
Abstract
Retinal pigment epithelium (RPE), whose main functions are to participate directly in light perception and ensure the trophic and functioning of the retina, is a traditional subject of numerous studies. A significant part of them is biomedical and is aimed at identifying the causes of aging and degeneration of RPE, which are, in turn, associated with several serious retinal diseases that lead to the loss of the eyesight. Rodents, in particular, the albino Wistar rats, are often used as the object of these studies. The work is aimed at identifying signs of aging in the RPE in young (from 2 weeks up to 2 months) albino rats. Using the methods of fluorescence analysis and Schmorl staining, the accumulation of the “aging pigment” lipofuscin in the isolated layer of RPE, as well as a high content of polyploid binuclear cells to mononuclear cells, were detected in albino rats of an early age. In parallel, the presence of lipofuscin was detected in the isolated vascular layer (choroid) of the retina that underlies RPE. The relationship between the early appearance of signs of aging in the RPE of albino rats and the absence of the melanin pigment in the cells of this tissue is discussed. The obtained data should be taken into account when conducting experimental ophthalmological studies on this animal model object.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Retinal pigment epithelium (RPE) is a monolayer of polarized and highly specialized pigmented cells. RPE is located between the neural retina and its vascular layer, which is called the choroid and supplies the retina with all the necessary substances and oxygen for its normal trophic function. RPE, in addition to transferring substances along the basal-apical cells' gradient, performs a number of important functions necessary for maintaining homeostasis and normal functioning of the retina. It is well known that the main function of RPE is phagocytosis of the exfoliating outer segments of retinal photoreceptors and then their digestion with the help of lysosomes (Strauss, 2005). This allows photoreceptors to synthesize new processes for light perception, ensuring cell viability and retinoid metabolism (Young et al., 1971; Mazzoni et al., 2014). However, complete digestion of the photoreceptor’s outer processes using lysosomal enzymes and their complete degradation by the RPE cells is not possible, which, as a result, leads gradually and steadily to the accumulation of lipofuscin (LF) (Katz, 1989; Gu et al., 2012). The outer segments of photoreceptor cells are rich in polyunsaturated fatty acids that are exposed to free radicals, which are found in large quantities in cells that function under constant light irradiation of the retina. As a result of peroxidation, which is catalyzed in lysosomes by metal ions, polyunsaturated fatty acids are transformed into fluorophores, autofluorescence components of LF (Gutteridge et al., 1982; Smith et al., 2016). It is also assumed that LF in RPE is a heterogeneous substance, which also includes fats and proteins, and the fluorescent components are different and the main one is identified as a derivative of vitamin A (Kennedy et al., 1995).
At the same time, it is known that, if RPE cells of adult animals in vivo have proliferative activity, then only on the periphery of the layer and at a very low level (Al-Hussaini et al., 2008; Grigoryan, 2014). For this reason, all accumulated LF remains in the cytoplasm, and the amount of it in the cells of RPE not only does not decrease but, on the contrary, increases. This process can be experimentally recreated when RPE is cultured in vitro under conditions of oxidative stress (Sundelin et al., 2001). The consequences of LF accumulation are degeneration of the RPE cells and impaired function of photoreceptor processes. Many diseases of the human retina are associated with degeneration and RPE dysfunction. These include macular degeneration of the retina (age-related macular degeneration, AMD), which often leads to blindness (Sparrow et al., 2010).
An increase in the relative number of multinuclear cells in the situation of a decrease in the total number of cells in the population is also related to age-related processes in RPE. The presence of mononuclear and polyploid, mainly binuclear cells, in the RPE layer is a common situation for rodents (Al-Hussaini et al., 2008). In old mice, for example, the proportion of multinuclear cells in RPE can reach 80%, and the vast majority of them are binuclear cells (Chen et al., 2016). Data of Chen et al. also indicate that cells, becoming binucleated, significantly increase in size, although they retain their phagocytic activity against the flaking disks of the outer segments of the photoreceptors. It is assumed that an important role in the acquisition of the RPE layer of multinuclear cells is played by an increasing phagocytic workload associated with the growth of the eye, which disrupts cell cytokinesis, and the multinuclearity of cells is a kind of protective mechanism for preserving the RPE layer and its main function (Chen et al., 2016).
