
Abstract
In the period 2001–2016, the authors studied the combined effect of trophic and weather–climatic factors on the population dynamics of three vole-eating bird of prey species on the study area in northern Moscow region, Crane Homland Reserve (56.7573° N, 37.792° E). The numbers of breeding pairs of the Common kestrels (Falco tinnunculus), Long-eared owls (Asio otus) and Short-eared owls (A. flammeus), as well as abudance of small mammals, were counted in the spring and summer months every year. Different indicators characterizing the weather–climatic conditions in the study area over the period of existence of the nesting groups of bird species were calculated based on the data of the nearest meteorological station. The simulation results showed that the numbers of the Long-eared owls depended significantly only on the trophic factor (β = 0.77 ± 0.1). The numbers of breeding pairs of the Short-eared owls and the Common kestrels depended significantly on both factors. The influence of the trophic factor on the change in the numbers of predators was the higher for the Short-eared owl (β ± SE = 1.64 ± 0.16) and the less for the Common kestrel (β = 0.38 ± 0.12). For the Common kestrel, the impact of climatic and weather factors (β = 0.41 ± 0.09) on its numbers slightly exceeded that of the trophic factor. For Short-eared owl, the effect of the climatic factor (β = 0.36 ± 0.09) was 4.5 times less than that of the trophic factor. A combination of temperature and humidity in the spring period can significantly influence the amplitude and frequency of fluctuations in the numbers of the three species of birds of prey under consideration.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
INTRODUCTION
Research on the population dynamics of predators in natural communities is a traditional aim of population ecology (Pianka, 1981; Newton, 2010). Most of the studies carried out in this area have revealed a correlation between the number of predators and the abundance of their main prey (Korpimäki, 1984; Litvin and Ovsyanikov, 1990; Tome, 2003; Sundell et al., 2004; etc.). However, the synchronism of cyclical fluctuations in the “predator–prey” system, determined not only by the prey abundances, is often observed to be interrupted. The reasons for such breaking are not always clear, especially when studies are conducted in small areas.
The reasons for the cyclical nature of the dynamics of the numbers of small mammals are not entirely clear, although these cycles are observed in most regions of the taiga and arctic zones. In recent decades, the dynamics of the numbers of small mammals, which serve as the main prey for most raptor birds and terrestrial predators, have been influenced by large-scale climate change (Stenseth et al., 2002; Korpela, 2014). This, in turn, can negatively affect the population dynamics of predators (Solonen, 2010; Schmidt et al., 2012; Cornulier et al., 2013). For example, in Greenland, climate warming has led to a sharp reduction in the abundance of lemmings and attenuation of cyclic fluctuations in their numbers, which previously caused the possibility of breeding of Snowy owls (Nyctea scandiaca) on this island during the years of high abundance of rodents (Gilg et al., 2009; Schmidt et al., 2012). Similar reactions were observed in other arctic predators: the Arctic foxes (Vulpes lagopus) and Long-tailed Skuas (Stercorarius longicaudus), which survival and breeding success depend on the abundance of lemmings also (Henden et al., 2008; Barraquand et al., 2013).
The direct effect of weather and climate fluctuations on the dynamics of bird numbers on nesting areas has been studied to a lesser extent. Numerous publications deal mainly with the problems of breeding success, changes in the timing of migration, the onset of reproduction, and other phenological phenomena (Sokolov et al., 1999; Both et al., 2005; Sokolov, 2006; Jonzen et al., 2006; Zalakevicius et al., 2006; Tottrup et al., 2010; Guillemain et al., 2013; Volkov et al., 2013, 2016). The possible impact on population dynamics is discussed in many articles, but there are few direct observations so far. In Germany, the nesting density of the Common kestrel (Falco tinnunculus) was largely determined by winter temperatures and snow depth (Kostrzewa, R. and Kostrzewa, A., 1991). In unfavorable years, the birds could not gain enough mass in order to start laying eggs. However, meteorological parameters did not significantly affect other species, for example, the Eurasian buzzards (Buteo buteo), Northern goshawks (Accipiter gentilis), Tengmalm’s owls (Aegolius funereus), and the Golden eagles (Aquila chrysaetos) (Kostrzewa, R. and Kostrzewa, A., 1991; Korpimäki and Hakkarainen, 1991; Steenhof et al., 1997). The Gray heron (Ardea cinerea) in England showed a decrease in the number of nesting populations after severe winters, which were accompanied by prolonged freezing of waterbodies, since the birds could not find enough food (Stafford, 1971).
In recent decades, there has been an interruption of cyclicity in the dynamics of voles abundance in several regions of Western and Central Europe (Cornulier et al., 2013; Korpela, 2014; Millon et al., 2014). The damping of cyclic fluctuations in voles abundance must lead to an imbalance in the relationships in the “predator–prey” systems and, most likely, to the decreased numbers of vole-eating birds. For the territory of Russia, such observations have not yet been published, but one can assume the existence of such trends in the Arctic and temperate regions.
