
Abstract
Unique data were obtained on the abnormally high level of phytoplankton biomass (30 g m–3) in the marginal ice zone of the deep-water High Arctic (the Nansen Basin, 83° N) during observations onboard a ship in the summer of 2020. The changes in species composition and the increase in abundance of plankton phytocenoses were determined by the complex hydrographic structure formed due to the interaction between the warm Atlantic and cold Arctic water masses and confirmed the important role of the North Atlantic Current in the European sector of the Arctic Basin. For the first time, a reliable relationship was established between the phytoplankton productivity and the halocline depth (an indicator of rising Atlantic water enriched with nutrients, primarily nitrogen). It was shown for the first time that the large cryopelagic centric diatom Porosira glacialis (= Podosira hormoides var. glacialis) can be a species forming phytoplankton bloom in the marginal ice zone of the High Arctic.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Warming in the Atlantic sector of the Arctic and a sudden shrinkage of the ice field area is related to intensification of the North Atlantic Current [1, 2]; one of its branches passes through the Nansen Basin and the second one, bifurcating largely in the southern Barents Sea, transfers the Atlantic waters to its eastern boundaries northward and southward of the Novaya Zemlya Archipelago. In the Barents Sea, in addition to ice melting, such intensification caused annual summer bloom of the Atlantic species coccolithophorids Emiliania huxleyi, with an abundance from 1.5 × 106 to 12.0 × 106 cell L–1, which has been recorded over the past 20 years [3] The bloom occurs exclusively in the waters of Atlantic genesis southward from the polar front in the warmest waters of the Norwegian (Nordkapp) Current. In the published works of the past 20 years, the intensification of the Atlantic water (AW) inflow into the Arctic Ocean is commonly known as “atlantification,” which includes mass development (“bloom”) of boreal species brought to the Arctic region by the currents from the North Atlantic [4, 5].
There is very little information on the AW influence on the pelagic ecosystems of the Nansen Basin and, in particular, on the primary producing component of the ecosystem. It is known that, in the midstream of the Atlantic Boundary Current (ABC) passing eastward along the continental slope, the coccolithophorid E. huxleyi can develop abundantly (to 5.0 × 105 cell L–1) [6]. This sharply contrasts with the phytocenoses of the shelf waters around Svalbard archipelago, which are not subject to the influence of this current. Here, the structure of the leading phytoplankton complexes is determined by the cold-water Arctic species [6–8]. The same refers to the northern Barents Sea where the Arctic species are dominant in the summer phytocenoses [9]. Exceptions are the ocean trenches: in particular, subsurface maxima of phytoplankton abundance and biomass were formed by the boreal-Arctic diatom Eucampia groenlandica in the summer of 2017 in the Franz-Victoria trench [10].
The studies performed in the marginal ice zone in the Nansen Basin (83° N, depth of 3000 m) in August 2012 at the section from the east coast of Svalbard to the open sea [8] showed that the plankton phytocenosis at low abundance and biomass of the community was dominated by representatives of “ice flora,” which is cold-water Arctic pennate diatoms from the genera Fragilariopsis, Nitzschia, and Navicula.
The studies of the hydrophysical structure performed in the Nansen Basin later from 2013 to 2017 suggested that the zone of AW influence on the hydrological and glacial regimes advanced along the trajectory of this water occurrence; in this case, the boundary of perennial ices was recorded to shift anomalously northeastward in 2017 [2, 11]. An anomalously high (9.0 × 106 cell L–1) bloom level of coccolithophorids was observed in the same period but in the southern branch of the North Atlantic (Norwegian) Current, being a part of the southwestern Barents Sea [3, 12]. The mass development of another Atlantic (Haptophyta) species, the Phaeocystis pouchetii, was recorded for the first time at the eastern boundary of the sea, in the Kara Strait, at approximately the same time [13]. In the summer of 2016, a high abundance of boreal phytoplankton, including coccolithophorids [14], which were likely to come here with the activated Kolguev Current, was found in the St. Anna Trench at the northern extremity of Novaya Zemlya archipelago. Judging by the results of other researchers, the development of the boreal flora (to 1.6 × 105 cell L–1, Chaetoceros compressus) in the southern part of the St. Anna Trench was also recorded to occur earlier, in 2007 and 2011 [15, 16].
At the same time, for the Nansen Basin, there are no data on the response of phytoplankton to the intensification of the AW inflow. The purpose of this work is to reveal the patterns of forming an unusually high bloom intensity in the marginal ice zone based on the comparative analysis of the structure of plankton phytocenoses in the Nansen Basin and the adjacent shelf of the Barents Sea, which is under the direct influence of the ABC.