It is known that the development of degenerative processes in RPE is often accompanied by the loss of cell melanin granules’ pigment (see review of Rozanowska, 2011). Melanin is a polymer of high molecular weight, synthesized and accumulated in the RPE during the development and growth of the eye. Melanin is formed as a result of the oxidation of precursor molecules tyrosine and dihydroxyphenylalanine (DOPA) and is contained in granules that fill the cytoplasm of RPE cells (Rozanowska, 2011). An important function of melanin, which is in high concentration in the apical cytoplasm and apical processes of RPE cells, is to protect photoreceptors from excessive light. It is assumed that the photoprotective role of melanin in RPE cells is due to its ability to bind redox-active metal ions (Sarna, 1992). LF is in quantitative competition with melanin: when there is more melanin in the cells of RPE, then there are fewer LF granules and vice versa (Sundelin et al., 2001).
Albinism is characterized by the destruction of melanosomes, where the pigment is synthesized and deposited, and/or by the violation of the process of synthesis of melanin (see the review of Grønskov et al., 2007; Summers, 2009). However, RPE and the neural retina of albino rodents are used in vivo (Novikova et al., 2013; Nadal-Nicolás et al., 2014; Iwai et al., 2016) and in vitro (Cortese et al., 2005; Grigoryan et al., 2012) as a model for studying the development of the retina and age-dependent processes that can lead to retinal degeneration. To conduct this type of research, additional characteristics of albino rodents RPE are needed, in particular, those related to the presence and accumulation of LF, as well as the development of multinuclear cells in the RPE layer, as another age-dependent process. In this work, we used whole amount preparations of RPE from albino rats to study these signs in the age range from 2 weeks to 2 months after birth.
MATERIALS AND METHODS
Object of Research
The work was performed on the RPE of young Wistar albino rats at the age of 2 weeks, 1 month, and 2 months after birth. The choice of the age of rats is due to our preliminary observation, which indicates that individual cells of these animals can contain a significant number of granules with the autofluorescence even after 1 month after birth. In vivarium conditions, all the rats were in the light mode of 12 h of light/dark and free access to food and water.
Isolation of Retinal Pigment Epithelium
After anesthesia, the eyes of albino rats aged 2 weeks, 1 month, and 2 months were enucleated. To study the RPE layer, we used (with modifications) the whole amount method of its isolation using dissociating media (Wang et al., 1993).
Immediately after isolation, the eyes were placed in a cooled DMEM medium and transferred to box conditions. The eyes were freed from muscle and connective tissue in the DMEM medium and washed twice in cold Hanks solution without calcium (HBSS, Gibco) containing EDTA (Helicon) at a concentration of 10 mmol/L, pH 7.4. Then, using binoculars, the eyes were divided into anterior and posterior sectors microsurgically, making the incision posteriorly outside the corneal limbus (Fig. 1). In this situation, the neural retina remained in the posterior wall of the eye. The posterior sectors of the eye bowls were then washed again three times in a cold Hanks solution with EDTA.

Scheme of the isolation of tissues of the back wall of the rat’s eye. (a) Back wall of the eye, (b) isolation of the neural retina, (c) isolation of retinal pigment epithelium, (d) vascular and scleral membranes remain. A detailed description of the method of tissue isolation is given in the article.
Then, the posterior sectors of rat eyes were incubated at 37°C for 10–12 min in Hank’s solution with EDTA, in which hyaluronidase type IV (220 units/mL, Merck) and collagenase type I (65 units/mL, PanEco) were added. The eye bowls were then again transferred to the cold Hank’s solution with EDTA, where the neural retina was isolated, while the RPE remained intact as part of the posterior sector of the eye. The posterior eye sectors were reincubated in the above-mentioned dissociating media for 8 min at 37°C and transferred to a cold Hank’s solution with EDTA, in which they were washed twice from collagenase and hyaluronidase enzymes.