The Common kestrel, Long-eared owl (Asio otus), and Short-eared owl (A. flammeus) are common species of open landscapes and areas partially overgrown with trees and shrubs in the central regions of the European Russia, including agricultural lands. The nesting abundance of these species varies cyclically and is associated primarily with annual changes in the food supply (Cramp, 1985). The diet of all three raptor species is based on voles, primarily Microtus (Korpimäki and Norrdahl, 1991; Volkov et al., 2009; Sharikov et al., 2009). Despite the significant similarities in both diet and choice of nesting and hunting habitats, the interannual fluctuations of breeding numbers of the study species are often not synchronous. It can be assumed that, besides the prey abundance on the number dynamics of the raptor birds also affect other factors, in particular meteorological and climatic factors (Korpimäki, 1992; Solonen, 2010; etc.).
The aim of our research was to study the cumulative impact of trophic and weather–climatic factors on the dynamics of the numbers of the three vole-eating species of birds.
MATERIALS AND METHODS
The studies were conducted 2001–2016 in northern Moscow Region in a 48 km2 study area located in the Crane Homeland Reserve (56.75732° N, 37.79202° E). The numbers of raptor birds were determined every year in the breeding period by mapping their nests, broods, and territories. Mapping was carried out many times during the season: in early spring during the period of vocal activity, in the middle and end of spring during the incubation period, and in summer during feeding of broods that have left nests. In the case of the Short-eared owl that builds nests on the ground, pairs were considered breeding if they had nests or broods or were observed to have regular territorial conflicts with other raptor bird species or corvids, to bring food to nests, or to make regular aerial territorial demonstrations to protect the nesting site. The survey data were applied to maps of scale 1 : 10 000; the location of nests and broods was recorded using GPS. The surveys and stationary observations were organized in such a way that the possibility of recounting the same pairs was excluded. In total, the study uses data on 315 nests/broods and 162 nesting sites.
The abundance of small mammals was estimated by standard methods (Naumov, 1963). Captures were carried out using Hero traps twice a season: in spring after the melting of snow cover and in summer, in the first half of June. The former to a greater extent characterize the number of overwintered small mammals, and the latter characterize the success of their reproduction in early summer. The surveys covered the main habitats used by the study species for hunting and breeding. These are, first of all, various meadow communities and shaded open spaces. Lines of 50 traps were placed for three days; checking was carried out once a day, usually in the morning. In total, the study covered 27 348 trap-days.
The analyzed meteorological indicators were calculated based on the data from the nearest Tver weather station, available from the NOAA Climate Prediction Center (http://www.cpc.ncep.noaa.gov). These include the following: half-monthly average temperatures from the second half of February to the end of May, snow depth in February–April and the amount of precipitation in March–May. The EA-WR index (East Atlantic/West Russia Pattern), which is an integral indicator reflecting large-scale fluctuations in atmospheric phenomena in Eastern Europe, was chosen as a climatic characteristic. It is calculated using data from several dozen weather stations, and numerous studies have shown the influence of these indices on the main hydrometeorological fields in the Atlantic–European region. During the positive phase of the EA-WR, relatively drier and hotter weather is observed in the European regions of Russia (Barnston and Livezey, 1987).
An analysis of the possible dependence of the dynamics of predator abundance on environmental conditions was performed using the method of constructing generalized linear models (GLMs) with a logarithmic connection function and the Poisson distribution of the dependent variable (Zuur et al., 2009). Using the principal components method, we combined the initial indicators into two main generalizing factors—the trophic factor (factor 1) and the climatic factor (factor 2). The trophic factor reflects the situation with the food supply, i.e., the numbers of microtine voles and mouse-like rodents in general in the spring and summer. The EA-WR integral monthly index (March–May), which summarizes the climatic conditions in Eastern Europe through an estimation of the temperature, atmospheric pressure, and precipitation, was chosen as the climatic factor. The number of annually breeding predator pairs in the model territory was used as a dependent variable, and the combined factors were used as independent ones. The correlation of the bird numbers with the mentioned factors was analyzed using the materials only for 2004–2016. To verify the equality of the medians of several samples, the Kruskal–Wallis nonparametric test (H) was used. Statistical data processing was carried out using the Statistica 8.0 program (StatSoft, Inc., 2007).
RESULTS
The Dynamics of Numbers of Raptor Birds
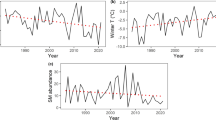
From 2001 to 2016, 4–5 peaks of high numbers abundance were recorded in each of the study species: for the Long-eared owls in 2003, 2007, 2013, and 2016; for the Short-eared owls in 2004, 2007, 2010, 2013, and 2016; for the Common kestrels in 2003, 2010, 2013, and 2016 (Fig. 1). In all three species, the peaks of high abundance coincided only twice, in 2013 and 2016. The Long-eared owls did not have a high abundance in the potentially favorable 2010, and the Common kestrels did not have a high abundance in 2007. The numbers of the Short-eared owls correlated with the numbers of the Long-eared owls (rs = 0.7, p ≤ 0.05) and the Common kestrels (rs = 0.67, p ≤ 0.05). No correlation was found between the numbers of the Long-eared owls and Common kestrels (rs = 0.38, p = 0.15).

