
Abstract
A population-based approach to determining the borders and number of natural plague foci implies that the specialists in the field accept the concept that the existence of a natural plague focus is always provided by a single population of the main carrier. The discussion of this approach was motivated by the data collected in the course of a long-term study of the Hissar natural plague focus in Tajikistan. It was demonstrated that the area of this focus constitutes only 300 km2, and its functioning is maintained by a single relatively small population of Microtus carruthersi Thomas, 1909, the main plague carrier. The Hissar mountain range territories neighboring the focus have been surveyed for plague for more than 20 years. However, the plague pathogen (Yersinia pestis Lehmann, Neuman, 1986) could not be detected notwithstanding the absolute identity of the species composition of rodents and fleas. Many known natural plague foci are considerably larger in size than the Hissar focus and are inhabited by several populations of the main carrier. For example, five groups of populations of the common vole (Microtus arvalis Pallas, 1778), the main plague carrier, are described in the Eastern Caucasus high mountain focus occupying an area of 23 500 km2. In the Tuva mountain focus, the plague pathogen was found in six populations of the main carrier, the long-tailed ground squirrel (Spermophilus undulatus Pallas, 1778). On the territory of the Muyunkum desert natural plague focus occupying an area of 93 000 km2, 17 populations of the great gerbil (Rhombomys opimus Lichtenstein, 1823), the main plague carrier, are distinguished. Similar data are available for a number of other foci. Significantly, in different populations of the main carriers, the differences in the genotypes of both rodents and the plague pathogen were described, and asynchrony in plague epizootics could be traced. In such a way, the plague parasitic cenosis functioning within the borders and the habitation areal of a single population of the main carrier constitutes an autonomous standalone natural focus. Hence, the foci occupied by several populations of the main carrier are, in fact, groups of autonomous foci with a similar biocenotic structure. Therefore, a considerably larger number of natural (autonomous) foci are present within the plague natural focality zones than is commonly assumed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
In 1970, on the northern macroslope of the Hissar mountain range (Tajikistan) in the area of Iskanderkul’ Lake, a natural plague focus was detected with Carruther’s vole as the main carrier (Microtus carruthersi Thomas, 1909). The area of the focus proved to be relatively small (300 km2), with only one population of Carruther’s vole inhabiting its territory. These data suggested that at least some of the known natural plague foci, the large ones (thousands of square kilometers) inhabited within their borders by several populations of primary carriers, are in fact groups of autonomous foci with a close biocenotic structure.
The theoretical basis for the study of parasitic systems at the population level was provided by V.N. Beklemishev (1956), who believed that the concept of simple or complex parasitic system, as well as the concept of a parasitic system in general, refers not to the species as a whole, but always to the population. Studies of the spatial structure were carried out for a number of natural plague foci, and the populations of the main plague carriers inhabiting them were described. According to D.I. Bibikov (1965), autonomous plague foci in the syrt part of the Central Tien Shan are associated with groups of geographical populations of the European marmot (Marmota baibacina Kastschenko, 1899). According to A.I. Dyatlov et al. (1980) and G.V. Trufanov and P.D. Golubev (1982), several populations of the main plague carrier, the mountain ground squirrel (Spermophilus musicus Menetrie, 1832), inhabit the Elbrus region (Central Caucasus highland natural plague focus). In the Tuva mountain focus, the plague pathogen (Yersinia pestis Lehmann, Neuman, 1986) was detected in six populations of the main carrier, the long-tailed ground squirrel (Spermophilus undulatus Pallas, 1778). The complete independence of these rodent populations was noted (Verzhutskii, 2005). Four populations of the main carrier, the Mongolian pika (Ochotona pricei Thomas, 1911), were identified on the territory of the Gornyi Altai highland plague foci in southeastern Altai. A high level of geographical isolation is observed amoung all the spatial groups (Korzun et al., 2016).
Having identified the populations of the main carriers, the specialists shifted away from the population approach in their further interpretation of the results obtained: the plague enzootic territory occupied by a single population of the main carrier is given a rather undefined status, a focality region, or mezofocus. The term mezofocus, proposed by B.K. Fenyuk (1958), is deciphered as a relatively small area of an autonomous focality region, strongly, sometimes very strongly, spatially restricted, but characterized by clear biotopic and biocenotic distinguishing features compared to the surrounding territories; mezofocus territories are the favorable habitats of the carriers or vectors, which are characterized by the constant inflow of the latter. As the definition makes clear, this is not a question of compatibility between the mezofocus area and the main carrier population range.