After incubation in Hank’s solution with EDTA at room temperature (30 min) to achieve complete detachment of the RPE layer from the underlying tissues, it was carefully separated with thin glass rods with a sealed end (0.3–0.5 mm) along the edge, moving towards the center of the eye, to the area of the exit of the optic nerve. After separation of the RPE, it was taken away with a pipette and carefully transferred to glass slides, which were placed in a wide Petri dish in formalin vapors for soft fixation of the RPE tissue. In parallel, we were able to isolate fragments of the underlying vasculature from two-month-old rats and fix them in the same way.
Fixed preparations of the RPE of albino rats of different ages were used for further histological treatment with hematoxylin-eosin staining, Schmorl staining, and, without histological treatment, for the analysis of autofluorescence of LF granules on total RPE preparations using a fluorescent microscope. For staining according to the Schmorl method, additional transverse paraffin sections (7 microns) of the posterior wall of the eyes of rats of different ages, fixed in 4% formalin, were used.
Staining by Schmorl’s Method
After dewaxing, sections taken from the equatorial and central regions of the posterior wall of the eye, as well as fixed fragments of RPE layers of rats of different ages, were washed three times in distilled water. They were then placed in a freshly prepared staining medium containing 1% ferric chloride (30 mL), potassium hexacyanoferrate (4 mL), and distilled water (6 mL). The intensity of staining was monitored using a microscope in the range from 5 to 10 min of reaction with the dye. After that, the preparations were thoroughly washed in running water, dehydrated, placed in xylene, and embedded in Canada balsam under cover glasses.
Analysis of the Fluorescence of Lipofuscin Granules
LF is an autofluorescent pigment of glycolipoprotein nature. For fluorescence analysis, the RPE preparations were thoroughly washed 1 day after fixation and embedded in glycerol with the addition of DABCO (Sigma, United States), a fluorescence protector. The presence of LF granules was estimated by registering their fluorescence using a AxioVev + 200M fluorescence microscope (Carl Zeiss Jena, Germany). Images were obtained in the light short-wave range (488 nm) of an argon laser (570 nm barrier filter) and with other identical parameters of the microscope. The resulting images were recorded with a digital camera and a computer equipped with the AxioVision LE Rel.4.3 program.
Quantitative analysis of the content of LF granules in the RPE of albino rats of 2 weeks, 1 month, and 2 months of age was performed by visual counting the number of cells containing plenty of brightly fluorescent granules relative to the total number of cells on 20 visual fields of 16 × 104 px in images of RPE layers obtained at 1000-fold magnifications of the microscope (100× objective, 10× ocular). Cells with weak light granules on the edge of identification or with a small number of them were not taken into account. This deliberately lowered the level of LF content in the RPE, setting a goal to avoid significant errors in calculations and to give a comparative characteristic of the RPE cells of rats at different ages by the parameter of the relative number of fluorescent cells in the layer. Calculations were performed on black-and-white images of RPE preparations obtained from both eyes of two rats for each term. Statistical data processing was performed using Excel. The average values of the indicator for each age, the standard deviation, and the analysis of variance of samples were calculated.
Determining the Ratio of Mono- and Binuclear Cells in the RPE Layer
For this purpose, RPE samples from rats aged 2 weeks, 1 month, and 2 months, fixed in formalin vapors and hematoxylin-eosin-stained, were used. The calculations took into account only the equatorial region of the layer along its entire circumference; the periphery and central (around the optic nerve head) areas of the RPE remained outside the quantitative analysis. This is due to the data provided in the literature, according to which many cells on the periphery of RPE in 3-month-old albino Wistar rats often have atypical morphology and fuzzy borders (Adams et al., 2010). According to our observations, in the central region near the optic nerve head, the morphology and distribution of RPE cells are irregular, which also makes quantitative studies very complicated. For this reason, we did not consider the central area in our calculations. Such changes were absent in the equatorial region, which we have chosen; this region is a much larger area of the RPE layer; the approximate ratio from the extreme periphery to the center is 0.5/1.5/0.5 (in mm).