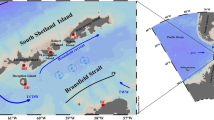
This work was carried out during the 80th cruise of R/V Akademik Mstislav Keldysh on August 16–19, 2020 [17]. The study of the structure of phytoplankton as a regulator of the global carbonate cycle was an integral part of the works on the study of suspended matter flows in the European Arctic seas, which are affected by climate change. The water samples from the surface to a depth of 3000 m were collected using an SBE oceanographic complex at seven stations in the Nansen Basin (82°–83° N) and on the adjacent Barents Sea shelf (80°–81° N) (Fig. 1, Table 1). A total of 80 phytoplankton samples were analyzed. The standard methods accepted at the Institute of Oceanology, Russian Academy of Sciences, were used during the cameral treatment of the samples. Formalin served as a fixative up to a final concentration of 0.5–1% in the sample. The species were identified, and the number of cells was counted on a light Carl Zeiss Ergaval microscope in a water-based preparation at 16 × 20 and 16 × 40 magnification. The representatives of all taxonomic and size groups except for algal picoplankton with linear sizes <2 μm were considered.

(1) Schematic map of phytoplankton sampling stations in August 2020. (2) The boundary of the marine ice sheet is provided from [http://www.aari.ru/odata/_d0015.php].
RESULTS AND DISCUSSION
In the anomalously warm summer period of 2020 for the Arctic, the boundary of the ice fields in the Nansen Basin was recorded at 83° N (Fig. 1, Table 2). The detailed (from the surface to 3000 m) examination of the species and quantity structure of plankton phytocenoses in the marginal zone (stations 6860 and 6861, Table 1) allowed us to establish intensive bloom in the upper freshened water layer at st. 6860, which was formed by large cryopelagic centric diatom Porosira glacialis (= Podosira hormoides var. glacialis Grunow, 1884) (3.5 × 105 cell L–1, 29.9 g m–3, 99% of the total wet biomass of phytoplankton, 13 mg m–3 of chlorophyll “а”, 64.1 mg С m–3 day–1 is primary production). This species belongs to representatives of the so-called “ice flora,” the life cycle of which is partly associated with the low surface of the one year sea ice.
The cells of this species concentrated in a narrow layer at a 5- to 10-m depth in the upper part of the sharply gradient halocline at a temperature of –1.36°C and salinity of 33.4 psu. The phytocenosis occurred at an early stage of settling because the nutrients in the surface five-meter layer of melt water were consumed almost completely (Table 2). The bloom was accompanied by an insignificant development of other cold-water diatoms of the genera Thalassiosira, Lauderia, Chaetoceros, Nitzschia, and Pseudonitzschia and dinoflagellates of the genera Gymnodinium, Gyrodinium, Protoperidinium, and genera Gymnodinium. Almost all phytoplankton were concentrated in the upper 40-m water layer within the photic zone. In the aphotic zone, starting from a depth of 50 m and to a depth of 3000 m, the community contained primarily spores of Thalassiosira gravida and Porosira glacialis, as well as empty frustule of Thalassiosira spp. The similar distribution in the aphotic layer was typical of all deep-water stations. We note especially the presence of the diatom P. glacialis (3.0 × 102 cell L–1) in the AW core at st. 6862.
Plankton phytocenoses of stations 6860 and 6862 (the second was located on the continental slope in the zone of the main flow of the ABC) demonstrated a high degree of similarity (>50% of common species). These phytocenoses included the AW species indicator coccolithophorid E. huxleyi. The representatives of the Atlantic oceanic complex [18] from the genera Rhizosolenia (R. hebetata f. semispina, R. hebetata f. hebetata, R. styliformis, R. setigera) and Chaetoceros (C. borealis, C. atlanticus, C. danicus) were encountered everywhere. A shallow-lying nutricline (3.82 μM of nitrate nitrogen, a horizon of 20 m, Table 2) and a sharply-gradient halocline (a horizon of 5–10 m), separating the thin surface layer of melt water from the lower Arctic waters, as well as the core of the warm AW (temperature from 1.22 to 2.53°С, salinity above 34.8 psu) at the horizons of 100–200 m, were recorded at st. 6860. All the above facts are convincing evidence of the ABC water presence in the Nansen Basin and their rise to the surface. For example, the centric diatom R. hebetata f. semispina, a summer mass form of the boreal Atlantic, was encountered at st. 6860 even in the layer of 5–10 m. This is explained by the fact that the occurrence depth of the halocline in the marginal ice zone varied during the day and could rise to a horizon of 5 m (Fig. 2).

Occurrence depth of the halocline in the Nansen Basin according to the data of STD soundings on August 17–18, 2020: (2, 4) st. 6860 and (1, 3) st. 6861.