The authors cited above clearly indicate the high degree of geographic isolation of the carrier populations, which is typical for mountain landscapes in general. Consequently, there is similar isolation between the populations of the plague pathogen and its vectors (different flea species). The possibilities for crossing the isolation barriers are, undoubtedly, the highest for the plague carriers. Fleas and the plague pathogen depend entirely on the mammalian transporters. Hence, the plague parasitic system functioning within the boundaries and on the habitation territory of a single population of the main carrier constitutes an independent natural focus. According to the definition proposed by V.V. Kucheruk (1972), the name of an individual natural focus should be attributed to the smallest area of the Earth’s surface within the borders of which under the existing conditions the pathogen circulates without being imported from the external environment for an indefinitely long period of time (dozens of successive ascending and descending cycles of epizootics).
Thus, if we take the population structure of the main plague carriers as the basis for identifying natural plague foci, then, undoubtedly, we need to revise the estimation of the borders and the number of plague foci described to date. Apparently, they are more numerous than is generally accepted.
MATERIALS AND METHODS
In the period 1976–1991, in order to study the spatial structure of the Carruther’s vole (Microtus carruthersi) population in the Hissar natural plague focus and on the adjacent territories, mapping of the settlements of this rodent was carried out. Three variants of settlement delineation were used due to the labor intensity of mapping in the mountains.
(1) Direct measurement of the length of the settlement boundaries and their contour (using a compass).
(2) The settlement was crossed approximately in the middle by a route, and perpendicular measurements (routes) were taken to the edge of the settlement at regular intervals. By connecting the points obtained on the plan, the settlement contour was obtained.
(3) The compass triangulation technique. Along the route through the settlement (the gorge bottom), landmarks along the settlement margin (on the range slope) were chosen. For each landmark, two or three spots along the route crossing the settlement azimuths were determined using a compass. The junction of the locations (determined by the crossing point of the azimuths drawn from one route point) of several landmarks shows the boundary of the settlement on the plan.
As a result of mapping, it was found that seven populations of Carruther’s vole inhabit the central part of the Hissar mountain range (the area of Iskanderkul’ Lake and Anzob Pass), with only one of them revealing circulation of the plague pathogen. It should be stressed that the territory of the Hissar mountain range beyond the plague focus borders has been surveyed for plague for over 20 years (both the northern and southern macroslopes). Nevertheless, neither in the northern macroslope areas neighboring the focus, nor on the southern macroslope, has the plague pathogen been detected despite the complete identity of the species composition of rodents and fleas.
RESULTS AND DISCUSSION
Mapping revealed that a single population of Carruther’s vole inhabits the Hissar natural focus and occupies 19 settlements with a total area of 3246 ha (Sludskii et al., 2003). Consequently, plague enzootia may be caused by a relatively small population of the main carrier. Most of the known natural plague foci are much larger in size than the Hissar focus, and each of them is inhabited by several populations of primary carriers. Another natural plague focus of the vole type (the main carrier is the common vole Microtus arvalis Pallas, 1778) is the East Caucasus highland focus with an area of 23 500 km2. In the highlands of the East Caucasus, five groups of common vole populations and three plague focality areas are distinguished (Dyatlov and Kazakov, 1987; Kazakov, 1989). The clear geographical isolation of the common vole populations suggests that there is no constant exchange of individuals between the populations and, consequently, that plague enzootics are independent in each population.
As has already been mentioned above, several populations of the mountain ground squirrel inhabit the territory of the Central Caucasus highland focus, which occupies an area of 4300 km2. It is indicative that in the Central Caucasus plague focus the distribution of biochemical phenes over the habitation range of mountain ground squirrels reveals a clear subdivision of populations into two groups, western and eastern, separated by the Malka River. The Neopsylla setosa Wagner, 1898 and Ctenophthalmus orientalis Wagner, 1898 fleas can be found only in the eastern part. In the western areas of the focus, specific strains of the plague pathogen (with proline auxotrophy and heterogenic virulence) circulate. Strains from the western part of the focus bear a 4 MDa cryptic plasmid, while the eastern strains lack it, which means that the differences between the strains are fixed at the genotype level. In the western parts of the Central Caucasus highland focus, the specific, somewhat reduced, plague parasitic system has developed with Citellophilus tesquorum Wagner, 1898 as the only main carrier and with the more conservative (proline-dependent) variant of the plague pathogen (Grigor’ev, 1998; Bryukhanov et al., 2001; Dyatlov et al., 2001). These facts clearly point to the independent existence and functioning of plague parasitic systems in the western and eastern parts of the Central Caucasus focus and, hence, to the presence of at least two natural plague foci in the central part of the Greater Caucasian Mountain Range with the mountain ground squirrel as the main carrier.
