
Abstract
This study investigated the polymorphism of hypothalamic-pituitary axis genes and their effects on milk production traits. To this end, we used genomic information of 134 animals. The studied genes were prolactin (PRL), growth hormone (GH), pituitary transcription factor (Pit-1), and signal transducers and activators of transcription (STAT5a). Genomic DNA was isolated from semen, and the existing genotypes for all studied genes were identified using PCR-RFLP assay. The frequencies of the genotypes AA, AG, and GG of PRL-RsaI, LL, LV, and VV of GH-AluI, AA, AB, and BB of Pit1-HinfI, and TT, TC, and CC of STAT5a-AvaI were 0.754, 0.246, and 0, 0.873, 0.127, and 0, 0.142, 0.358, and 0.500, and 0, 0.261, and 0.739, respectively. The genotypes of PRL-RsaI, GH-AluI, and STAT5a-AvaI were in the Hardy–Weinberg equilibrium (P > 0.05), except for Pit1-HinfI (P < 0.05). The association of Pit1-HinfI and STAT5a-AvalI gene polymorphisms with milk fat yield (MFY) and milk protein yield (MPY) was significant (P < 0.05). However, no effective association (P < 0.05) was observed between genotypes and other studied traits. The results suggest that Pit1-HinfI and STAT5a-AvalI polymorphisms can be used as possible candidates in selecting some milk protein production traits, such as MFY and MPY.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The main goal of the dairy cattle industry is to improve milk quantity and quality through selection programs. In addition to conventional selection methods based on phenotype information, marker-assisted selection methods can be used as a fundamental strategy in animal breeding programs [1]. Although milk yield and composition traits are polygenic traits, the effects of some candidate genes, such as prolactin (PRL), growth hormone (GH), pituitary transcription factor (Pit-1), and signal transducer and activator of transcription 5A (STAT5A), on the physiological pathway of those traits have been identified [2–4]. Such genes are related to endocrine systems that have a physiological influence on milk yield and composition.
The PRL gene consists of 5 exons and 4 introns and encodes a polypeptide with 199 amino acids associated with reproduction [5, 6], mammary gland growth, lactation initiation, milk yield and content [6–11]. Then, PRL could be an outstanding candidate gene to affect milk production traits [9, 12, 13]. The GH gene contains 5 exons [14] and encodes a single chain polypeptide with 191 amino acids released from the pituitary gland. This gene is essential for growth, fertility, mammary gland development, and lactation process [2, 15–17]. The GH gene polymorphisms and their associations with production traits, especially milk yield characteristics, have been investigated by various researchers [8, 13, 18–27]. The Pit1 with 6 exons [28] encodes a protein with 291 amino acids containing DNA binding POU domain [29]. This gene activates bovine GH, thyrotrophin ß, and PRL genes [2] and is considered a candidate gene for regulating the expression of milk protein genes [30–33]. The STAT5A gene with 19 exons encodes a protein with 794 amino acids [34]. The STAT5A gene, as a member of the JAK-STAT signaling pathway, has a leading role in developing mammary glands, secreting milk, regulating lactation, and resisting infections, such as mastitis in bovine [35–38]. Moreover, the JAK-STAT pathway regulates the casein gene and balances GH and milk protein contents [39]. Additionally, STAT5A is used by the PRL gene as a mediator for the lactation process in mammals [40].
According to the abovementioned facts about the hypothalamic-pituitary axis candidate genes, in this study, we examine polymorphisms of the PRL-RsaI, GH-AluI, Pit1-HinfI, and STAT5a-AvaI gene polymorphisms and their associations with milk production characteristics, including milk yield (MY), milk fat yield (MFY), milk protein yield (MPY), milk fat percentage (MFP), and milk protein percentage (MPP).
MATERIALS AND METHODS
DNA Samples
In this study, 134 frozen semen related to proven Iranian Holstein and Milk Production characteristics records of 187,481 individuals from 1992 to 2015 were provided by the Animal Breeding Center of Iran. These animals were under the recording system and progeny test program of this center. DNA was extracted from frozen semen straws according to Zadworney and Kuhnlein’s protocol [41]. The quality of the extracted DNA was evaluated by 1% agarose gel and spectrophotometer based on absorbance at 260 nm/280 nm.