By contrast, at st. 6861, which was also located in the marginal ice zone, the temperature and salinity being lower than at st. 6860 (Table 2) but the same type (P. glacialis) being dominant, bloom did not occur. The abundance and biomass of the community were low, the biomass did not exceed 30.6 mg m–3 (1 mg m–3 chlorophyll “а”) and was maximum at a horizon of 3 m, T = –1.43°С, and salinity = 31.3 ups (Table 2). Such considerable differences could be caused by the weaker influence of the ABC. Here, the sharply gradient halocline lay lower (14–20 m), and the nutricline occurred at 34 m (Table 2).The weak impact produced by the ABC on the phytocenosis structure at st. 6861 can also be indicated by the low (<20%) degree of similarity between the species structure at st. 6861 and st. 6862: the occurrence of the Atlantic Ocean complex species (Rhizosolenia spp., Chaetoceros spp.) at a great depth (34–90 m) and the presence of the indicator species (coccolithophorid Coccolithus pelagicus) only in the ABC core (the horizon of 185 m) at st. 6861. Nevertheless, the high physiological activity of cells in the surface water layer at st. 6861 (primary production of 30.3 mg С m–3 day–1, horizon of 1 m) could have held the potential for the bloom increase upon the intensification of the ABC impact. In August 2012, there was no “bloom ” in the marginal ice zone of this area [8]. The phytocenosis was dominated by cold-water Arctic species, primarily from the genus Fragilariopsis. The АW core was located much deeper than in 2020, at a depth of 600 m, which could have caused the noticeable difference in the structure of plankton phytocenoses of different years.
The comparative analysis of the phytoplankton structure in the study area indicates that on the northern Barents Sea shelf (stations 6854, 6856, 6864, and 6866) and near the continental slope (st. 6862), there were later stages of phytoplankton succession determined by the water column stratification due to summer heating of the surface layer. The depletion of nutrients in the upper producing layer results in the termination of diatom bloom followed by its descent to the underlying layers. For example, the subsurface maximum abundance and biomass of diatoms from the genera Eucampia and Thalassiosira and Phaeocystis pouchetii located at the bottom of the photic zone and associated with the nutricline were recorded at shelf stations 6854 and 6856, as well as at st. 6862 (Table 2). Such maxima are a typical characteristic of summer phytocenoses on the northern Svalbard shelf under the ABC impact [8].
In the vicinity of the Franz Josef Land archipelago (stations 6864 and 6866), the phytoplankton development was limited by the low nutrients concentrations, primarily zero concentrations of nitrate nitrogen consumed during the spring bloom of diatoms (Table 2). The mixotrophic and heterotrophic dinoflagellates from the genera Gymnodinium and Gyrodimium dominated in the community. At st. 6864, where the ABC core was markedly pronounced, the AW indicators were coccolithophorid E. huxleyi (3.2 × 103 cell L–1, the layer of 0–20 m) and the warmwater large dinoflagellate Ceratium horridum, which was encountered at a depth of 20 m at the boundary between the polar surface and the Arctic intermediate water masses [8]. Extremely low abundances of phytoplankton were recorded at st. 6866 (as much as 4.6 mg m–3, the horizon of 20 m) compared to the remaining stations, where the presence of the AW core with a salinity of 34.8 psu was the least pronounced: the highest temperature for the layer of 100–200 m was recorded at the horizon of 111 m (–0.02°C, Table 2).
Thus, the results of the studies performed in the summer of 2020 are indicative of cardinal restructuring of plankton phytocenoses in the marginal ice zone, which occurred in the dominants upon a sharp increase in abundance of the new dominant species, the boreal Arctic P. glacialis, to the bloom level. This species replaced the cold-water Arctic species of the “ice flora” from the genera Fragilariopsis, Nitzchia, and Navicula, which dominated in the plankton phytocenoses of the marginal ice zone of this segment in the Nansen Basin in August 2012 [8]. Interestingly, the unprecedented high level of community biomass (30 g m–3) was reached due to the mass development of one species (P. glacialis), which was widespread in the North Atlantic and in those areas of the boreal zone that are characterized by the occurrence of a seasonal ice sheet. Taking into account the discovery of this species in the ABC core at st. 6862, we suggest that it was transported by the waters of this current to the High Arctic.
The discovered phenomenon of mass development of Porosira glacialis in the Nansen Basin is by its nature a spring bloom that is developed in the marginal ice zone during ice melting and is usually observed in the High Arctic in July–August. At such high latitudes (83○ N), the phenomenon of spring bloom was registered in situ for the first time. Increased productivity in the marginal ice zone was recorded earlier only by the satellite data [19]. The bloom was initiated by the active influence of the ABC waters. The influence of these waters was traced up to the water surface, which led to the formation of the shallow sharply-gradient halocline that separated the melt waters from the underlying layers. The absence of bloom at st. 6861, which was also located in the marginal ice zone, can be explained by the weak ABC influence.