To perform cell counting, histological preparations of RPE were examined using an Olympus AH-3 microscope equipped with a digital camera. For each sample, a series of images of the equatorial zone of the RPE were taken. On each of the images obtained, rectangular areas were chosen, where we first counted binuclear and then mononuclear cells using the Image J program. After that, the ratio of the number of mononuclear cells to the number of binuclear cells was determined. We then performed statistical data processing using Excel. We calculated the average values of indicators for each age, standard deviations, and variance analysis of samples to determine the reliability of differences in the samples.
RESULTS AND DISCUSSION
The RPE of the normal eye of an 2-month-old adult albino is a single-row layer of hexagonal cells that are tightly adjacent to each other (Fig. 2a). RPE cells can be mononuclear or binuclear, and their ratio depends on the age of the animal (Chen et al., 2016). The majority of RPE cells in rats of the age of 1 month or more have two equal-sized nuclei, usually containing 1–2 nucleoli (Fig. 3a). In addition to binuclear cells, mononuclear cells are also present at a relatively low number, the size of the nuclei of which is often larger than that of binuclear cells (Figs. 3b, 3c, 3d). In the population of binuclear and mononuclear cells, there were also unusually large cells, about one and a half times larger in diameter as normal mononuclear cells. It is important to note that the cytoplasm of RPE cells in albino rats does not contain melanin granules and has a uniform homogeneous light color when using hematoxylin-eosin for staining. When compared with the pigmented RPE of C57Bl mice, which we analyzed as an additional sample, this difference is obvious (Figs. 4a, 4b). It is known that the main distinguishing feature of albino RPE in development is not only the absence of pigment but also the absence of the components of its synthesis, which are also involved in the regulation of cell cycles in the retina. For example, a low level of dihydroxyphenylalanine (DOPA), a precursor of melanin synthesis, can affect the process of cell exit from the cycle, which in turn can lead to both excessive proliferation and cell death (Ilia and Jeffery, 1999). Some authors (Adams et al., 2010) attribute this phenomenon to frequent cases of cytokinesis disorders and other abnormalities in the population of RPE cells, especially its peripheral division, which differentiates late in development and has a very low but greater, relative to the equatorial and central regions of the layer, proliferative activity (Al-Hussaini et al., 2008).

Transversal section of the posterior wall of the eye of an albino rat at the age of 2 months. (a) Normal structure, hematoxylin-eosin staining, (b) staining by the Schmorl method. The arrows indicate the region adjacent to each other the endings of the photoreceptor processes of the retinal pigment epithelium and the choroid.

Isolated layer of retinal pigment epithelium of an albino rat at the age of 2 months. (a) General view, (b, c, d) fragments at high resolutions of the microscope. Thin arrows indicate mononuclear cells; thick arrows indicate cells with large oval nuclei.

Retinal pigment epithelium of an adult C57Bl mouse at different microscopic resolutions.
This feature, the absence of melanin granules (or, perhaps, an extremely low number of them), which occurs in the model used, allows us to determine quite accurately the inclusions of LF in the cell cytoplasm that accumulate granules during phagocytosis. When studying the RPE fields belonging to different fields of the equatorial region of the layer, we did not reveal significant cellular morphological differences. Figures of mitotic divisions of nuclei in RPE in samples obtained from four eyes of two animals for each term were not found.
On sections of the posterior wall of the normal eye of albino rats, the RPE is located between the neural retina and the vascular membrane. It forms a single-row, nonpigmented, epithelium consisting of binuclear and mononuclear cells that are tightly adjacent to each other by lateral surfaces. On the basal side, the layer is lined with a Bruch’s membrane and a vascular (choroidal) membrane (Fig. 2a). These three layers are normally arranged densely to each other and together constitute an independent complex. The vascular membrane in rats has a developed heterogeneous structure, a vascular network consisting of blood vessels of different calibers. Vascular endothelial cells and “melanophore” cells with process and without pigment, fibroblasts, mast cells, and resident macrophages are also cellular elements of the vascular membrane (Fig. 5a). The vasculature is followed by a dense, scleral connective tissue that provides mechanical tensions and preserves the topology of the RPE and the vasculature.

