
Abstract
In the intricate molecular world, post-translational modifications play pivotal roles in modulating protein activity and function. The Drosophila melanogaster genome encodes an array of enzymes to mediate these processes. Here, we elucidate a previously unrecognized connection between the Drosophila transforming growth factor-β-activated kinase 1 (dTak1) and the deubiquitinase Ovarian tumor (Otu). We demonstrate that dTak1 mediates both the phase-separated condensation and the deubiquitinase enzymatical activity of Otu in a manner independent of its kinase function. Using a combination of biochemical assays and in vivo genetic interactions, our findings highlight the significance of dTak1 in orchestrating the assembly of Otu condensates, and consequently, in the regulation of Otu’s biological functions. Our studies underscore the multifaceted nature of protein-protein interactions and open up avenues for exploring non-traditional roles of kinases in cellular processes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The cellular milieu, once thought of as a well-mixed solution of biomolecules, has in recent years been re-envisioned as a dynamic tapestry of micro-environments and structures (Ashfaq et al., 2020; Chen et al., 2023; Esvelt et al., 2011). One of the groundbreaking insights reshaping this understanding is the phenomenon of phase separation—the propensity of certain biomolecules, particularly proteins and nucleic acids, to demix from the surrounding medium, forming distinct, membrane-less compartments within the cell (Hyman et al., 2011; Aguzzi and Altmeyer, 2016). These liquid-liquid phase-separated domains, termed biomolecular condensates, are believed to serve as specialized reaction hubs, sequestering specific sets of molecules to regulate biological processes (Hyman et al., 2011; Chong et al., 2016).
While the principles governing phase separation are progressively being decoded, what remains intriguing is the regulatory mechanisms underpinning the formation, maintenance, and dissolution of these biomolecular condensates. Among the myriad of post-translational modifications that proteins undergo, phosphorylation stands out as a ubiquitous and versatile regulator of protein function, localization, and interaction (Derouiche and Mijakovic, 2012; Pawson and Scott, 2005). Given its dynamic and reversible nature, phosphorylation has the potential to be a key modulator of phase separation. For instance, phosphorylation can add a negatively charged phosphate group to a protein. This alteration normally leads to changes in protein-protein interactions, promoting or inhibiting phase separation depending on the context. Moreover, phosphorylation may induce conformational changes in proteins, which can expose or hide specific domains responsible for mediating phase separation. In addition, by modifying specific residues, phosphorylation enables to change the affinity between proteins, affecting multivalent interactions that drive phase separation. Over the last decades, a series of proteins (e.g., FUS, Tau, and MYC) have been demonstrated to undergo phase separation under the dynamical control of phosphorylation (Lao et al., 2022; Owen and Shewmaker, 2019; Li et al., 2022; Monahan et al., 2017; Boyko and Surewicz, 2023; Das et al., 2023).
Ovarian tumour (Otu) is a typical deubiquitinase (Dub) enzyme known for its critical roles in the ubiquitination/deubiquitination system in Drosophila melanogaster (Ji et al., 2017). A recent study revealed that Otu is essentially involved in maintaining intestinal homeostasis and delaying aging in adult flies (Ji et al., 2019). To fulfil these biological functions, Otu needs to shape into phase-separated condensates, thereby benefiting its Dub enzymatical activity (Ji et al., 2019). Yet, the dynamical regulation of such pivotal condensates remains to be deeply probed. Our prior attention was of course focused on kinases, regarding the broad involvements of kinase-mediated phosphorylation in modulating protein phase separation and condensation. Drosophila transforming growth factor-β-activated kinase 1 (dTak1) is a well-known kinase traditionally recognized for its roles in signal transduction pathways, particularly those implicated in inflammation and stress responses (Takatsu et al., 2000; Vidal et al., 2001; Sliverman et al., 2003; Kleino et al., 2005). In the canonical understanding, dTak1 functions primarily by phosphorylating specific substrates. This presents a compelling premise: could kinase dTak1 engage in mediating the condensate assembly of Otu?