Dynamics of numbers of the (1) Long-eared owl, (2) Short-eared owl and (3) Common Kestrel on the study area, in 2001–2016.
In the period analyzed, peaks of high abundance in the Short-eared owls occurred every three years; in the Common kestrel, the interval between peaks was seven years and two times it was three years; in the Long-eared owl, these intervals were four, six, and three years (Fig. 1). The average number (median) of pairs of the Long-eared owls and Short-eared owls breeding in the model territory was six and four pairs, respectively, and this indicator for the common kestrel was seven pairs. In some years, the Long-eared owls and Short-eared owls did not breed in the study area.
The beginning and duration of the breeding period in the species differed. In the Long-eared owl, it was the most extended—the earliest known egg laying dates fell on the end of February, and the latest ones fell on the beginning of June (the median was on March 31, n = 24). In the Short-eared owls, these dates varied from late March to mid-June (the median was on April 9, n = 9), and in the Common kestrel, they varied from the second decade of April to mid-June (the median on May 1, n = 5). Thus, abundance of these three raptors species is formed in the study area from the end of February to the first half of June. Therefore, the wether conditions in this period must most strongly influence the annual number of breeding pairs.
The Influence of the Abundance of Prey on the Numbers of Raptor Birds
In the model area, 14 species of small mammals that are potential prey for the study predators were recorded: four shrew species (Soricomorpha): the Eurasian shrew (Sorex araneus), Eurasian pygmy shrew (S. minutus), Even-toothed shrew (S. isodon), and Laxmann’s shrew (S. caecutiens), Red-toothed shrews, and ten species of rodents (Myomorpha): the Common vole (Microtus arvalis), European field vole (M. agrestis), Tundra vole (M. oeconomus), Bank vole (Clethrionomys glareolus), European water vole (Arvicola amphibius), Pygmy wood mouse (Sylvaemus uralensis), Striped field mouse (Apodemus agrarius), House mouse (Mus musculus), Northern birch mouse (Sicista betulina), and Harvest mouse (Micromys minutus).
The influence of the abundance of potential prey on the numbers of vole-eating birds was analyzed by grouping the species represented in the captures into three cohorts: (1) microtine voles: only representatives of the genus Microtus (three species); (2) mouse-like rodents: microtine voles and all other rodents (eight species); (3) small mammals: mouse-like rodents and shrews (14 species).
The abundance of representatives of each of the three cohorts changed cyclically (Fig. 2). In early summer, peaks of high abundance of mouse-like rodents were observed in 2004, 2007, 2010, 2011, 2013, and 2016; their frequency with rare exceptions was three years. Since 2009, the share of microtine voles in the captures began to decrease, while there was a simultaneous gradual increase in the abundance of Bank voles and Pygmy wood mice, which became common in the mouse-like rodents community on the study area. An increase in the total abundance of potential prey during the years of the peak abundance of small mammals until 2008–2009 occurred mainly due to an increase in the abundance of microtine voles (cohort 1), mainly the Common vole and Tundra vole. After 2012, the peaks in the total number of small mammals were mainly due to the mass spread of the Bank vole and Pygmy wood mouse (cohort 2).

Spring and summer abundance of small mammals on the study area in 2004–2016: 1, cohort of “microtine voles”; 2, cohort of “mouse-like rodents”; 3, cohort of “small mammals.”
The spring abundance of small mammals , during the formation of nesting groups of the raptor birds, significantly affected their numbers (Table 1). For the Long-eared owls the total abundance of mouse-like rodents and small mammals (cohorts 2 and 3) turned out to be the most significant; for the Short-eared owls and Common kestrels, the correlation coefficient with the abundance of microtine voles (cohort 1) was the highest.
In summer, the abundance of microtine voles (cohort 1) remained significant for all three raptor bird species and the abundance of mouse-like rodents (cohort 2) and small mammals (cohort 3) was significant only for owls (Table 1). For Short-eared owls the coefficients of correlation of its abundance with the abundance of all three cohorts of potential prey were somewhat higher in the summer than in the spring. For Long-eared owls and Common kestrels, the values of the correlation index in the summer and spring almost did not differ.
The Influence of Meteorological and Climatic Parameters on the Numbers of Vole-Eating Birds
We calculated various indicators that characterize the weather and climatic conditions in the study area during the nesting period of owls and the Common kestrel and analyzed the influence of these indicators on the their numbers (Table 2). In most cases, no correlation was observed.
