaeophyta) on the coasts of Hokkaido and adjacent regions, J. Fac. Sci., Hokkaido Univ., Ser. 5, 1974, vol. 10, pp. 32–47.
Huelsenbeck, J.P. and Ronquist, F., MRBAYES: Bayesian inference of phylogenetic trees, Bioinformatics, 2001, vol. 17, pp. 754–755.
ACKNOWLEDGMENTS
We are grateful to N.K. Khristoforova (Dr. Sci. Biol., Far Eastern Federal University), who provided us with the collection of algae gathered in 1986 during the hydrobiological expedition conducted by the Zhirmunsky Institute of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, off the Kuril Islands. We also thank our colleagues, phycologists V.N. Kulepanov (Cand. Sci. Biol.) and L.A. Balkonskaya (Cand. Sci. Biol.), who provided specimens of Agarum algae from the collections of the Pacific Research Fisheries Center and its Sakhalin branch for our study.
Funding
The study was supported by the Russian Foundation for Basic Research (research project no. 19-04-00285 A).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
The article is published in the original.
Rights and permissions
About this article

Cite this article
Klochkova, T.A., Klimova, A.V., Yotsukura, N. et al. A Revision of the Genus Agarum (Laminariaceae, Phaeophyceae) from the Far Eastern Seas with a Description of Agarum undulatum sp. nov. from the Kuril Islands. Russ J Mar Biol 47, 364–372 (2021). https://doi.org/10.1134/S1063074021050059
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1063074021050059