In the Gornyi Altai focus, three focality areas have been identified, namely, Ulandryk, Tarkhata, and Kurai, which are geographically and functionally associated with the corresponding Mongolian pika populations. The areas of these zones are 934, 911, and 350 km2, respectively. The focality areas (and populations) are isolated from each other by distances of 7–25 km. An important fact has been established that the plague pathogen circulating in each of these areas has specific phenotypic and genotypic characteristics (Balakhonov et al., 2009, 2014; Logachev et al., 2012; Popkov et al., 2012). Notwithstanding these data V.M. Korzun et al. (2016) still consider the Gornyi Altai natural plague focus as an integral parasitic system. At the same time, they note the high degree of geographical isolation between all the spatial groups (populations) of the Mongolian pika. Here, we can observe a contradiction between the actual data and the conclusions made.
We believe that based on the size, the high degree of isolation of each population, and the presence of genotypic differences in the plague microbe strains circulating in different populations, the so-called focality areas described above should be regarded as independent natural foci.
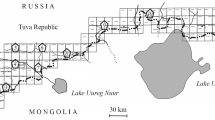
In Southwest Tuva, eight populations of the long-tailed ground squirrel are distinguished, between which there occur uninhabited areas from 1–2 to 10–12 km across. No visible exchange of individuals between the populations has been noted. The spatial structure of the Tuva natural plague focus is currently entirely determined by the pattern of territorial distribution of the main carrier, the long-tailed ground squirrel, and its dominant flea species. The presence of the plague pathogen was found in six populations of the long-tailed ground squirrel out of the eight described in southwestern Tuva. Given the complete independence of the populations of this animal, the territories with the presence of the pathogen within the borders of a separate ground squirrel population are referred to as independent areas of focality, i.e., mezofoci. It was found that, in most cases, the boundaries of the host population and its specific ectoparasite Citellophilus tesquorum mostly coincide. Differences in the nutritional requirements of the plague pathogen between the focality areas have been revealed. The dynamics of the epizootic activity of the individual focality areas indicates certain independence of epizootics which take place in them (Verzhutskii, 2003, 2005). The materials presented convincingly demonstrate the presence of several natural plague foci with the same main carrier, the long-tailed ground squirrel, in southwestern Tuva.
Despite the difficulties in determining the boundaries between the populations of plague carriers in the absence of spatial and territorial barriers, a number of scientists still describe the population structure of the main plague carriers in lowland plague foci. Yu.A. Dubrovskii and V.V. Kucheruk (1971) identify about 70 populations of the great gerbil (Rhombomys opimus Lichtenstein, 1823), the main plague carrier, within the Central Asia desert focus. L.P. Rapoport et al. (1977) for the territory of Muyunkum (Moyunkum desert natural plague center, covering an area of 93 000 km2) distinguished 17 populations of Great Gerbils. The populations differ in the dynamics of the population numbers and show the presence of settlement cores and less densely populated peripheral zones. Therefore, significant differences in the patterns of the plague epizootic process in different parts of the Muyunkum desert focus are observed. Simultaneously with the activation of the epizootic process in some areas, its decline may be observed in other areas. This is due to the fact that each great gerbil population has its own rhythm of fluctuations in the population number, which depends on both external and intrapopulation factors (Rapoport et al., 2002). Two populations of the great gerbil inhabit the left bank of the Ili River (left-bank part of the Ili intermountain focus with an area of 23 900 km2). The populations differ in the manifestations of the plague epizootic process, i.e., in the intensity and extensiveness of epizootics and the virulence of the isolated strains of the plague pathogen (Kasenova et al., 2004). It can be stated with a great deal of certainty that some of the lowland natural plague foci described so far (Cadastre of Epidemic …, 2016) are actually groups of “twin” foci. As E.I. Korenberg (2010) emphasizes, many anti-plague service specialists give the name of the natural focus to a vast territory with a similar main carrier.
CONCLUSIONS
Korenberg, based on the assessment of the structure of the habitation areal of a number of pathogens causing infectious diseases with natural foci, believes that the real number of natural foci is much larger than is generally assumed. For example, within the range of the tick-borne encephalitis virus, there seem to be several tens of thousands of natural foci (Korenberg et al., 2013). In the zone of the natural plague focality, there, undoubtedly, also exist a larger number of natural (autonomous) foci than is commonly believed.