In this discourse, we explore this very paradigm, delving into the interrelationship between the Dub Otu and the kinase dTak1. We show that dTak1 promotes both the condensation and the Dub enzymatical activity of Otu. Silencing dTak1 restricts Otu’s functions in controlling gut integrity and fly lifespan. Mechanistically, dTak1 positively contributes to Otu condensation largely in a phosphorylation-independent manner. Taken together, our studies advance our comprehension of the regulatory manner of Otu condensation.
MATERIALS AND METHODS
Drosophila Strains
All flies were maintained on the standard Drosophila medium (6.65% cornmeal, 7.15% dextrose, 5% yeast, 0.66% agar, 2.2% nipagin, and 3.4 mL/L propionic acid) at 25°C with a light/dark cycle of 12 h/12 h and 65% humidity. For genetic experiments employing the UAS/Gal4 system, crossings were first carried out at 18°C. After the eclosion of the progenies, flies were collected and maintained at 29°C for 5 to 7 d. The dTak1 RNAi strain was obtained from the Tsinghua RNAi Center (#0756). The UASp-Flag-Otu transgenic fly and the NP1-gal4; Tub-gal80 ts strain was described previously (Ji et al., 2019; Zhu et al., 2021).
Antibodies
The following primary antibodies were utilized for Western blot: Mouse anti-Otu (Ji et al., 2017); Rabbit anti-dTak1 (Abcam, Cat#239353); Mouse anti-β-Tubulin (Cwbio, Cat#CW0098M); Rabbit anti-Flag (Medical and Biological Laboratories, Cat#PM020); and Rabbit anti-Myc (Medical and Biological Laboratories, Cat#562). The secondary antibodies include Goat anti-Rabbit IgG H&L (Abcam, Cat#ab150078) and Goat anti-Mouse IgG H&L (Abcam, Cat#ab6789).
Semi-Denaturating Detergent Agarose Gel Electrophoresis (SDD-AGE)
The SDD-AGE assays were performed to determine the condensation status of Otu as previously described (Hua et al., 2022). In detail, dissected gut or S2 cell samples were lysed in buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 10% glycerol, 0.5% Triton X‑100). Samples were then centrifugated at 13 000 rpm for 10 min at 4°C. Supernatant was transferred to a fresh tube and incubated with equal volume of loading buffer (0.5× TBE, 10% glycerol, 2% SDS, 0.0025% bromophenol blue) at room temperature. The agarose (1.5%) gel was first run without samples for 15 min, followed by sample loading and electrophoresis for 1 h on ice. Protein condensates were transferred to a PVDF membrane. The membrane was first blocked in PBST (0.1% Tween-20 in PBS) buffer with 5% bovine serum albumin for 30 min at room temperature, and then incubated with indicated primary and secondary antibodies serially. The blot was revealed by using the enhanced chemiluminescence substrate (Bio-Rad, Cat#1705061) in a Bio-Rad platform.
S2 Cell Transfection and Western Blot Assay
S2 cells were manipulated according to the methods described previously (Cai et al., 2022). Briefly, cells were transfected with indicated expressing plasmids for 48 h using Lipofectamine 2000 reagents (Invitrogen, Cat#11668019). Cells were then harvested and lysed in buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 10% glycerol, 0.5% Triton X-100). Samples were subjected to centrifugation at 13 000 rpm for 10 min at 4°C, followed by phosphorylase treatment at 37°C for 30 min. Samples were then loaded on an 8 to 15% gradient SDS-PAGE gel and separated by electrophoresis, followed by transferring to a PVDF membrane. The antibody treatment and Western blotting were performed as described above.