Vasculature of the eye of an albino rat at the age of 1 month. (a) General view, hematoxylin–eosin staining, (b) staining by the Schmorl method.
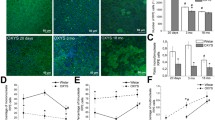
LF granules are autofluorescent, electron-dense, heterogeneous, polymer substances that are accumulated inside the lysosomal vacuolar apparatus (in secondary lysosomes) of cells. Accumulation occurs throughout life in a linear relationship with age and it is an indicator (biomarker) of aging (Katz, 1989). The detection of LF using a fluorescent microscope and the subsequent quantitative analysis of the relative number of cells with significant amounts of LF, both by the number of granules and their fluorescence, allowed us to judge the accumulation of LF in the cells of the RPE of albino rats over a period from 2 weeks to 2 months after birth. These results are shown in Figs. 6 and 7. It should be noted that, with the higher probability, in the population of rats of the same age, there are differences in the number of intensely fluorescent cells in the RPE, which is most likely due to the individual characteristics of the animals. However, we can still identify certain rules. The first is that LF-containing cells are detected already in 2 weeks of postnatal development in albino rats in the equatorial zone of RPE, but the relative content of them does not exceed 10%. LF fluorescence is most often diffused. Cells with a fluorescent cytoplasm can be detected both in groups and individual cells.

Autofluorescence in retinal pigment epithelium cells of albino rats (a) 2 weeks, (b) 1 month, and (c) 2 months after birth.

Increase in the number of cells with high LF fluorescence in the retinal pigment epithelium of albino rats in early development. The average number of cells with a high level of fluorescence relative to the number of all cells in the analyzed image field of the RPE layer is on the ordinate axis.
Later, by 1 month of development, the number of such cells increases significantly and reaches 17–20% by 2 months (Fig. 7). Intergroup analysis revealed significant differences between the average values of the indicator when comparing the ages of 2 weeks and 1 month (p < 0.01). As the relative number of LF-containing cells increases, the variation of this indicator in the samples also increases. Thus, it is obvious that the accumulation of LF in the RPE of the eyes of albino rats during aging is a natural process for these animals that takes place soon after birth and occurs quite quickly in a short time interval between 2 weeks and 2 months. What is the reason for such a rapid appearance of this sign of aging? There are works that directly indicate a feedback link between the presence of melanin (melanosomes) and the accumulation of LF in the cells of RPE. Sundelin et al. (2001) studied rabbit and bovine RPE under conditions of oxidative stress in vitro caused by the addition of external segments of photoreceptor cells to the RPE cells. The authors compared the RPE of normal rabbits and albinos as well as bovine RPE cells rich and depleted with melanin granules. It was shown that RPE cells of albino rabbits and bovine RPE cells with a low content of melanin accumulated significantly higher amounts of LF compared to the LF content in RPE cells of normal rabbits as well as in bovine RPE cells rich in melanin. This work, according to the authors opinion (Sundelin et al., 2001), showed that melanin is an effective antioxidant, which, in turn, is probably due to the connection of melanin with the metal transfer process. It is also assumed that the absence of melanin pigment in the RPE may cause a violation of intercellular communication carried out through contacts not only on the lateral surfaces of RPE cells but also with the neural retina as it develops, which leads to a violation of cellular integration (Iwai et al., 2016). This can manifest itself in changes in the shape of cells as well as changes in the process of phosphorylation and assembly of cell contact proteins.
