
Abstract
The size–age structure, growth, and maturation of the spotted catfish Arius maculatus (Thunberg, 1792) from the Hau River estuary, Mekong Delta, Vietnam, were studied for the first time. A total of 460 fish specimens collected from January to March 2019 were examined. The annuli on the otoliths were visualized; their formation may be related with the alternation of the dry and rainy seasons. The male to female ratio in the catches was 1 : 1.81. The proportion of males decreased in the older age groups. The maximum total body length of the females was 21.9 cm; for the males, it was 23.7 cm. The maximum recorded age of females in catches was 7+; for males it was 8+. No inter-gender differences in growth were detected. The parameters of the von Bertalanffy equation for both sexes according to empirical data were as follows: L∞ = 28.3 cm; k = 0.15/yr; t0 = –1.03 yr; growth performance index, φ = 2.08. The coefficients of the relationship between mass and length were as follows: a = 0.0054; b = 3.22. Maturation began at a body length of 13 cm at an age of 3 yr and ended at a body length of 18 cm at an age of 6 yr; 50% of the individuals became mature at a body length of 15 cm at an age of 4 yr. No inter-gender differences in the timing of maturation were found. A. maculatus from the Mekong Delta is characterized by a low growth rate compared to other populations, which may be associated with the labile habitat conditions in the estuary zone.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
The Mekong is the largest river in Indochina, with its length reaching almost 4500 km and a drainage basin area of 810 000 km2. The Mekong River is characterized by exceptionally high productivity and a biodiversity comparable to those of the Amazon River basin [25]. Nevertheless, the Mekong River is exposed over its length to extremely high anthropogenic pressure associated with hydrological constructions, fishing, ship traffic, and rapid development of aquaculture, agriculture, and other industries in the Asia–Pacific Region. The hydrological regime in the river delta has drastically changed over the past 30 years due to the intensification of agricultural activities including, in particular, rice production [24]. The currently ongoing climate change contributes to the rapid transformation of the Mekong River ecosystem, which poses a risk of partial or even complete loss of its components. Therefore, studying these components is very important for addressing conservation issues.
Interest in the ecosystem of the Mekong River basin has increased significantly; the biological diversity and structure of ichthyocenes are actively studied, and inventory of fish fauna has been conducted [1, 7, 20]. However, the biology of some species receives much less attention.
The spotted catfish Arius maculatus (Thunberg, 1792) is a demersal species that inhabits the coastal waters of the Indo-West Pacific zoogeographic region of the World Ocean. It is distributed along the western and eastern coasts of India, Sri Lanka, Pakistan, Bangladesh, and Myanmar to the Arafura Sea and the Indo-Australian archipelago [18]. In the Mekong River, the spotted catfish occurs in the lower reaches [5, 17, 19], where it is a target species of local commercial fisheries.
The biology of A. maculatus remains extremely poorly studied. Some data on the growth, age structure, mortality rate, and weight-to-length relationships are known for populations of this species off the coasts of Taiwan [8, 9], Indonesia [21], and Malaysia [15]. No data on the biology of A. maculatus in the Mekong estuarine system are available in the literature.
The study of age and growth has both theoretical and applied significance. The ecological and geographical variation in these parameters, which allows identification of the major adaptation and morphogenesis patterns, is of particular interest. However, the lack of knowledge of the biological characteristics of the population makes it impossible to assess its condition and production capacities and, therefore, organize rational fishing and nature conservation measures. The goal of the present work is to study the distribution, age structure, and growth and maturation characteristics of the spotted catfish A. maculatus in the Mekong Delta.
MATERIALS AND METHODS
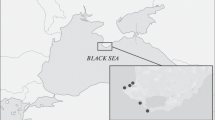
The study area included the Hau and Tien rivers, which are the largest branches of the Mekong Delta (Fig. 1). In January–March 2019, a total of 133 trawl hauls were performed in this water area. A beam trawl (with a frame width of 4 m; height, 40 cm; mesh size of the trawl bag, 10 mm) was used as fishing gear. The salinity of the near-bottom water layer was measured with a HM Digital TDS-3 portable total dissolved solid meter at each station.

A schematic map of the study area (outlined by rectangle). Black circles indicate the sites of capture of the spotted catfish Arius maculatus.