A correlation with some weather and climate factors was found only for the numbers of the Common kestrel and Short-eared owl. Colder conditions at the end of February and in March and great depth of snow in March were followed by a higher nesting abundance of the Common kestrel. In addition, its numbers correlated negatively with the value of the climate index in May. The number of territorial pairs of the Short-eared owls correlated negatively with the temperature at the end of February, which is the beginning of the period of prenesting movements in this species, as well as with the amount of precipitation in May during laying and incubation of eggs.
Assessment of the Cumulative Effect of Climatic and Trophic Factors on the Dynamics of Numbers of Vole-Eating Birds
It can be assumed that trophic and meteorological factors affect the numbers of vole-eating birds together, complementing each other. To test this hypothesis, we combined the trophic and weather-climatic parameters into two generalizing factors by the principal components method for further analysis of their possible joint effect on the numbers of birds (Fig. 3). This analysis showed that the numbers of the Long-eared owl and Short-eared owl positively correlated with the spring and summer abundance of microtine voles (cohort 1) and the summer abundance of mouse-like rodents (cohort 2). The numbers of the Common kestrel correlated positively with the spring abundance of mouse-like rodents (cohort 2) and the climate characteristics of March.

The results of combining the parameters of the seasonal numbers of rodents and the EA-WR circulation indices by the principal components method into two factors: the climatic factor (X axis, Factor 1) and the trophic factor (Y axis, Factor 2). (а) Factor loads of the main variables: the spring and summer abundance of microtine voles or the total abundance of mouse-like rodents (Rodentia), as well as the values of loads of EA-WR climatic indices in March, April, and May (the month is indicated by a number). (b) Factor loads of auxiliary variables: 1, the number of breeding pairs of long-eared owls, 2, the number of breeding pairs of short-eared owls, 3, the number of breeding pairs of common kestrels.
All 13 years of observations can be divided into four groups, which differ in various combinations of trophic and weather–climatic factors. The first group includes 2004, 2007, and 2011, which fell into the first sector on the graph (Fig. 4). In these years, the combination of factors that was the most favorable for vole-eating birds was observed: a high abundance of rodents and warm and dry weather. The second sector includes 2010, 2013, and 2016, which were notable for good foraging conditions, but humid and relatively cold weather at the beginning of breeding. The third sector includes 2008, 2009, 2014, and 2015, which were characterized by a low abundance of rodents, but generally favorable weather conditions. The fourth sector groups the seasons, which are characterized by a low abundance of rodents and unfavorable weather characteristics for breeding: low temperature and high humidity (2005, 2006, and 2012).

(1) Characteristics of the 2004–2016 research seasons and distribution of the numbers of (2) Long-eared owl, (3) Short-eared owl, and (4) Common kestrel depending on a combination of the trophic factor (Factor 1) and weather and climate conditions (Factor 2). The points on the graphs correspond to a specific year; the number given next indicates the number of pairs of a given species in a given year.
The application of the Kruskal–Wallis test showed statistically significant differences in the average indices (medians) of the numbers of the Long-eared owls (H (3, 13) = 8.8, p = 0.03), Short-eared owls (H (3, 13) = 10.2, p = 0.02), and Common kestrel (H (3, 13) = 9.4, p = 0.02) between the sectors analyzed (Table 3).
According to the relative suitability of conditions for breeding, sectors 1 and 2 are distinguished when the abundance of all three vole-eating bird species is much higher (Table 3). These seasons are characterized by a high abundance of potential prey. The spring temperature in such seasons can be, on average, both relatively high (Fig. 4, sector 1) and relatively low (sector 2). Meanwhile, the numbers of owls are usually smaller at low temperatures (Table 3).
In the years with a low abundance of potential prey (sectors 3 and 4 in Fig. 4), the numbers of all three vole-eating bird species decreases significantly, by three times or more. Moreover, the response of the studied species to different combinations of temperature and precipitation in such years is somewhat different (Table 3): a greater number of breeding pairs of Long-eared owls begin to breed in dry and relatively warm seasons, and Common kestrels start to breed in wet and cold seasons. Short-eared owls do not breed in the study area in the seasons with insufficient food supply.
In order to evaluate and compare simultaneously the influence of trophic and climatic factors on the numbers of the three raptor species, regression analysis was applied (Table 4). The simulation results showed that the abundance of the Long-eared owls was significantly affected only by the trophic factor. For the Short-eared owls and Common kestrels, both factors were statistically significant. The degree of influence of foraging conditions on the change in the numbers of predators was the greatest for the Short-eared owls and the least for the Common kestrels. For Common kestrels, the influence of climatic factors slightly exceeded the effect of the trophic factor in significance. For Short-eared owls the effect of the climate factor was 4.5 times less than that of the trophic one.
DISCUSSION
Occupying the upper trophic levels, raptor birds form an important part of natural ecosystems. A change in their numbers is a reflection of the processes occurring in natural communities; therefore, it can be used as an indicator of the current state and forecast of the stability of these communities, including in a changing climate.