DNA Amplification and PCR-RFLP
A PCR-RFLP method was used to determine the gene’s polymorphisms. The detailed information about the target regions on the genome and their primers and enzymes for PCR reactions is presented in Table 1. A PCR for all studied genes was conducted in 25 µL containing 50 ng DNA, 10× reaction buffer (16 mM (NH4)2SO4, 67 mM Tris-HCl pH 8.8, 0.1% Tween-20), MgCl2 (2.5 mM), dNTP (200 µM), primers (5 pmol), and Taq polymerase (1 unit belonging Metabion, Germany). The amplification conditions of the studied sites are depicted in Table 2.
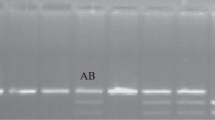
The digestion of PCR products was carried out in 15 µL consisting of PCR product (10 µL), Tango buffer (1.5 µL), and nuclease-free water (3.15 µL), which were similar for all genes plus RsaI (5 units Biolabs, New England) for PRL, AluI (4 units Metabion, Germany) for GH, HinfI (10 units) for Pit-1, and AvaI (7 units Fermentase, Germany) for STAT5A. RFLP fragments were distinguished on 2.5% for PRL, Pit-1, and STAT5A and 3% for GH and were then visualized on an agarose gel.
Statistical Analysis
The studied loci’s allele and genotype frequencies and HWE tests were calculated using GENALEX 6.4 software [42]. Table 3 shows the summary statistics for pedigree and milk production data sets. The production records of first were collected from 1992 to 2015 on 74,213 to 187,481 animals for different traits (Table 3). The breeding values (BV) of genotyped bulls for milk production traits, including MY and its MFY, MPY, MFP, and MPP contents of first lactation production period, were predicted based on a sire model using the AIREML procedure in MATVEC software [43]. The analysis model included herd-year-season as a fixed, animal as a random and calving age as a covariate effect. A fixed model was used to associate genes polymorphisms and estimated BV of the studied traits using the GLM procedure in SAS 9.4 [44]. The Tukey–Kramer’s test was used to compare the least square mean results. The statistical fitted model was:
where yijk is the dependent variable (Estimated BV), µ is the overall population mean for each trait, yeari is the ith birth-year, gj(k) is the jth genotype (j = 1, 2, 3) for kth gene (PRL, GH, Pit-1 and STAT5a), and eijk is the residual effects.
RESULTS AND DISCUSSION
A pedigree with 6 generations back was used in BV estimation with 42 to 3033 daughter records for each evaluated bull. The BV reliabilities ranged from 79.40 to 99.08 percent. The relationship among the evaluated bulls ranged from 0.0001 to 0.3900. All studied genotypes, except for Pit1-HinfI, reached the Hardy–Weinberg equilibrium (P > 0.05).
Allele Frequency
The RFLP results showed that all studied sites were polymorphic. The allele frequencies of the genes are shown in Table 4. Based on the table, the frequencies of alleles A and G of PRL-RsaI, L and V of GH-AluI, A and B of Pit1-HinfI, and T and C of STAT5a-AvaI are 0.877 and 0.123, 0.937 and 0.063, 0.321 and 0.679, and 0.131 and 0.869, respectively.
An allele frequency (0.87) for PRL-RsaI was in the same range reported by Çitek et al. [45] from 0.81 to 0.90 for five studied cattle breeds, Alipanah et al. [19], and higher than 0.53 in Jersey crossbred [6], 0.65 in Holstein [9], and 0.6 in 1198 Indian dairy cows, which were calculated based on a meta-analysis of 15 published studies [26].
The L allele frequency of GH-AluI was reported to be 0.85, 0.93, and 0.51 in Danish Red, Holstein, and Jersey, respectively [46], 0.884 in Holstein [4], and 0.52 in Jersey [20], which are in line with our findings (L allele frequency of 0.937) (Table 4).
The A allele frequency (0.321) was lower than the B allele for Pit1-HinfI, which agrees with previous studies reporting from 0.05 in dairy Gyr breed to 0.41 in East Anatolian Red cattle [2, 47–50]. The frequency of STAT5a-AvaI alleles was in line with the one in previous studies [3, 51].