Protein Purification in Eukaryotic System
HEK293 cells were cultured in DMEM Medium (Hyclone, Cat#SH30285.FS) with 10% fetal bovine serum (Gibco, Cat#10091155) and seeded in 10 cm culture plates at 105 cells per well. Cells were then transfected with 5 μg pcDNA3.0-Flag-Otu or pcDN-A3.0-Flag-dTak1 plasmids using Lipofectamine 2000 reagents (Invitrogen, Cat#11668019). 48 h after transfection, cells were lysed in buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5% Triton X-100, 10% glycerol with 1 mM phenylmethylsulfonyl fluoride). Lysates were then subjected to centrifugation at 13 000 rpm for 15 min at 4°C. The supernatants were transferred into fresh EP tubes and immunoprecipitated with Flag beads for 4 h, followed by washing treatment for 1 h. Beads were then incubated with Flag peptides (in 50 mM Tris-HCl, pH 7.5, 150 mM NaCl) to elute Flag-Otu or Flag-dTak1 proteins from Flag beads. To check the purification efficiency, 100 ng of indicated proteins were subjected to Western blot assays.
In Vitro Deubiquitination Assay
Indicated amounts of proteins or BSA were incubated with Ub-Rhodamine 110 (final concentration at 1 μM) in the reaction buffer (20 mM Tris-HCl, pH 7.5, 200 mM NaCl, 5 mM MgCl2, 2.5 mM dithiothreitol). Ub-Rhodamine 110 is an exquisitely sensitive Dub enzyme substrate for detecting ubiquitin C-terminal hydrolytic activity, because cleavage of the amide bond between the C-terminal glycine of ubiquitin and rhodamine by Dub leads to an increase in rhodamine fluorescence. The reaction mixture was added into a black 384-well plate and incubated at room temperature by using MD SpectraMax M5 Microplate Reader. Dynamic fluorescence was monitored with excitation and emission wavelengths set at 485/20 and 535/20 nm, respectively. Fluorescence intensity for each condition was averaged from triplicates and plotted as a function of time.
“Smurf” and Lifespan Assays
In order to avoid sex dimorphism, female flies were utilized for “Smurf” and lifespan assays as previously described (Zhu et al., 2021). In brief, fresh progenies (d 1 to 2) were collected and raised on normal medium for 2 d (for mating). Females were then collected and transferred to new vials containing normal medium. The number of flies in each vial was between 30 to 35. For lifespan analyses, flies were counted for death every 2-d throughout adult life. For “Smurf” assays, flies were transferred to medium containing blue dye (2.5%, FD&C blue #1) at indicated time points (d 10, 30, and 50, respectively). 6 h later, the numbers of “Smurf” flies were counted.
Statistical Analyses
All statistical analyses were performed by using the GraphPad Prism 9. Statistical significances in Figs. 1b, 1c, 2c, and 3a were determined by using the ANOVA test. Statistical significances in Fig. 3b were calculated by using the Log-Rank test. The p value of less than 0.05 was considered statistically significant. *, p < 0.05; **, p < 0.01; ns, not significant.

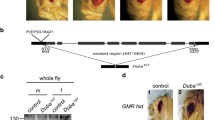
dTak1 mediates Otu condensation. (a) Guts were dissected from young (5- to 7-day) flies including NP1>+ (wild-type control) and NP1>dTak1 RNAi. Samples were lysed in lysis buffer and subjected to SDD-AGE to examine the condensate assembly of Otu. SDS-PAGE assays were performed to determine the expression levels of indicated proteins. (b and c). Densitometry analyses to quantify condensate (b) or protein (c) levels of Otu are shown. Each dot represents one biological replicate. Data were collected from 3 independent replicates and shown as means plus standard errors. **, p < 0.01; ns, not significant. (d) 100 ng of purified Flag-Otu or Flag-dTak1 were subjected to Western blot assay. (e) In vitro Dub assay showing that addition of dTak1 enhances the Dub enzymatical activity of Otu in a dosage-dependent manner.