The ABC impact in August 2020 was traced on the phytocenoses of both the shelf stations and over the continental slopes. In the absence of an ice sheet, the subsurface maxima abundance and biomass were formed in the 30- to 50-m layer, primarily due to the diatom Eucampia groenlandica, the species forming spring bloom in the subpolar North Atlantic and the Arctic in the open waters. The areas where the ABC impact was minimal (st. 6866) were where the lowest level of quantitative development of phytoplankton was recorded (4.6 mg m–3).
CONCLUSIONS
The first in-situ data were obtained on the intense bloom of diatoms with a biomass of 30 g m–3 in the marginal ice zone (83° N) of the Nansen Basin resulting from the active influence of the Atlantic water.
It was shown for the first time that cryopelagic large centric diatom Porosira glacialis can be a species forming blooms in the marginal ice zone in the High Arctic. At high abundances (3.6 × 105 cell L–1), the contribution of this alga to the total biomass reached 99%.
It was established that the formation of the shallow sharply-gradient halocline separating the melt waters from the underlying waters and causing the Atlantic water to rise to the surface was the main trigger of the phytoplankton bloom.
REFERENCES
I. V. Polyakov, A. V. Pnyushkov, M. B. Alkire, et al., Science 536, 285–291 (2017).
V. V. Ivanov and I. A. Repina, Izv., Atmos. Ocean. Phys. 54 (1), 65–73 (2018).
L. Oziel, A. Baudena, M. Ardyna, et al., Nat. Commun. 11, 1705 (2020). https://doi.org/10.1038/s41467-020-15485-5
J. E. Søreide, E. Leu, J. Berge, et al., Global Change Biol. 16, 3154–3163 (2010).
G. Neukermans, L. Oziel, and M. Babin, Global Change Biol. 24 (6), 1–9 (2018).
J. Giraudeau, V. Hulot, V. Hanquiez, et al., J. Mar. Syst. 158, 93–105 (2016).
G. Owrid, G. Socal, G. Civitarese, et al., Polar Res. 19 (2), 155–171 (2000).
H. Hop, P. Assmy, A. Wold, et al., Front. Mar. Sci. 6, 181 (2019). https://doi.org/10.3389/fmars.2019.00181
P. R. Makarevich and A. A. Oleinik, Tr. Kol’sk. Nauchn. Tsentra Ross. Akad. Nauk 8 (2–4), 50–58 (2017).
L. A. Pautova, in The Barents Sea System, Ed. by A. P. Lisitsyn (Nauchn. mir, Moscow, 2021), pp. 317–330 [in Russian].
P. V. Aksenov and V. V. Ivanov, Probl. Arkt. Antarkt. 64 (1(115)), 42–54 (2018).
V. Silkin, L. Pautova, M. Giordano, et al., Mar. Pollut. Bull. 158, 111392 (2020). https://doi.org/10.1016/j.marpolbul.2020.111392
A. F. Sazhin, S. A. Mosharov, N. D. Romanova, et al., Oceanology 57 (1), 222 (2017).
V. M. Sergeeva, I. N. Sukhanova, M. V. Flint, et al., Oceanology 60 (4), 458–473 (2020).
I. N. Sukhanova, M. V. Flint, E. I. Druzhkova, et al., Oceanology 55 (4), 547–561 (2015).
V. M. Sergeeva, I. N. Sukhanova, E. I. Druzhkova, et al., Oceanology 56 (1), 107–114 (2016).
A. A. Klyuvitkin, N. V. Politova, A. N. Novigatsky, and M. D. Kravchishina, Oceanology 61 (1), 139–142 (2021).
E. Paasche, Rep. Norw. Fish. Mar. Invest. 12 (11), 1–77 (1960).
M. Perrette, A. Yool, G. D. Quartly, and E. E. Popova, Biogeosciences 8, 515–524 (2011).
ACKNOWLEDGMENTS
The authors are grateful to the crew of R/V Akademik Mstislav Keldysh, as well as thank N.V. Politova, A.N. Novigatskii, A.L. Chultsova, A.S. Shchuka, I.A. Zamyatin, and I.I. Rusanov for their assistance in water sampling and analysis.
Funding
The analysis of phytoplankton samples and hydrological–hydrochemical data was supported by the Russian Foundation for Basic Research (project no. 19-05-50090). The expedition research was funded by the State Budget (financing of marine expeditions) and by the Russian Science Foundation (project no. 20-17-00157). This work was carried out under the state task on subject no. 0128-2021-0009.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Translated by M. Hannibal
Rights and permissions
About this article

Cite this article
Pautova, L.A., Silkin, V.A., Kravchishina, M.D. et al. Pelagic Ecosystem of the Nansen Basin under the Influence of Variable Atlantic Water Inflow: The Mechanism Forming Diatom Bloom in the Marginal Ice Zone. Dokl. Earth Sc. 499, 590–594 (2021). https://doi.org/10.1134/S1028334X21070138
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1028334X21070138