Complete biological analysis of the material was carried out by the standard methods [3]. The total length (TL) of fish was measured to an accuracy of 0.1 cm, and body weight (W) to 0.01 g. The age of individuals was estimated using otoliths. An annual ring (annulus) was considered as the boundary between the inner edge of a wide opaque (light in reflected light) zone (corresponding to a high growth rate) and the outer edge of a narrow translucent (dark in reflected light) zone (corresponding to a low growth rate). Otoliths were measured under 15× magnification. The otolith length (OL) was measured as the maximum distance between the rostral and caudal edges to an accuracy of 0.01 mm (Fig. 2). The annulus length was determined as the distance between the rostral and caudal edges of a translucent zone. The size of the studied sample was 460 specimens.

An otolith of a female Arius maculatus aged 4+ yr in incident light, TL = 166 mm. OL is the otolith length; dots indicate annuli.
The following statistical tests were used to describe the size–age structure: the Fisher’s F-test to assess the regression quality; the Shapiro–Wilk W-test to test for normality of distribution; the Mann–Whitney U-test to assess differences in length and weight between samples of females and males in catches; and the chi-squared (χ2) test to assess the sex ratio. Statistical analysis was carried out using the MS Excel and Statistica 12 software packages.
Back calculations were performed using the following formula: Li = (Si(Lc – c)/Sc) + c, where Si is the length of the annulus (mm) corresponding to the ith age; Lc is the TL of fish at capture, cm; Sc is the otolith length (OL, mm) at capture; and c is the free term of the linear regression of TL from OL [14]. To analyze the linear growth and weight increase, the following von Bertalanffy equations were used [2, 4, 22]: L = L∞(1 – \({{e}^{{ - k(t\,\, - \,\,{{t}_{0}})}}}\)) and W = W∞(1 – \({{e}^{{ - k(t\,\, - \,\,{{t}_{0}})}}}\))b, where L∞ is the asymptotic length, cm; W∞ is the asymptotic weight, g; k is a constant that characterizes the rate of L approximation to L∞; t0 is the age of fish with the length and weight in the considered model assumed to be 0, yr; and b is the power index of an equation where weight is a function of length (W = aLb). The growth performance index for linear sizes and weight was calculated as follows [16]: φ = log k + 2 log L∞ and φ′ = log k + (2 log W∞)/3.
RESULTS
Distribution. Within the area of the Hau River basin, the spotted catfish A. maculatus was found mainly in the estuarine zone with a salinity of the near-bottom water layer of 5‰ or higher. Single specimens were recorded from the area off the cities of Can Tho and Long Xuyen, where the water salinity was not higher than 0.8‰. The major part of individuals were caught from a depth of 5–15 m during trawling in the main channel of the river. The spotted catfish was found to tend to hard clayey sediments.
Age. Otoliths of A. maculatus had well-expressed opaque and hyaline zones (Fig. 2). Significant differences in the length of conditional annuli were observed in the first five groups (Fig. 3). The annuli were characterized by regular formation associated with the effect of cyclically varying environmental factors. The formation of the opaque zone, which corresponds to a high growth rate is completed by the end of the dry season. The slowdown of the otolith growth is timed to the rainy season and is accompanied by the formation of a narrow translucent zone in otoliths.

The annulus length in Arius maculatus from the Hau River estuary.
The sex ratio and size–age structure of catches. The total male to female ratio in catches was 1 : 1.81, which was statistically significantly different from the equal sex ratio (χ2 = 16.4; df = 1; p < 0.001). As the age of the fish increased, the sex ratio changed. In the first years of life, it was close to 1 : 1; in the older age groups, the proportion of males decreased. It should be noted that the largest and “oldest” fish in the catches was a male (TL = 23.7 cm; body weight 201 g; age 8+).
The linear sizes of A. maculatus females varied from 12.0 to 21.9 cm and those of males varied from 11.9 to 23.7 cm, with the mean TL values of 15.78 ± 0.18 and 14.85 ± 0.23 cm, respectively (Fig. 4a). In the catches, the length–frequency distribution differed from the normal one for both females and males (Shapiro–Wilk W-test: nfemales = 127, p = 0.002; nmales = 71, p < 0.001). The inter-gender differences in mean TL were statistically significant (Mann–Whitney U-test: nfemales = 127, nmales = 71, p < 0.001), which is associated with the dominance of females in the older age groups. The mean TL value of all mature individuals (n = 198) was 15.45 ± 0.15 cm.

The linear size (a), weight (b), and age (c) in the mature part of Arius maculatus catches from the Hau River estuary.