dTak1KD is dispensable for modulating Otu condensation and Dub enzymatical activity. (a) Guts were dissected from flies including NP1>+ (wild-type control) and NP1>dTak1 RNAi. Samples were lysed and treated with phosphatase (Lanes 3 and 4), followed by Western blot assays. (b) S2 cells were transfected with indicated combinations of expressing plasmids, followed by SDD-AGE and SDS-PAGE assays. (c) Densitometry analyses to quantify Otu condensate are shown. Each dot represents one biological replicate. Data were collected from 3 independent replicates and shown as means plus standard errors. *, p < 0.05; ns, not significant. (d and e) In vitro Dub assays were performed to examine the effects of purified dTak1WT or dTak1KD on the Dub enzymatical activity of Otu.

Silencing dTak1 prevents Otu’s roles in regulating gut barrier function and fly lifespan. (a and b) Percentages of “Smurf” (a) and lifespan curves (b) of flies including NP1>+, NP1>Flag-Otu, NP1>dTak1 RNAi, and NP1>dTak1 RNAi;Flag-Otu. In (a), each dot represents one biological replicate. Data were collected from 3 independent replicates and shown as means plus standard errors. The numbers of flies for “Smurf” assays are as follows. On day 10, NP1>+: 72, 67, 71; NP1>Flag-Otu: 68, 69, 66; NP1>dTak1 RNAi: 69, 71, 65; NP1>dTak1 RNAi;Flag-Otu: 70, 66, 71. On day 30, NP1>+: 66, 64, 71; NP1>Flag-Otu: 70, 71, 67; NP1>dTak1 RNAi: 65, 69, 73; NP1>dTak1 RNAi; Flag-Otu: 66, 68, 70. On day 50, NP1>+: 65, 68, 65; NP1>Flag-Otu: 67, 69, 70; NP1>dTak1 RNAi: 62, 68, 65; NP1>dTak1 RNAi; Flag-Otu: 67, 64, 69. *, p < 0.05; **, p < 0.01; ns, not significant. In (b), the numbers of flies in lifespan assays are: NP1>+: 104; NP1>Flag-Otu: 102; NP1>dTak1 RNAi: 105; NP1>dTak1 RNAi;Flag-Otu: 104.
RESULTS
dTak1 is Essential for Otu Condensation
To investigate whether dTak1 is involved in mediating the phase-separated condensation of Otu, we utilized the UAS/Gal4 system and generated flies with dTak1 RNAi in guts. We dissected guts from age-paired NP1>dTak1 RNAi and the NP1>+ (wild-type control) flies and examined the Otu condensates by performing the SDD-AGE assays as previously described (Hua et al., 2022). Consistent with what was found previously (Ji et al., 2019), endogenous Otu underwent condensation in intestinal cells (Figs. 1a and 1b). Intriguingly, when dTak1 was knocked down, the level of Otu condensates was markedly reduced (Figs. 1a and 1b), indicating that the kinase dTak1 plays a positive role in governing the condensate assembly of Otu. Notably, silencing dTak1 did not affect the protein level of Otu (Figs. 1a and 1c).
dTak1 Mediates the Dub Enzymatical Activity of Otu
Condensation represents an active state of Otu efficiently executing its Dub enzymatical activity (Ji et al., 2019). We therefore examined whether dTak1 contributes to the Dub enzymatical activity of Otu. For this, we purified Flag-Otu and Flag-dTak1 proteins from a eukaryotic system (Fig. 1d). As demonstrated in the in vitro Dub assays, additional dTak1 elevates the enzymatical activity of Otu in cleaving ubiquitin from substrates in a dosage-dependent manner (Fig. 1e).
dTak1 Is Dispensable for Affecting Otu Phosphorylation
dTak1 has been demonstrated to play crucial roles in a series of biological processes, including immunity, apoptosis, and organismal development in Drosophila (Takatsu et al., 2000; Vidal et al., 2001; Sliverman et al., 2003; Kleino et al., 2005). As a kinase, dTak1 normally phosphorylates the targeted substrates to change the activity, stability, and/or function of the phosphorylated proteins (Takatsu et al., 2000; Shin et al., 2013; Chen et al., 2017). We therefore examined whether dTak1 is involved in modulating Otu phosphorylation. For this, we dissected guts from adult flies and prepared lysates for phosphatase treatment. As demonstrated in Fig. 2a, Otu was detected with a smaller size in the samples with Alkaline treatment, indicating that Otu is phosphorylated in gut cells. However, silencing dTak1 didn’t affect the phosphorylation status of Otu (Fig. 2a).