The Short-eared owl, Long-eared owl and Common kestrel are obligate myophages, in which a significant part of the diet, regardless of the region and season, is made up of small mammals, mainly voles. Most often, they hunt microtine rodents (Korpimäki, 1987; Korpimäki and Norrdahl, 1991; Sharikov et al., 2009); accordingly, the spring and summer abundance of this particular cohort has the greatest influence on the dynamics of numbers of the predators. However, the influence of the trophic factor on the abundance dynamics is not equally pronounced in the bird species studied. The maximum dependence was found for the Short-eared owl, and the smallest dependence was revealed for the Common kestrel. The differences revealed may be due to the different sizes of the food range of these species: in the study area, it is the greatest in the Common kestrel and the smallest in the Short-eared owl (Sharikov et al., 2009; Buslakov, 2012).
The increase in the average monthly temperatures in winter and spring, which has been noted in many regions of Europe, including Moscow region (Volkov et al., 2013), contributes to earlier snowmelt, a shift in the timing of floods, and an earlier start of vegetation. In the Arctic regions, the effects of such climatic changes are the most noticeable and can be very significant, affecting the demographic and spatial characteristics of populations of various species in natural communities (Henden et al., 2008; Gilg et al., 2009; Schmidt et al., 2012; Barraquand et al., 2013; etc.). In the temperate zone, the influence of climate warming on the dynamics of the numbers of small mammals and through them on predators is still poorly studied, but it cannot be ruled out that it also has large-scale consequences. In particular, some researchers attribute the disappearance of cyclic abundance changes in microtine rodents in Western and Central Europe solely to climate warming (Hörnfeldt 2004; Ims et al., 2008; Cornulier et al., 2013). For many species that inhabit very different types of habitats, local causes, such as fragmentation of forests or overgrowing of farmland, cannot fully explain the observed trends of the sharp decrease in abundance or changes in its dynamics (Hörnfeldt, 2004; Hörnfeldt et al., 2006). So, in the United Kingdom, a decrease in the severity of winters led to a shift in the timing of the onset of breeding in sedentary Tawny owls; winter weather also largely determined the breeding success of this species, in particular, the number of successfully grown-up fledglings, although no relationship between the weather and fluctuations in the numbers of owls was established (Millon et al., 2014).
The duration of the snowy period can determine the nature of cyclical changes in the numbers of voles in the temperate zone (Hansson and Henttonen, 1985; Hansson, 1999; Taylor et al., 2013). The shortening of this period is considered to be the reason for the equalization of the amplitude of fluctuations in the abundance of representatives of this group of microtine rodents with the advance to the south. That is, there are indications that their population dynamics are undoubtedly sensitive to climatic factors, although direct correlations are often not found by researchers. It is assumed that the cyclicity of voles abundance in areas where the snow cover lasts longer are more pronounced due to the decrease in the influence of generalist predators (Hansson and Henttonen, 1985; Hansson, 1987; Norrdahl and Korpimäki, 2002) and reduction of the breeding season (Taylor et al., 2013).
In Italy, climate changes have led to the changes of small mammal communities composition in which southern xerophilous species have become increasingly dominant, and this has affected the composition of prey of the barn owl (Szpunar et al., 2008). In the area of our studies, the changes of small mammal communities is also taking place; the Common voles, which previously dominated on the microtine rodent communities of open agricultural landscapes, has recently had low numbers. While until 2009 the peaks in voles numbers were mainly due to outbreaks of the abundance of the Common vole, at the present stage fluctuations in its numbers have a low amplitude and weakly affect the total abundance of microtine rodents. The decrease in the amplitude of fluctuations in this species was mainly determined not by climate change, but by a cardinal restructuring of the habitat structure that took place in the same years, up to the significant overgrowth of habitats with trees and shrubs during the agricultural lands abandonment due to economic crisis (Sviridova et al., 2006, 2016). As a result, the proportion of ecotone and forest species (Bank vole and Pygmy wood mouse) has increased in the small mammals communities. The cyclical dynamics of the numbers of small mammals persists, but the intervals between the years with high abundance have become unpredictable due to changes in the species composition with the persistent amplitude of fluctuations. Observations on abandoned farmland in Spain show similar trends (Torre et al., 2015).
Local meteorological conditions can affect the dynamics of the numbers of predators either directly or indirectly through changes in the numbers of their preys (Millon et al., 2014). However, the influence of winter temperature conditions as well as depth of snow cover on the numbers dynamics of small mammals is often not statistically confirmed (Steenhof et al., 1997; Korpela, 2014). Meanwhile, significant changes in weather conditions in the late winter and spring periods negatively affect the onset dates and success of rodent breeding, causing a noticeable decrease in their numbers (Aars and Ims, 2002; Bierman et al., 2006; Korslund and Steen, 2006; Korpela, 2014).