The body weight of females varied within a range of 16.0–117.4 g and that of males varied within 14.1–201.2 g, with the mean values of 41.6 ± 1.8 and 35.3 ± 2.8 g, respectively (Fig. 4b). The body weight–frequency distribution of both females and males also differed from the normal one (Shapiro–Wilk W-test: nfemales = 127, nmales = 71, p < 0.001). The differences in mean body weight between the sexes were significant (Mann–Whitney U-test: nfemales = 127, nmales = 71, p < 0.05) due to the predominance of older females. The mean body weight of all mature individuals (n = 198) was 39.3 ± 1.51 g.
It should be noted that immature individuals dominated the catches (51%) due to the specifics of the fishing gear; the proportion of young-of-the-year was 29%. The age–frequency distribution of the mature part of A. maculatus catches is shown in Fig. 4c. The maximum age of females was 7 yr; for males, it was 8+. The proportion of males aged 4+ was almost two times lower than that of males aged 3+. Unlike females, males at ages of 6+ and 7+ were not found in catches; therefore, the capture of a male at age 8+ is considered a single successful case.
An age–length key was compiled for the spotted catfish using the results of age estimation of 206 specimens (Table 1); on the basis of the key, the ages of another 254 individuals of mainly the younger age groups (0+ and 1+) were also determined.
The relationship of TL (cm) and otolith length OL (mm) in A. maculatus is approximated by a linear regression as follows: TL = 2.28OL + 2.12 with R2 = 0.95 (n = 214). The relationship is statistically significant (F-test, p < 0.05).
Since the equation allows calculating the length of only an averaged individual, we used an approach that takes into account the individual OL and TL values, on the one hand, and the free term in the regression equation, on the other (see the Materials and Methods section).
According to the empirical data, the growth of A. maculates can be provisionally divided into two stages (Table 2). The stage of rapid growth occurs in the first three years of life until the onset of maturation. The latter occurs when fish reach 3 years of age (at a body length of 13 cm and a body weight of 21 g), which marks the onset of the second stage. This stage is characterized by linearly decreasing growth and an allometric weight increase (in the equation of body weight to length relationship, b = 3.19 in females and b = 3.12 in males). The maturation of females and males ends at a body length of 18 cm and at age 6 yr.
The Ford–Walford plot [23] of the observed length values for both sexes has the following form: L(t+1) = 0.79L(t) + 4.75 (regression is statistically significant: F-test, p < 0.05). Its value at the intersection with the straight line L(t+1) = Lt is close to the estimated L∞ and is 22.58 cm, which indicates the high significance of the approximation of the empirical growth curve by the von Bertalanffy equation (Table 3).
According to the coefficients of the von Bertalanffy equation, by an age of 8–9 yr, both males and females reach 80% of the maximum length. According to the empirical rule [16], the mean maximum length is 95% of L∞, i.e., 24 cm for females and 22 cm for males, while the limit age for both sexes is 15 years. The mean values of the calculated lengths for the age groups were slightly lower than the empirical ones (Fig. 5); the discrepancy in the models is explained by the effect of the Lee’s phenomenon [4]. In both cases the growth performance index φ almost did not differ.

The linear growth curves for female and male Arius maculatus from the Hau River estuary described by the von Bertalanffy equation using the empirical and calculated data.
DISCUSSION
An analysis of the distribution of the spotted catfish A. maculatus in the study region has shown that it is an estuarine resident that does not make long migrations and avoids significantly freshened waters. However, local movements of fish within the estuary and marine coastal waters are also possible, especially during the rainy season.
Estimating the age of tropical fish usually poses some challenge. Waters of low latitudes are characterized by constant temperature conditions; therefore, fish of tropical species grow quite evenly throughout the year. However, in some cases, the growth of tropical fish during the year may be affected by the alternation of the dry and rainy seasons, when the hydrological and feeding conditions change [12]. The alternation of the periods of high and low growth rates in fish that are sensitive to these changes is naturally manifested in the age registering structures. The annual cyclicity in this case allows interpreting the boundary between the growth zones on otolith as an annulus. As an example, Fabré and Saint-Paul [11], in their work on the biology of the Amazonian anastomid fish Schizodon fasciatus, noted that the annulus on the scales of this species is formed during January–February. Thus, the maximum growth rate in this case occurs during the dry season at a low water level (July–November). In river systems, the annulus formation is typical for many Amazonian Cichla species, which is also associated with the cyclicity of the hydrological regime [13]. In water bodies where habitat conditions are stable throughout the year, no annulus formation is observed in fish. As in the above examples, the well-pronounced alternating opaque and hyaline zones on otoliths of A. maculates indicate a cyclical growth associated with the seasonality of the hydrological regime.