To explore whether the kinase activity of dTak1 is responsible for its contribution to Otu condensation, we utilized the kinase dead form of dTak1 (dTak1KD) for SDD-AGE assays. As illustrated in Figs. 2b and 2c, supplementation of dTak1KD still promoted Otu condensation. To obtain more evidence, we carried out the in vitro Dub assays to test the potential role of dTak1KD in regulating the enzymatical activity of Otu. In this assay, the same amount of the wild-type dTak1 (dTak1WT) was used as the positive control. As expected, additional dTak1KD indeed enhanced the Dub enzymatical activity of Otu (Figs. 3d and 3e). Put together, our results support the notion that dTak1 positively regulates Otu condensation and Dub enzymatical activity in a kinase-independent manner.
Silencing dTak1 Prevents Otu’s Roles in Modulating Gut Homeostasis and Fly Lifespan
Phase-separated condensation is essential for Otu executing its functional assessments in controlling intestinal homeostasis and Drosophila lifespan (Ji et al., 2019). Since dTak1 plays a critical role in Otu condensation, one would like to expect that dTak1 contributes to fulfilling Otu’s biological functions. To prove this, we first carried out the “Smurf” assay as we previously did (Zhu et al., 2021). We observed that over-expression of Otu in intestinal cells reduced the percentages of “Smurf” (Fig. 3a). However, knocking down dTak1 markedly prevented the beneficial role of Otu in maintaining gut barrier function (Fig. 3a). Consistently, we obtained similar results when we performed lifespan assays (Fig. 3b).
DISCUSSION
The findings of this study provide compelling evidence that in Drosophila, dTak1 mediates the condensation and the Dub enzymatical activity of Otu in a manner that is independent of its kinase activity. This challenges the conventional understanding that the primary role of dTak1 in cellular signaling is primarily as a kinase.
Phase separation, often observed as the formation of biomolecular condensates, is emerging as a crucial organizing principle in cellular biology (Hyman et al., 2011; Aguzzi and Altmeyer, 2016; Chong et al., 2016). These condensates can serve various cellular functions, including signal transduction, RNA processing, and DNA repair (Boeynaems et al., 2018; Mitrea and Kriwacki, 2016). The data presented here suggest that Otu undergoes phase-separated condensation, which is mediated by dTak1. This is intriguing given that many proteins that undergo phase separation are typically enriched in low complexity or intrinsically disordered regions, which was also reported for Otu (Ji et al., 2019). Further studies might delve into the specific domains or motifs in Otu responsible for this phenomenon. It’s remarkable that the mediation of Otu condensation by dTak1 is independent of its kinase activity. As a kinase, dTak1 is typically associated with the phosphorylation of target proteins, leading to the activation or inactivation of various signaling pathways (Takatsu et al., 2000; Vidal et al., 2001; Sliverman et al., 2003; Kleino et al., 2005). The non-canonical role of dTak1, as elucidated in this paper, opens avenues for understanding the multifunctionality of kinases beyond their enzymatic activity. Such kinase-independent roles might be widespread and offer alternative regulatory nodes that have been overlooked in the analyses of traditional signaling pathways. One strategy in the following studies is to utilize different truncated forms of dTak1 proteins and to explore which region(s) of dTak1 is (are) responsible for its regulation on Otu condensation. Another challenge moving forward will be deciphering the structural basis for dTak1’s interaction with Otu. Does dTak1 bind directly to Otu? Or are there intermediate proteins or other molecules involved in this process? Identifying these interaction sites might provide new insights into understanding how Otu forms functional condensates.