The demographic indicators of local predator communities are not always associated exclusively with the abundance of the food supply in a given year. This becomes especially noticeable in years with cold and rainy weather, when unfavorable conditions lead to a decrease in the numbers and reproductive success of vole-eating birds of prey. The degree of influence of weather conditions on different species is not the same. For the Long-eared owls, the meteorological conditions in the breeding area are not significant, and for the Short-eared owls and Common kestrels, the weather turned out to be significant in the period that immediately precedes their arrival in the nesting area. The reason for such differences between the species is most likely due to the peculiarities of their life cycles: the Long-eared owls usually migrates relatively short distances from nesting sites, and in some years it may remain for wintering in the study area. Accordingly, the weather conditions at the end of winter and the beginning of spring are not as important for this species, and the birds are guided by the abundance and availability of prey. The Short-eared owl and Common kestrel migrate over longer distances and potentially have more options for choosing the breeding area. Cold weather in spring in the northern parts of their nesting range can delay the birds of these species in the southern regions, including in the area of our research, where some of them begin to breed. A similar situation is observed in some migratory passerines, the breeding numbers of which can locally increase in cold spring conditions (Sokolov, 2006).
Thus, in addition to the abundance of potential prey, the dynamics of the numbers of raptor birds can be affected by climatic and local meteorological factors. The amplitude of fluctuations in the numbers of vole-eating birds of prey is determined primarily by the abundance of prey. However, environmental factors (primarily the combination of temperature and humidity in the spring) can significantly influence both the amplitude of fluctuations in the numbers of vole-eating birds and the frequency of the forming peaks or decreases in the numbers of raptor birds.
REFERENCES
Aars, J. and Ims, R.A., Intrinsic and climatic determinants of population demography: the winter dynamics of tundra voles, Ecology, 2002, vol. 83, pp. 3449–3456.
Barnston, A.G. and Livezey, R.E., Classification, seasonality and persistence of low-frequency atmospheric circulation patterns, Mon. Weather Rev., 1987, vol. 115, pp. 1083–1126.
Barraquand, F., Høye, T.T., Henden, J.-A., Gilg, O., Schmidt, N.M., et al., Demographic responses of a site-faithful and territorial predator to its fluctuating prey: long-tailed skuas and arctic lemmings, J. Anim. Ecol., 2013, vol. 83, pp. 375–387.
Bierman, S.M., Fairbairn, J.P., Petty, S.J., Elston, D.A., Tidhar, D., and Lambin, X., Changes over time in the spatiotemporal dynamics of cyclic populations of field voles (Microtus agrestis L.), Am. Nat., 2006, vol. 167, pp. 583–590.
Both, C., Piersma, T., and Roodbergen, S.P., Climate change explains much of the 20th century advance in laying date of northern lapwing vanellus vanellus in the netherlands, Ardea, 2005, vol. 93, no. 1, pp. 79–88.
Buslakov, V.V., The diversity and variability of foraging of kestrel in the northern part of Moscow environs, in Khishchnye ptitsy v dinamicheskoi srede tret’ego tysyacheletiya: sostoyanie i perspektivy (Birds of Prey in the Dynamic Environment of the Third Millennium: Status and Prospects), Krivoi Rog: FL-P Chernyavskii, D.A., 2012, pp. 420–422.
Cornulier, T., Yoccoz, N.G., Bretagnolle, V., Brommer, J.E., Butet, A., et al., Europe-wide dampening of population cycles in keystone herbivores, Science, 2013, vol. 340, no. 6128, pp. 63–66.
Cramp, S., The Birds of the Western Palearctic, Oxford: Oxford Univ. Press, 1985, vol. 4.
Gilg, O., Sittler, B., and Hanski, I., Climate change and cyclic predator-prey population dynamics in the high arctic, Global Change Biol., 2009, vol. 15, pp. 2634–2652.
Guillemain, M., Pöysä, H., Fox, A.D., Arzel, C., Dessborn, L., et al., Effects of climate change on European ducks: what do we know and what do we need to know?, Wildl. Biol., 2013, vol. 19, no. 4, pp. 404–419.
Hansson, L., Intraspecific variation in dynamics: small rodents between food and predation in changing landscapes, Oikos, 1999, vol. 86, pp. 159–169.
Hansson, L., An interpretation of rodent dynamics as due to trophic interactions, Oikos, 1987, vol. 50, pp. 308–318.
Hansson, L. and Henttonen, H., Gradients in density variations of small rodents—the importance of latitude and snow cover, Oecologia, 1985, vol. 67, pp. 394–402.
Henden, J.-A., Bårdsen, B-J., Yoccoz, N.G., and Ims, R.A., Impacts of differential prey dynamics on the potential recovery of endangered arctic fox populations, J. Appl. Ecol., 2008, vol. 45, pp. 1086–1093.
Hörnfeldt, B., Long-term decline in numbers of cyclic voles in boreal Sweden: analysis and presentation of hypotheses, Oikos, 2004, vol. 107, pp. 376–392.
Hörnfeldt, B., Christensen, P., Sandström, P., and Ecke, F., Long-term decline and local extinction of Clethrionomys rufocanus in boreal Sweden, Landscape Ecol., 2006, vol. 21, pp. 1135–1150.