Unlike many fish species, A. maculatus does not show any pronounced inter-gender differences in the growth and timing of maturation. This may be explained by the exceptional role of males in the breeding process; they brood eggs in the mouth and continue to care for the offspring even after the embryonic development is completed. Large males can provide the survival of a larger number of offspring and, therefore, the increase in the size of males is an adaptive strategy fixed by selection. Some of the inter-gender differences are associated with the increase in weight of individuals of older age groups due to the predominant generative growth of females.
With a weakly expressed sexual dimorphism in growth, the proportion of males in the older age groups in catches was found to decrease. This may indicate a higher mortality rate in males or a lower catchability during fishing. Taking in account the care of offspring that is characteristic of male A. maculatus, an assumption can be made that they tend to stay in shelters.
A. maculatus that live in the Mekong Delta are characterized by a low growth rate compared to other populations and species of the genus Arius (Table 4). This may be due to their inhabiting the estuarine zone, which is an area with unstable hydrological conditions formed under the effect of freshening and changes in the thermal regime during the rainy season, as well as tidal and wind-induced phenomena. The hydrological instability causes an increase in energy expenditures for water–salt metabolism or for moving to areas with a more favorable hydrological regime. What advantages does this marine species gain by inhabiting an area with labile habitat conditions? By occupying the chorohaline zone, the spotted catfish probably avoids competition and/or predation pressure from the coastal zone of the sea, where the biological diversity is much higher. The low growth rate is compensated by a lower mortality rate, which allows maintaining the reproductive capacity. The occupation of freshened waters, where the direction and intensity of selection differ from those in the marine coastal zone, can lead to a phylogenetic shift and morphogenesis.
The new data obtained on the biology of A. maculatus from the Mekong Delta during the dry season can become a starting point in monitoring the status of its population. Of particular interest is the study on the distribution and biology of A. maculatus during rainy season. This will allow determination of the migration activity and the degree of relationship between members of the estuarine and marine coastal zones.
REFERENCES
Boltachev, A.R., Karpova, E.P., Statkevich, S.V., et al., Characteristics of quantitative distribution of fish and decapod crustaceans in the Mekong Delta during the low-water season of 2018, Morsk. Biol. Zh., 2018, vol. 3, no. 4, pp. 14–28.
Mina, M.V. and Klevezal, G.A., Rost zhivotnykh (Animals’ Growth), Moscow: Nauka, 1976.
Pravdin, I.F., Rukovodstvo po izucheniyu ryb (A Guide to the Study of Fish), Moscow: Pishchevaya Promyshlennost’, 1966.
Ricker, W.E., Computation and Interpretation of Biological Statistics of Fish Populations, Ottawa: Dep. Environ. Fish. Mar. Serv., 1975.
Serov, D.V., Polevoi opredelitel’ presnovodnykh ryb Yuzhnogo Vietnama (A Field Key to Freshwater Fishes of South Vietnam), Moscow: Nauka, 1994.
Bawazeer, A.S., The fishery management of the stock of chim, the giant sea catfish Arius thalassinus in Kuwait waters, Kuwait Bull. Mar. Sci., 1987, vol. 9, pp. 87–100.
Chea, R., Lek, S., Ngor, P., and Grenouillet, G., Large-scale patterns of fish diversity and assemblage structure in the longest tropical river in Asia, Ecol. Freshwater Fish, 2017, vol. 26, no. 4, pp. 575–585.
Chu, W.-S., Hou, Y.-Y., and Ueng, Y.-T., Estimates of age, growth and mortality of spotted catfish, Arius maculatus (Thunberg, 1792), off the Coast of Yunlin, Southwestern Taiwan, Afr. J. Biotechnol., 2011, vol. 10, no. 66, pp. 15416–15421.
Chu, W.-S., Hou, Y.-Y., Ueng, Y.-T., and Wang, J.-P., Correlation between the length and weight of Arius maculatus off the southwestern coast of Taiwan, Braz. Arch. Biol. Technol., 2012, vol. 55, no. 5, pp. 705–708.
Conand, F., Camara, S.B., and Domain, F., Age and growth of three species of Ariidae (Siluriformes) in coastal waters of Guinea, Bull. Mar. Sci., 1995, vol. 56, no. 1, pp. 58–67.