The acceptance of this general pattern, namely, one population of the main carrier and one natural plague focus, entails the need to correct the approaches to epidemiological surveillance, to replace (or update) the set of basic instructional and methodological documents, etc. It may be for this reason that anti-plague system experts, the scientific developments of whom are “tightly” connected with their practical work, are cautious in interpreting the results of the studies of the population structure of the main carriers in connection with plague enzootics.
REFERENCES
Balakhonov, S.V., Korzun, V.M., Chipanin, E.B., Afanas’ev, M.V., Mikhailov, E.P., Denisov, A.V., Fomina, L.A., Eshelkin, I.I., Mashkovsky, I.K., Mishchenko, A.I., Rozhdestvensky, E.N., and Yarygina, M.B., Gorno-Altaiskii prirodnyi ochag chumy: retrospektivnyi analiz, epizootologicheskii monitoring, sovremennoe sostoianie (Gorno-Altai Natural Plague Focus: Retrospective Analysis, Epizootiological Monitoring, and Current State), Balakhonov, S.V. and Korzun, V.M., Eds., Novosibirsk: Nauka-Tsentr, 2014.
Balakhonov, S.V., Shestopalov, M.Yu., and Romanova, I.F., Results of the VNTR-analysis in the locus (5'-NAAA-3')n of Yersinia pestis strains from the active natural foci of plague of Siberia, Mol. Genet., Microbiol., Virol., 2009, no. 3, pp. 14–17.
Beklemishev, V.N., Agents of diseases as members of biocenoses, Zool. Zh., 1956, vol. 35, no. 12, pp. 1755–1779.
Bibikov, D.I., Marmots and plague in Central Asia mountains, Extended Abstract of Doctoral (Biol.) Dissertation, Alma-Ata, 1965.
Bryukhanov, A.F., Eremenko, E.D., Bryukhanova, G.D., Grizhebovsky, G.M., Zharinova, N.V., Tsareva, N.S., and Mezentseva, O.N., Genotyping of plague microbe strains by variable number tandem repeats, Karantin. Zoonozn. Infekts. Kazakh., 2001, no. 3, pp. 72–75.
Dubrovskii, Yu.A. and Kucheruk, V.V., Spatial structure of Central Asiatic–Kazakhstan part of great gerbil (Rhombomys opimus) range, Zool. Zh., 1971, vol. 50, no. 2, pp. 259–273.
Dyatlov, A.I. and Kazakov, V.P., Population structure of common vole settlements in High-Mountain Dagestan, in Osobo opasnye infektsii na Kavkaze: tezisy dokladov (Particularly Dangerous Infections in the Caucasus, Abstracts of Papers), Stavropol: Nauchno-issledovatel’skii protivochumnyi institut Kavkaza i Zakavkaz’ia Ministerstva zdravookhraneniia SSSR, 1987, pp. 297–299.
Dyatlov, A.I., Petrov, P.A., Golubev, P.D., and Trufanov, G.V., Concerning the structure of the small souslik (Citellus pygmaeus Pall., 1778) range in the vicinity of the Mount Elbrus, Ekologiya, 1980, no. 5, pp. 77–83.
Dyatlov, A.I., Antonenko, A.D., Grizhebovsky, G.M., and Labunets, N.F., Prirodnaya ochagovost' chumy na Kavkaze (Natural Plague Focality in the Caucasus), Stavropol: Protivochumnyi institut., 2001.
Fenyuk, B.K., Geography issues of natural plague foci, Zool. Zh., 1958, vol. 37, no. 7, pp. 961–971.
Grigor’ev, M.P., Analysis of the vector population structure in Central-Caucasian natural plague focus, Extended Abstract of Cand. Sci. (Biol.) Dissertation, Stavropol, 1998.
Kadastr epidemicheskikh i epizooticheskikh proiavlenii chumy na territorii Rossiiskoi Federatsii i stran blizhnego zarubezh’ya (s 1876 po 2016 god) (Cadastre of Epidemic and Epizootic Plague Manifestations on the Territory of the Russian Federation and Former Soviet Union States (between 1876 and 2016)), V.V. Kutyrev and A.Yu. Popova, Eds., Saratov: Amirit, 2016.