The obtained RPE samples of albino rats, stained using the Schmorl method for LF detecting, made it possible to conduct an additional analysis of RPE in the developing albino rat. Quantitative analysis with this approach is limited due to the peculiarities of any histological staining, the intensity of which depends on many factors. However, the method made it possible to detect colored (dark-blue) inclusions both on the cross sections of the posterior wall of the rat eye (Fig. 2b) and on whole amount RPE preparations (Figs. 8a, 8b, 8c, 8d). On traversal sections, the colored inclusions were localized in the RPE cells at the border with retinal photoreceptors as well as in the vascular membrane (Fig. 2b). This localization of staining was more obvious in rats at the age of 2 months than in rats of an earlier age. In the early stages of development (2 weeks and 1 month), the concentration was insignificant, often on the verge of background of dye inclusion (not shown). In general, these results were consistent with the data obtained when assessing the accumulation of LF using a fluorescent microscope, indicating the overall presence of LF at all follow-up periods, and with a maximum of 20% in 2-month-old albino rats.

Presence of LF granules in isolated retinal pigment epithelium. (a) Hematoxylin-eosin staining, 1 month, (b) Schmorl staining, 1 month, (c) hematoxylin-eosin staining, 2 months, (d) Schmorl staining, 2 months.
It is interesting that, in addition to RPE, in the posterior wall of the eye of 2-month-old rats, specific staining is also present in the vascular membrane (Fig. 5b). The presence of LF in the vasculature on paraffin sections stained by the Schmorl method was also noted for earlier ages. Recently, an accumulation of LF with age as a result of lipid peroxidation in human retinal vessels has been described (Nag et al., 2019).
Thus, the early accumulation of LF in RPE cells revealed in the study and the vasculature may be evidence of one of the signs of RPE aging in albino rats already at an early postnatal age. Accumulation of LF was also shown for the tissue of the lacrimal glands of aging (aged from 2 to 24 months) rats. The authors (Batista et al., 2011) showed that the accumulation of LF occurs in the case of a decrease in the expression of various antioxidant markers, which indicates the suppression of antioxidant protection with age, which in turn leads, in particular, to the accumulation of LF and other signs of aging.
The results of determination of the mononuclear to binuclear cell ratio in the equatorial region of the RPE layer of rats of different ages are shown in Fig. 9. It follows from the histogram data that there is still a significant number of mononuclear cells in the equatorial zone of RPE after 2 weeks of postnatal development of albino rats; however, by the first month of postnatal development of rats, their content significantly decreases, and this trend persists later. Intergroup analysis revealed significant differences between the final average values of the ratio of mononuclear and binuclear cells when comparing 2-week-old and 1-month-old rats (p < 0.005).

Change in the ratio of mononuclear and binuclear cells in the retinal pigment epithelium of albino rats at an early age. The ratio of mononuclear cells to the number of binuclear cells in the analyzed image field of the layer of RPE is on the ordinate axis.
It is known from the literature that the RPE of pigmented rodents accumulates binuclear cells as early as the third postnatal week (Bodenstein and Sidman, 1987). According to our data, albino rats have a significant number of such cells in the RPE after 2 weeks of postnatal development, which, in turn, may be due to one of the factors of albinism, insufficient levels of DOPA. The last is, in turn, responsible for the exit of the cell from the cycle and, probably, cytokinesis (Adams et al., 2010).
An interesting question is a connection between the accumulation of LF by RPE cells and the development of multinuclearity. According to our observations, the accumulation of LF occurs in both binuclear and mononuclear cells, but the question on the relationship between the rate of accumulation of LF and the disturbance of cytokinesis leading to multinucleation cannot be answered since the number of mononuclear cells at all periods of observation is small. However, when comparing the quantitative results presented in Figs. 7 and 9, we can find a positive correlation between these two events.
The data obtained on RPE of early age albino rats indirectly confirm and complement information about the role of melanin in RPE cells as a necessary participant in maintaining homeostasis, protecting against excessive light, and developing and aging of the retina. The appearance of such signs as the presence and accumulation of LF, as well as the presence of a high relative number of binuclear cells in the RPE in albino rats in early postnatal development are doubtlessly associated with the absence of melanin. Both processes also indicate early aging of RPE in albino rats, which should be taken into account when conducting ophthalmological medical and biological studies on this animal model object.