Ims, R.A., Henden, J., and Killengreen, S.T., Collapsing population cycles, Trends Ecol. Evol., 2008, vol. 23, pp. 79–86.
Jonzen, N., Linden, A., Ergon, T., Knudsen, E., Vik, J.O., et al., Rapid advance of spring arrival dates in long-distance migratory birds, Science, 2006, vol. 312, no. 5782, pp. 1959–1961.
Korpela, K., Biological Interactions in the Boreal Ecosystem under Climate Change—Are the Vole and Predator Cycles Disappearing?, Jyväskylä: University of Jyväskylä, 2014.
Korpimäki, E., Population dynamics of birds of prey in relation to fluctuations in small mammal populations in western Finland, Ann. Zool. Fenn., 1984, vol. 21, pp. 287–293.
Korpimäki, E., Dietary shifts, niche relationships and reproductive output of coexisting kestrels and long-eared owls, Oecologia, 1987, vol. 74, pp. 277–285.
Korpimäki, E., Population dynamics of Fennoscandian owls in relation to wintering conditions and between-year fluctuations of food, in The Ecology and Conservation of European Owls, Peterborough, 1992, p. 1–10.
Korpimäki, E. and Norrdahl, K., Numerical and functional responses of kestrels, short-eared owls, and long-eared owls to vole densities, Ecology, 1991, vol. 72, pp. 814–826.
Korpimäki, E. and Norrdahl, K., Do breeding nomadic avian predators dampen population fluctuations of small mammals?, Oikos, 1991a, vol. 62, pp. 195–208.
Korslund, L. and Steen, H., Small rodent winter survival: snow conditions limit access to food resources, J. Anim. Ecol., 2006, vol. 75, pp. 156–166.
Kostrzewa, R. and Kostrzewa, A., Winter weather, spring and summer density, and subsequent breeding success of Eurasian kestrels common buzzards and northern goshawks, Auk, 1991, vol. 108, pp. 342–347.
Litvin, K.E. and Ovsyanikov, N.G., The dependence of reproduction and abundance of snowy owls and Arctic foxes on the abundance of lemmings on Wrangel Island, Zool. Zh., 1990, vol. 69, no. 4, pp. 52–64.
Millon, A., Petty, S.J., Little, B., Gimenez, O., Cornulier, T., and Lambin, X., Dampening prey cycle overrides the impact of climate change on predator population dynamics: a long-term demographic study on tawny owls, Global Change Biol., 2014, vol. 20, no. 6, pp. 1770–1781.
Naumov, R.L., Organizatsiya i metody ucheta ptits i vrednykh gryzunov (The Organization and Methods of Counts of Birds and Harmful Rodents), Moscow: Akad. Nauk SSSR, 1963.
Newton, I., Population Ecology of Raptors, Berkhamsted: A&C Black, 2010.
NOAA Climate Prediction Center [Electronic resource]. http://www.cpc.ncep.noaa.gov. Accessed December 5, 2017.
Norrdahl, K. and Korpimäki, E., Seasonal changes in the numerical responses of predators to cyclic vole populations, Ecography, 2002, vol. 25, pp. 428–438.
Pianka, E., Evolutionary Ecology, 2nd ed., New York: Harper and Row, 1978.
Schmidt, N.M., Ims, R.A., Høye, T.T., Gilg, O., Hansen, L.H., et al.,Rresponse of an arctic predator guild to collapsing lemming cycles, Proc. R. Soc. London, B: Biol. Sci., 2012, vol. 279, no. 1746, pp. 4417–4422.
Sharikov, A.V., Kholopova, N.S., Volkov, S.V., and Makarova, T.V., Overview of foraging of owls in Moscow and Moscow oblast, in Sovy Severnoi Evrazii: ekologiya, prostranstvennoe i biotopicheskoe raspredelenie (Owls of Northern Eurasia: Ecology and Spatial and Biotopic Distribution), Moscow, 2009, pp. 188–203.
Sokolov, L.V., Impact of global warming on migration and breeding periods of passerine birds in the 20th century, Zool. Zh., 2006, vol. 85, no. 3, pp. 317–341.
Sokolov, L.V., Markovets, M.Yu., Shapoval, A.P., and Morozov, Yu.G., Long-term monitoring in the timing of spring migration of passerine birds in the Curonian Spit, Zool. Zh., 1999, vol. 78, no. 6, pp. 709–717.
Solonen, T., Reflections of winter season large-scale climatic phenomena and local weather conditions in abundance and breeding frequency of vole-eating birds of prey, in Trends in Ornithology Research, New York: Nova, 2010, pp. 95–119.
Stafford, J., Heron populations of England and Wales 1928–1970, Bird Study, 1971, vol. 18, pp. 218–221.
StatSoft, Inc., STATISTICA (data analysis software system), version 8, 2007. www.statsoft.com.
Steenhof, K., Kochert, M.N., and McDonald, T.L., Interactive effects of prey and weather on golden eagle reproduction, J. Anim. Ecol., 1997, vol. 66, pp. 350–362.