Kasenova, A.K., Sabilaev, A.S., Davydova, V.N., Sapozhnikov, V.I., Larionov, G.I., and Kosilov, V.A., Some peculiarities of plague epizooties in the left bank areas of the Il’ River, Karantin. Zoonozn. Infekts. Kazakh., 2004, no. 1, pp. 44–47.
Kazakov, V.P., Natural plague focality in high-mountain areas of East Caucasus, Extended Abstract of Cand. Sci. (Biol.) Dissertation, Saratov, 1989.
Korenberg, E.I., Natural focality of infections: modern issues and research prospects, Zool. Zh., 2010, vol. 89, no. 1, pp. 5–13.
Korenberg, E.I., Pomelova, V.G., and Osin, N.S., Prirodno-ochagovye infektsii, peredayuschiesya iksodovymi kleschami (Infections with Natural Focality Transmitted by Ixodid Ticks), Gintsburg, A.L. and Zlobin, V.N., Eds., Moscow: Nauchno-issledovatel’skii institut biologicheskogo priborostroeniia, 2013.
Korzun, V.M., Balakhonov, S.V., Chipanin, E.V., Denisov, A.V., Mikhailov, E.P., Mishchenko, A.I., Yarygina, M.B., Rozhdestvensky, E.N., and Fomina, L.A., Formation, evolution, and functioning of natural plague focus in Altai Mountains, Med. Parazitol. Parazitar. Bol., 2016, no. 1, pp. 17–25.
Kucheruk, V.V., Structure, typology, and zoning of natural foci of human diseases, in Itogi razvitiia ucheniia o prirodnoi ochagovosti boleznei cheloveka i dal’neishie zadachi (Results of Development of the Doctrine on the Natural Focality of Human Diseases and Further Tasks), Moscow: Meditsina, 1972, pp. 180–212.
Logachev, A.I., Korzun, V.M., Mikhailov, E.P., Rozhdestvensky, E.N., and Balakhonov, S.V., Dissemination of tryptophan-dependent variants of Yersinia pestis in the territory of the Altai Mountain natural plague focus, Probl. Osobo Opasn. Infekts., 2012, no. 3, pp. 20–25.
Popkov, A.F., Chipanin, E.V., and Korzun, V.M., Population phenetic differentiation of Mongolian pika in South-Eastern Altai, Baikal. Zool. Zh., 2012, no. 1, pp. 114–117.
Rapoport, L.P., Kovtun, I.P., Karpov, A.A., Maleev, A.N., Vetrov, F.E., and Balabas, N.G., On the matter of population structure of the great gerbil range in Muyunkums, in Ekologiya i meditsinskoe znachenie peschanok fauny SSSR (Ecology and Medical Purpose of Pikas of USSR Fauna), Moscow: Institut epidemiologii i mikrobiologii im. N. F. Gamalei Akad. Med. Nauk SSSR, 1977, pp. 86–87.
Rapoport, L.P., Rakhimov, K.R., Putyatin, V.V., Balabas, N.G., Orlova, L.M., Kovtun, I.P., Kondratenko, L.P., and Mochalova, E.P., Concerning asynchrony of plague epizooties in different areas of Moiynkum, Karantin. Zoonozn. Infekts. Kazakh., 2002, no. 5, pp. 83–87.
Sludsky, A.A., Derlyatko, K.I., Golovko, E.N., and Ageev, V.S., Gissarskii prirodnyi ochag chumy (Hissar Natural Plague Focus), Saratov: Sarat. Univ., 2003.
Truphanov, G.V. and Golubev, P.D., On population structure of the mountain souslik Citellus musicus in Central Caucasus, Zool. Zh., 1982, vol. 61, no.1, pp. 96–101.
Verzhutskii, D.B., Activity dynamics of natural focality areas of Tuva natural plague focus, Med. Parazitol. Parazitarn. Bol., 2003, no. 3, pp. 36–39.
Verzhutskii, D.B., Spatial organization of host population and host ectoparasites: theoretical and applied aspects (by the example of the long-tailed souslik and its fleas), Extended Abstract of Doctoral (Biol.) Dissertation, Irkutsk, 2005.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
Translated by E. Martynova
Rights and permissions
About this article

Cite this article
Sludsky, A.A., Boiko, A.V., Lyapin, M.N. et al. A Population-Based Approach to Determining the Borders and Number of Natural Plague Foci. Biol Bull Russ Acad Sci 47, 1437–1441 (2020). https://doi.org/10.1134/S1062359020100222
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1062359020100222