Polymorphisms Association with Milk Production Traits
The results of the association analysis between the genes and milk production traits are shown in Table 5.
PRL-RsaI Polymorphisms
No significant (P > 0.05) associations were found between the three genotypes of PRL-RsaΙ with studied traits. Numerous studies were carried out on PRL-RsaI polymorphisms and their association with milk production traits in different cattle breeds [9, 52, 53]. Our results contrast with the studies on PRL-RsaI polymorphisms and their association with milk production traits (in all or at least in some studied traits) in different cow breeds [9, 52–54]. Dybus et al. [52] studied five milk production traits in three lactation periods of Jersey and Black-and-White cattle breeds. They found that Jersey cows with AA genotype in the first lactation had lower MFY and MFP. In another study, Dybus et al. [55] found a significant effect of PRL-RsaI polymorphisms on MPY. Oğuzkan and Bozkurt [9] studied the association of MY, MFP, and MPP traits with PRL-RsaI polymorphisms. They observed a higher MY for AA genotype in Holstein cattle [9], and similar results were obtained for MY [6, 55].
GH-AluI Polymorphisms
The GH-AluI polymorphisms have been introduced as a potential candidate gene for milk yield and meat production traits. Previous studies have confirmed this fact for body weight [56, 57]. However, the association of GH-AluI with milk production traits varied from no association [58–61] to significant association with higher milk yield to LL genotype [4] or higher performance for other genotypes [50, 62]. A comprehensive meta-analysis of GH-AluI polymorphism effects on milk production traits by Akcay et al. [63] revealed no association between GH-AluI polymorphisms and milk production traits. Our results agree with those of Akcay et al. [63] and other previous studies [26, 58–61].
Pit1-HinfI Polymorphisms
In this study, significant associations (P < 0.05) were detected between Pit1-HinfI gene polymorphisms and MFY and MPY traits. For both traits (MFY and MPY), individuals with AB genotype yielded the highest performance, and the lowest performance for MFY and MPY was related to the AA genotype. The allele substitution means effect (B instead of A) was estimated to be 1.12 ± 0.99 kg for MFY. These confirm those of other studies that found significant associations of Pit I-HinfI genotypes with MFY and MPY. Renaville et al. [48] showed the association of Pit I-HinfI genotypes with MFY in Italian Holstein-Friesian bulls. De Mattos et al. [2] in Gyr cows detected two genotypes with significant differences between daughter MFY deviations of AB, BB, and AB genotypes. By contrast, Pozovnikova et al. [64] found no significant association for MFY. The association of Pit I-HinfI genotypes with MPY in this study is consistent with the findings of Renaville et al. [48] and Dybus et al. [65] and is inconsistent with those of Pozovnikova et al. [64].
Furthermore, the associations of Pit I-HinfI with MY, MFP, and MPP were not significant. This finding agrees with the conclusions made by Alejandra et al. [66], Pozovnikova et al. [64], and Anggraeni et al. [67] for MY, and Dybus et al. [65], Gorbani et al. [68], and Daniela et al. [69] for milk production traits. Nonetheless, Zabeel et al. [32] and Carșai et al. [70] were reported an association between Pit I-HinfI and MY.
STAT5a-AvaI Polymorphisms
We detected significant associations (P < 0.05) between STAT5a-AvaI gene polymorphisms and MFY and MPY traits. More specifically, the individuals with TC genotype had a higher MFY and MPY than those with CC genotype for MFY and MPY traits. Nonetheless, no individuals were observed for TT genotype.
The allele substitution means effect (T instead of C) was 2.88 and 3.7 kg/year for MFY and MPY, respectively. These results are consistent with those reported by Selvaggi et al. [3] and Dario and Selvaggi [51], who found a significant difference between CC and TC genotypes for MFY (P < 0.01) and a higher MFY in CC compared to TC genotypes. Our results for MPY agree with those of Bao et al. [71], Cosier et al. [72], and Dario and Selvaggi et al. [51], who observed a higher MPY for CC than for CT genotype.