Stenseth, N.C., Mysterud, A., Ottersen, G., Hurrell, J.W., Chan, K.S., and Lima, M., Ecological effects of climate fluctuations, Science, 2002, vol. 297, pp. 1292–1296.
Sundell, J., Huitu, O., Henttonen, H., Kaikusalo, A., Korpimaki, E., et al., Large scale spatial dynamics of vole populations in finland revealed by the breeding success of vole eating avian predators, J. Anim. Ecol., 2004, vol. 73, pp. 167–178.
Sviridova, T.V., Volkov, S.V., Grinchenko, O.S., Zubakin, V.A., Kontorshchikov, V.V., et al., Impact of agricultural activity intensity on birds of agrolandscapes of northern Moscow oblast, in Trudy XII Mezhdunar. ornitol. konf. (Proc. XII Int. Ornithol. Conf.), Stavropol’, 2006, pp. 371–399.
Sviridova, T.V., Volkov, S.V., Grinchenko, O.S., and Kol’tsov, D.B., Monitoring of birds and their habitats in agricultural landscapes of northern Moscow oblast: the results of 20 years of observations, in Ptitsy i sel’skoe khozyaistvo: sovremennoe sostoyanie, problemy i perspektivy izucheniya (Bird and Agriculture: Current State, Problems, and Prospects of Research), Moscow, 2016, pp. 268–277.
Szpunar, G., Aloise, G., Mazzotti, S., Nieder, L., and Cristaldi, M., Effects of global climate change on terrestrial small mammals communities in Italy, Fresenius Environ. Bull., 2008, vol. 17, pp. 1526–1533.
Taylor, R.A., White, A., and Sherratt, J.A., How do variations in seasonality affect population cycles?, Proc. R. Soc. London, B: Biol. Sci., 2013, vol. 280, p. 20122714. https://doi.org/10.1098/rspb.2012.2714
Tome, D., Functional response of the long-eared owl (Asio otus) to changing prey numbers: a 20-year study, Ornis Fennica, 2003, vol. 80, no. 2, pp. 63–70.
Torre, I., Gracia-Quintas, L., Arrizabalaga, A., Baucells, J., and Diaz, M., Are recent changes in the terrestrial small mammal communities related to land use change? A test using pellet analyses, Ecol. Res., 2015, vol. 30, no. 5, pp. 813–819.
Tottrup, A.P., Rainio, K., Coppack, T., Lehikoinen, A., Rahbek, C., and Thorup, K., Local temperature fine-tunes the timing of spring migration in birds, Integr. Comp. Biol., 2010, vol. 50, pp. 293–304.
Volkov, S.V., Sharikov, A.V., Basova, V.B., and Grinchenko, O.S., The influence of the abundance of small mammals on the selection of habitats and population dynamics of the long-eared (Asio otus) and short-eared (Asio flammeus) owls, Zool. Zh., 2009, vol. 88, no. 10, pp. 1248–1257.
Volkov, S.V., Grinchenko, O.S., and Sviridova, T.V., Arrival dates of gray crane (Grus grus) in the north of Moscow region and their relation to weather and climate factors, Zool. Zh., 2013, vol. 92, no. 7, pp. 834–840.
Volkov, S.V., Grinchenko, O.S., and Sviridova, T.V., The effects of weather and climate changes on the timing of autumn migration of the common crane (Grus grus) in the north of Moscow region, Biol. Bull. (Moscow), 2016, vol. 43, no. 9, pp. 1203–1211.
Zalakevicius, M., Bartkeviciene, G., Raudonikis, L., and Janulaitis, J., Spring arrival response to climate change in birds: a case study from Eastern Europe, J. Ornithol., 2006, vol. 147, pp. 326–343.
Zuur, A.F., Ieno, E.N., Walker, N.J., Saveliev, A.A., and Smith, G.M., Mixed Effects Models and Extensions in Ecology with R, New York: Springer, 2009.
ACKNOWLEDGMENTS
The authors thank O.S. Grinchenko for comprehensive assistance in conducting field research in the Crane’s Homeland Reserve, as well as A.V. Makarov for the field material on the common kestrel for 2001–2003 and A.A. Lisovskii, who made valuable comments on the structure and content of the manuscript. The authors are also very grateful to all our colleagues, especially students, postgraduates, and employees of the Department of Zoology and Ecology of Moscow State Pedagogical University for their help in collecting and processing the field material.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. This article does not contain any studies involving animals performed by any of the authors.
Additional information
Translated by L. Solovyova
Rights and permissions
About this article

Cite this article
Sharikov, A.V., Volkov, S.V., Sviridova, T.V. et al. Cumulative Effect of Trophic and Weather–Climatic Factors on the Population Dynamics of the Vole-Eating Birds of Prey in Their Breeding Habitats. Biol Bull Russ Acad Sci 46, 1097–1107 (2019). https://doi.org/10.1134/S1062359019090139
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1062359019090139