CONCLUSIONS
The RPE of the albino rat eye already shows signs of aging in the early stages of postnatal development. This is expressed in the accumulation of lipofuscin granules by cells as well as in an increase in the relative content of binuclear polyploid cells in the layer. In the first 2 weeks of postnatal development, these signs are expressed insignificantly, but they become obvious by a month. The reason for the phenomenon of early aging of RPE in albino rats is the absence of melanin, which protects against over-oxidation, and components of the synthesis of melanin, regulators of cytokinesis. These data complement the data on the state of RPE of albino rats used in experiments in vivo and in vitro as an animal model in the study of aging and retinal pathology.
REFERENCES
Adams, T., Shahabi, G., Hoh-Kam, J., and Jeffery, G., Held under arrest: many mature albino rpe cells display polyploidal features consistent with abnormal cell cycle retention, Exp. Eye Res., 2010, vol. 90, no. 2, pp. 368–337.
Al-Hussaini, H., Vugler, A., Semo, M., and Jeffery, G., Mature mammalian retinal pigment epithelium cells proliferate in vivo, Mol. Vision, 2008, vol. 14, pp. 1784–1791.
Batista, T.M., Tomiyoshi, L.M., Dias, A.C., et al., Age-dependent changes in rat lacrimal gland anti-oxidant and vesicular related protein expression profiles, Mol. Vision, 2012, vol. 18, pp. 194–202.
Bodenstein, L. and Sideman, R., Growth and development of the mouse retinal pigment epithelium. Cell and tissue morphometrics and topography of mitotic activity, Dev. Biol., 1987, vol. 121, pp. 192–204.
Chen, M., Rajapakse, D., Fraczek, M., et al., Retinal pigment epithelial cell multinucleation in the aging eye—a mechanism to repair damage and maintain homoeostasis, Aging Cell, 2016, vol. 15, pp. 436–445.
Cortese, K., Giordano, F., Surace, E.M., et al., The ocular albinism type 1 (OA1) gene controls melanosome maturation and size, Invest. Ophthalmol. Vis. Sci., 2005, vol. 46, no. 12, pp. 4358–4364.
Grigoryan, E.N., Competence factors of retinal pigment epithelium cells for reprogramming in the neuronal direction during retinal regeneration in newts, Izv. Ross. Akad. Nauk, Ser. Biol., 2014, no. 1, pp. 5–16.
Grigoryan, E.N., Novikova, Y.P., and Gancharova, O.S., New antioxidant SkQ1 is an effective protector of rat eye retinal pigment epithelium and choroid under conditions of long-term organotypic cultivation, Adv. Aging Res., 2012, vol. 1, pp. 31–37.
Grønskov, K., Ek, J., and Brondum-Nielsen, K., Oculocutaneous albinism, Orphanet. J. Rare Dis., 2007, vol. 2, pp. 43–53.
Gu, X., Neric, N.J., Crabb, J.S., Crabb, J.W., et al., Age-related changes in the retinal pigment epithelium (RPE), PLoS One, 2012, vol. 7, no. 6, e38673.
Gutteridge, J.M.C., Fluorescent products of phospholipid peroxidation: formation and inhibition in model systems, in Ceroid Lipofuscinosis (Batten’s Sisease), Armstrong, D., Koppang, N., and Rider, J.A., Eds., Amsterdam: Elsevier Biomedical Press, 1982, pp. 351–364.
Ilia, M. and Jeffery, G., Retinal mitosis is regulated by DOPA, a melanin precursor that may influence the time at which cells exit the cell cycle: analysis of patterns of cell production in pigmented and albino retinae, J. Comp. Neurol., 1999, vol. 405, pp. 394–405.
Iwai, L., Ramos, A., Schaler, A., et al., Retinal pigment epithelial integrity is compromised in the developing albino mouse retina, J. Comp. Neurol., 2016, vol. 524, no. 18, pp. 3696–3716.
Katz, M.L., Incomplete proteolysis may contribute to lipofuscin accumulation in the retinal pigment epithelium, Adv. Exp. Med. Biol., 1989, vol. 266, pp. 109–116.