The Dub enzymatical activity of Otu has been well established in previous literature (Ji et al., 2017, 2019). Ubiquitination is a post-translational modification that plays a pivotal role in various cellular processes, from protein degradation to DNA repair and signal transduction (Pickart and Eddins, 2004). The finding that dTak1 can modulate the Dub activity of Otu offers a novel regulatory mechanism that might be crucial for the fine-tuning of these processes. This dTak1-mediated regulation of Otu’s Dub activity can provide the cell with an efficient mechanism to respond to various internal or external stimuli promptly. In this study, we indeed observed that silencing dTak1 prevented Otu’s functions in maintaining gut integrity and delaying aging. Our current data of course cannot exclude other biological roles of dTak1-mediated Otu condensates. It would be worthwhile in the future to test this regulatory relationship in other biological processes in Drosophila and potentially, other organisms.
In conclusion, the findings presented in this paper shed light on a previously unrecognized aspect of dTak1 function in Drosophila. By elucidating a kinase-independent role of dTak1 in the regulation of Otu condensation, this work broadens our understanding of the intricacies of cellular regulation and offers exciting avenues for future research.
REFERENCES
Aguzzi, A., and Altmeyer, M., Trends Cell Biol., 2016, vol. 26, no. 7, pp. 547–558.
Ashfaq, A., Clochard, M.C., Coqueret, X., Dispenza, C., Driscoll, M.S., Ulanski, P., and Al-Sheikhly, M., Polymers (Basel), 2020, vol. 12, no. 12, p. 2877.
Boeynaems, S., Alberti, S., Fawzi, N.L., Mittag, T., Polymenidou, M., Rousseau, F., Schymkowitz, J., Shorter, J., Wolozin, B., Bosch, L.V.D., Tompa, P., and Fuxreiter, M., Trends Cell Biol., 2018, vol. 28, no. 6, pp. 420–435.
Boyko, S. and Surewicz, W.K., J. Biol. Chem., 2023, vol. 299, no. 6, p. 104722.
Cai, Q., Guo, H., Fang, R., Hua, Y., Zhu, Y., Zheng, X., Yan, J., Wang, J., Hu, Y., Zhang, C., Zhang, C., Duan, R., Kong, F., Zhang, S., Chen, D., and Ji, S., Cell Rep., 2022, vol. 41, no. 11, p. 111795.
Chen, Y., Zuo, M., Chen, Y., Yu, P., Chen, X., Zhang, X., Yuan, W., Wu, Y., Zhu, W., and Zhao, Y., Nat. Commun., 2023, vol. 14, no. 1, p. 5229.
Chen, L., Paquette, N., Mamoor, S., Rus, F., Nandy, A., Leszyk, J., Shaffer, S.A., and Silverman, N., J. Biol. Chem., 2017, vol. 292, no. 21, pp. 8738–8749.
Chong, P.A. and Forman-Kay, J.D., Curr. Opin. Struct. Bio-l., 2016, vol. 41, pp. 180–186.
Das, S.K., Lewis, B.A., and Levens, D., Trends Cell Biol., 2023, vol. 33, no. 3, pp. 235–246.
Derouiche, A., Cousin, C., and Mijakovic, I., Curr. Opin. Biotechnol., 2012, vol. 23, no. 4, pp. 585–590.
Esvelt, K.M., Carlson, J.C., and Liu, D.R., Nature, 2011, vol. 472, no. 7344, pp. 499–503.
Hua, Y., Zhu, Y., Hu, Y., Kong, F., Duan, R., Zhang, C., Zhang, C., Zhang, S., Jin, Y., Ye, Y., Cai, Q., and Ji, S., Front. Immunol., 2022, vol. 13, p. 932268.
Hyman, A.A., Weber, C.A., and Julicher, F., Annu. Rev. Cell Dev. Biol., 2011, vol. 30, pp. 39–58.