The observed difference between STAT5a-AvaI genotypes and MY was not significant (P < 0.05). This result is consistent with the findings of He et al. [73], Al-Azzawi and Al-Dulaimi [74], and Metin al. [23]. In contrast with our findings, Selvaggi et al. [3] found a significant difference (P < 0.01) between the CC and CT genotypes for MY. Another study by Dario and Selvaggi [51] demonstrated a higher MY for CC than CT genotypes. In the present study, while no significant differences were found between the genotypes of MFP and MPP, the individuals with TC genotype, instead of CC, had lower MFP and MPP. These results are in agreement with those of Selvaggi et al. [3], Dario and Selvaggi [51], Al-Azzawi and Al-Dulaimi [74], and Metin et al. [23]. However, He et al. [73] found significant associations between STAT5a-AvaI and MPP and introduced this site as a potential candidate gene for MPP selection.
CONCLUSIONS
In this study, the loci, including PRL-RsaI, GH-AluI, Pit1-HinfI, and STAT5a-AvaI, were selected due to their importance in functional and economic traits in livestock. Although the effect of these genes on milk production traits in various cattle breeds has been studied previously, the results were inconclusive. Therefore, we examined the polymorphisms of genes and their associations with milk production characteristics. Based on the results, the association of Pit1-HinfI and STAT5a-AvalI gene polymorphisms with MFY and MPY was significant (P < 0.05). However, no effective association (P < 0.05) was observed between genotypes and other traits. According to our results, Pit1-HinfI and STAT5a-AvalI polymorphisms can be used as possible candidates in selecting some milk production traits, such as MFY and MPY. However, further associational studies and comprehensive Meta-analyses are warranted to understand the exact role of the studied genes in milk production traits.
REFERENCES
Dekkers, J.C., Commercial application of marker- and gene-assisted selection in livestock: strategies and lessons, J. Anim. Sci., 2004, vol. 82, E-suppl., pp. E313—E328.
De Mattos, K., Del lama, K.S.N., Martinez, M.L., and Freitas, A.F., Association of bGH and pit_1 gene variants with milk production traits in dairy Gyr bulls, Resq. Agropec. Bras., 2004, vol. 39, no. 2, pp. 147—150.
Selvaggi, M., Dario, C., Normanno, G., Celano, G.V., and Dario, M., Genetic polymorphism of STAT5A protein: relationships with production traits and milk composition in Italian Brown cattle, J. Dairy Res., 2009, vol. 76, pp. 441—445.
Heidari, M., Azari, M.A., Hasani, S., et al., Effect of polymorphic variants of GH, Pit_1, and ß_LG genes on milk production of Holstein cows, Russ. J. Genet., 2012, vol. 48, no. 4, pp. 417—421.
Ladani, D.D., Pipalia, D.L., Brahmkshtri, B.P., et al., PCR RFLP polymorphism at prolactin locus in buffaloes, Buffalo J., 2003, vol. 2, pp. 237—242.
Gayari, I., Ferdoci, A.M., Aziz, A., et al., Prolactin gene polymorphism in crossbred cattle of Assam and its association with productive and reproductive traits, Int. J. Chem. Stud., 2020, vol. 8, no. 2, pp. 717—722.
Brym, P. and Kaminski, S., Nucleotide sequence polymorphism within exon of the bovine prolactin gene and its associations with milk performance traits, J. Appl. Genet., 2004, vol. 5, no. 2, pp. 179—185.
Hernández-Cordero, A.I., Sánchez-Castro, M.A., Zamorano-Alg, R., et al., Genotypes within the prolactin and growth hormone insulin-like growth factor-I pathways associated with milk production in heat stressed Holstein cattle: genotypes and milk yield in heat stressed Holstein cows, Genet. Mol. Res., 2017, vol. 16, no. 4, p. 16039821. https://doi.org/10.4238/gmr16039821
Oğuzkan, S.B. and Bozkurt, A.S., A study on the effect of prolactin gene variants on milk production traits of Holstein cattle, Russ. J. Genet., 2019, vol. 55, no. 4, pp. 480—486.
El-Magd, M.A., Fathy, A., Kahilo, K.A., et al., Polymorphisms of the PRLR gene and their association with milk production traits in Egyptian buffaloes, Animals, 2021, vol. 11, no. 5, p. 1237.
Shah, R.M., Ganai, N.A., Sheikh, F.D., et al., Exon IV prolactin (PRL) gene polymorphism and its association with milk production traits in dairy cattle of Kashmir, India, J. Entomol. Zool. Stud., 2021, vol. 9, no. 2, pp. 521—524.