Kennedy, C.J., Rakoczy, P.E., and Constable, I.J., Lipofuscin of the retinal pigment epithelium: a review, Eye (London), 1995, vol. 9, pp. 763–771.
Mazzoni, F., Safa, H., and Finnemann, S.C., Understanding photoreceptor outer segment phagocytosis: use and utility of RPE cells in culture, Exp. Eye Res., 2014, vol. 126, pp. 51–60.
Nadal-Nicolás, F.M., Salinas-Navarro, M., Jiménez-López, M., et al., Displaced retinal ganglion cells in albino and pigmented rats, Front. Neuroanat., 2014, vol. 8, pp. 99–105.
Nag, T.C., Maurya, M., and Sankar, RoyT., Age-related changes of the human retinal vessels: possible involvement of lipid peroxidation, Ann. Anat., Anatomischer Anzeiger, 2019, vol. 226, pp. 35–47.
Novikova, Yu.P., Gancharova, O.S., Eichler, O.V., et al., Preventive and therapeutic effects of SkQ1 containing Visomitin eye drops against light induced retinal degeneration, Biochemistry (Moscow), 2014, vol. 79, pp. 1101–1110.
Rozanowska, M., Properties and functions of ocular melanins and melanosomes, in Melanins and Melanogenesis: Biosynthesis, Biogenesis, Physiological and Pathological Functions, Borovansky, J. and Riley P.A., Eds., Weinheim, Germany: Wiley-Blackwell, 2011, pp. 187–224.
Sarna, T., Properties and function of ocular melanin-a photophysical view, J. Photochem. Photobiol. Biol. B., 1992, vol. 12, pp. 215–258.
Smith, R.T., Farsiu, S., and Allingham, M., Understanding RPE lipofuscin, Invest. Ophthalmol. Vis. Sci., 2016, vol. 57, no. 15, pp. 6766–6772.
Sparrow, J.R., Hicks, D., and Hamel, C.P., The retinal pigment epithelium in health and disease, Curr. Mol. Med., 2010, vol. 10, no. 9, pp. 802–823.
Strauss, O., The retinal pigment epithelium in visual function, Physiol. Rev., 2005, vol. 85, no. 3, pp. 845–881.
Summers, C.G., Albinism: classification, clinical characteristics, and recent findings, Optom. Vis. Sci., 2009, vol. 86, pp. 659–662.
Sundelin, S.P., Nilsson, S.E.G., and Brunk, U.T., Lipofuscin formation in cultured retinal pigment epithelial cells is related to the melanin content, Free Radical Biol. Med., 2001, vol. 30, no. 1, pp. 74–81.
Wang, N., Koutz C.A., Anderson R.E., A method for the isolation of retinal pigment epithelial cells from adult rats, Invest. Ophthalmol. Vis. Sci., 1993, vol. 34, no. 1, pp. 101–107.
Young, R.W., The renewal of rod and cone outer segments in the rhesus monkey, J. Cell Biol., 1971, vol. 49, pp. 303–309.
ACKNOWLEDGMENTS
We thank O.V. Kilina (MSU) for her assistance at the initial stage of conducting experiments. The work was carried out using the equipment of the Core Centrum of Koltzov Institute of Developmental Biology, Russian Academy of Sciences.
Funding
The work was performed within the framework of State Assignment of Koltzov Institute of Developmental Biology of Russian Academy of Sciences, 2020, no. 0108-2020-0005).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. During this study, all manipulations performed with experimental animals, methods of anesthesia, euthanasia, and animal care before and after experimental interventions matched international standards on bioethics.
AUTHOR CONTRIBUTIONS
Yu.P. Novikova participated in experiments and data processing, E.N. Grigoryan participated in conducting experiments, analyzing results, and writing articles.
Additional information
Translated by A. Ermakov
Rights and permissions
About this article

Cite this article
Novikova, Y.P., Grigoryan, E.N. Early Appearance of Aging Signs in the Retinal Pigment Epithelium in Young Albino Rats. Russ J Dev Biol 51, 377–386 (2020). https://doi.org/10.1134/S1062360420060065
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1062360420060065