Ji, S., Li, C., Hu, L., Liu, K., Mei, J., Luo, Y., Tao, Y., Xia, Z., Sun, Q., and Chen, D., Proc. Natl. Acad. Sci. U. S. A., 2017, vol. 114, no. 24, pp. 6316–6321.
Ji, S., Luo, Y., Cai, Q., Cao, Z., Zhao, Y., Mei, J., Li, C., Xia, P., Xie, Z., Xia, Z., Zhang, J., Sun, Q., and Chen, D., Mol. Cell, 2019, vol. 74, no. 2, pp. 363–377.
Kleino, A., Valanne, S., Ulvila, J., Kallio, J., Myllymaki, H., Enwald, H., Stoven, S., Poidevin, M., Ueda, R., Hultmark, D., Lemaitre, B., and Ramet, M., EMBO J., 2005, vol. 24, no. 19, pp. 3423–3434.
Lao, Z., Dong, X., Liu, X., Li, F., Chen, Y., Tang, Y., and Wei, G., J. Chem. Inf. Model., 2022, vol. 62, no. 13, pp. 3227–3238.
Li, J., Zhang, M., Ma, W., Yang, B., Lu, H., Zhou, F., and Zhang, L., Mol. Biomed., 2022, vol. 3, no. 1, p. 13.
Mitrea, D.M. and Kriwacki, R.W., Cell Commun. Signal., 2016, vol. 14, p. 1.
Monahan, Z., Ryan, V.H., Janke, A.M., Burke, K.A., Rhoads, S.N., Zerze, G.H., O’Meally, R., Dignon, G.L., Conicella, A.E., Zheng, W., Best, R.B., Cole, R.N., Mittal, J., Shewmaker, F., and Fawzi, N.L., EMBO J., 2017, vol. 36, no. 20, pp. 2951–2967.
Owen, I. and Shewmaker, F., Int. J. Mol. Sci., 2019, vol. 20, no. 21, p. 5501.
Pawson, T. and Scott, J.D., Trends Biochem. Sci., 2005, vol. 30, no. 6, pp. 286–290.
Pickart, C.M. and Eddins, M.J., Biochim. Biophys. Acta, 2004, vol. 1695, nos. 1–3, pp. 55–72.
Shin, J.H., Min, S.H., Kim, S.J., Kim, Y.I., Park, J., Lee, H.K., and Yoo, O.J., Sci. Rep., 2013, vol. 3, p. 1561.
Silverman, N., Zhou, R., Erlich, R.L., Hunter, M., Bernstein, E., Schneider, D., and Maniatis, T., J. Biol. Chem., 2003, vol. 278, no. 49, pp. 48928–48934.
Takatsu, Y., Nakamura, M., Stapleton, M., Danos, M.C., Matsumoto, K., O’Connor, M.B., Shibuya, H., and Ueno, N., Mol. Cell Biol., 2000, vol. 20, no. 9, pp. 3015–3026.
Vidal, S., Khush, R.S., Leulier, F., Tzou, P., Nakamura, M., and Lemaitre, B., Genes Dev., 2001, vol. 15, no. 15, pp. 1900–1912.
Zhu, Y., Cai, Q., Zheng, X., Liu, L., Hua, Y., Du, B., Zhao, G., Yu, J., Zhuo, Z., Xie, Z., and Ji, S., Aging Dis., 2021, vol. 12, no. 7, pp. 1821–1834.
ACKNOWLEDGMENTS
We thank the Tsinghua Stock Centre for providing fly strains.
Funding
This work was supported by the grant 2008085J14 from the Anhui Provincial Natural Science Foundation, China.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
CONFLICT OF INTEREST
The authors of this work declare that they have no conflicts of interest.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
This work does not contain any studies involving human and animal subjects.
Additional information
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chuchu Zhang, Shao, X., Xia, Z. et al. Drosophila Tak1 Mediates the Condensation and Deubiquitinase Activity of Otu in a Kinase-Independent Manner. Biol Bull Russ Acad Sci 51, 511–517 (2024). https://doi.org/10.1134/S1062359023605013
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1062359023605013