Fabré, N.N. and Saint-Paul, U., Annulus formation on scales and seasonal growth of the Central Amazonian anastomid Schizodon fasciatus, J. Fish Biol., 1998, vol. 53, pp. 1–11.
Henderson, P.A., The growth of tropical fishes, Fish Physiol., 2005, vol. 21, pp. 85–100.
Jepsen, D.B., Winemiller, K.O., Taphorn, D.C., and Rodrigues Olarte, D., Age structure and growth of peacock cichlids from rivers and reservoirs of Venezuela, J. Fish Biol., 1999, vol. 55, pp. 433–450.
Johdal, M.S., Esmaeili, H.R., and Tandon, K.K., A comparison of back-calculated lengths of silver carp derived from bony structures, J. Fish Biol., 2001, vol. 59, no. 6, pp. 1483–1493.
Mazlan, A.G., Abdullah, S., Shariman, M.G., and Arshad, A., On the biology and bioacoustic characteristic of spotted catfish Arius maculatus (Thunberg 1792) from the Malaysian Estuary, Res. J. Fish. Hydrobiol., 2008, vol. 3, pp. 63–70.
Pauly, D., Moreau, J., and Prein, M., A comparison of overall growth performance of tilapia in open waters and aquaculture, in The Second International Symposium on Tilapia in Aquaculture, Pullin, R.S.V., Bhukaswan, T., Tonguthai, K., and Maclean, J.L., Eds., ICLARM Conference Proceedings, vol. 15, Bangkok: Dep. Fish., 1988, pp. 469–479.
Rainboth, W.J., Fishes of the Cambodian Mekong, in FAO Species Identification Field Guide for Fishery Purposes, Rome: FAO, 1996.
Russell, B.C. and Houston, W., Offshore fishes of the Arafura Sea, Beagle: Rec. North. Territ. Mus. Arts Sci., 1989, vol. 6, pp. 69–84.
Tran, D.D., Shibukawa, K., Nguyen, T.P., et al., Fishes of the Mekong Delta, Vietnam, Can Tho, Vietnam: Can Tho Univ. Publ. House, 2013.
Valbo-Jørgensen, J., Coates, D., and Hortle, K., Chapter 8 – Fish diversity in the Mekong River basin, in The Mekong: Biophysical Environment of an International River Basin, New York: Academic, 2009, pp. 161–196.
Velasco, G. and Oddone, M.C., Growth parameters and growth performance indexes for some populations of marine catfishes (Actinopterygii, Siluriformes, Ariidae), Acta Biol. Leopold., 2004, vol. 28, pp. 307–313.
Von Bertalanffy, L., A quantitative theory of organic growth (Inquiries on growth laws. II), Human Biol., 1938, vol. 10, pp. 181–213.
Walford, L.A., A new graphic method of describing the growth of animals, Biol. Bull., 1946, vol. 90, no. 2, pp. 141–147.
Wassmann, R., Hien, N.X., Hoanh, C.T., and Tuong, T.P., Sea level rise affecting the Vietnamese Mekong Delta: water elevation in the flood season and implications for rice production, Clim. Change, 2004, vol. 66, nos. 1–2, pp. 89–107.
Ziv, G., Baran, E., Nam, S., et al., Trading-off fish biodiversity, food security, and hydropower in the Mekong River Basin, Proc. Natl. Acad. Sci. U. S. A., 2012, vol. 109, no. 15, pp. 5609–5614.
Funding
The study was carried out in the framework of the Ekolan E-3.4 theme (entitled The Impact of Climate Change and Anthropogenic Factors on the Ecosystems of the Lower Mekong River) and the State assignment to the Kovalevsky Institute of Biology of the Southern Seas, Russian Academy of Sciences (The Patterns of Formation and the Anthropogenic Transformation of Biodiversity and Bioresources of the Azov–Black Sea Basin and Other Areas of the World Ocean, no. AAAA-A18-118020890074-2).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by E. Shvetsov
Rights and permissions
About this article

Cite this article
Kutsyn, D.N., Ablyazov, E.R., Truong, B.H. et al. The Size–Age Structure, Growth, and Maturation of the Spotted Catfish Arius maculatus (Thunberg, 1792) (Siluriformes: Ariidae) from the Mekong Delta, Vietnam. Russ J Mar Biol 47, 56–63 (2021). https://doi.org/10.1134/S1063074021010053
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1063074021010053