Sodhi, M., Mukesh, M., Mishra, B.P., et al., Analysis of genetic variation at the prolactin-RsaI (PRL_RsaI) locus in Indian native cattle breeds (Bos indicus), Biochem. Genet., 2011, vol. 49, pp. 39—45.
Hernández-Cordero, A.M., Parra-Bracamonte, G.M., Segura-Correa, J.C. and Magaña-Monforte, J.G., Effect of leptin, pituitary transcription factor and luteinizing hormone receptor genes polymorphisms on reproductive traits and milk yield in Holstein cattle, Braz. Arch. Biol. Technol., 2021, vol. 64. e21190643.
Gordon, D.F., Quick, D.P., Ewin, C.R., et al., Nucleotide sequence of the bovine growth hormone chromosomal gene, Mol. Cell. Endocrinol., 1983, vol. 33, pp. 81—95.
Tatsuda, K., Oka, A., Iwamoto, E., et al., Relationship of the bovine growth hormone gene to carcass traits in Japanese black cattle, J. Anim. Breed. Genet., 2006, vol. 125, no. 1, pp. 45—49.
Moravčíková, N., Trakovická, A., and Hazuchová, E., The association of bovine growth hormone gene polymorphism with milk performance traits in Slovak Spotted cows, Sci. Rep. J. Anim. Sci. Biotechnol., 2012, vol. 45, no. 1, pp. 206—210.
Amiri, S., Jemmali, B., Ferchichi, M.A., et al., Assessment of growth hormone gene polymorphism effects on reproductive traits in Holstein dairy cattle in Tunisia, Arch. Anim. Breed., 2018, vol. 61, no. 4, pp. 481—489.
Grochowska, R., Sorensen, P., Zwierzchowski, L., et al., Genetic variation in stimulating GH release and in IGF I of young dairy cattle and their associations with the leucine/valine polymorphism in the GH gene, J. Anim. Sci., 2001, vol. 79, no. 2, pp. 470—476.
Alipanah, M., Kalashnikova, L., and Rodionov, G., Association of prolactin gene variants with milk production traits in Russian Red Pied cattle, Iran. J. Biotech., 2007, vol. 5, no. 3, pp. 158—161.
Dario, C., Carnicella, D., Ciotola, F., Peretti, V., and Bufano, G., Polymorphism of growth hormone GH1-AluI in Jersey cows and its effect on milk yield and composition, Asian-Aust. J. Anim. Sci., 2008, vol. 21, no. 1, pp. 1—5.
Shariflou, M.R., Moran, C., and Nicholas, F.W., Association of the Leu(127) variant of the bovine growth hormone (bGH) gene with increased yield of milk, fat, and protein in Australian Holstein—Friesians, Aust. J. Agric. Res., 2000, vol. 51, pp. 515—522.
Kayumov, F.G., Kosilov, V.I., Gerasimov, N.P. and Bykova, O.A., The effect of SNP polymorphisms in growth hormone gene on weight and linear growth in crossbred red Angus × Kalmyk heifers, Digital Agriculture-Development Strategy (Proc. Int. Sci. Pract. Conf., 2019), Advances in Intelligent Systems Research, pp. 325—328.
Metin, J., Arslan, K., Akyuz, B., Kaliber, M., Aksel, E.G., and Çinar, M.U., Relationships between polymorphisms of growth hormone, leptin and myogenic factor 5 genes with some milk yield traits in Holstein dairy cows, Int. J. Dairy Technol., 2019, vol. 72, no. 1, pp. 1—7.
Bordonaro, S., Tumino, S., Marletta, D., De Angelis, A., Di Paola, F., Avondo, M., and Valenti, B., Effect of GH p. L127V polymorphism and feeding systems on milk production traits and fatty acid composition in Modicana cows, Animals, 2020, vol. 10, no. 9, p. 1651.
El-Komy, S.M., Saleh, A.A., Abdel-Hamid, T.M., and El-Magd, M.A., Association of GHR polymorphisms with milk production in buffaloes, Animals, 2020, vol. 10, no. 7, p. 1203.
Bangar, Y.C. and Magotra, A., Meta-analysis of SNP in growth hormone gene associated with milk traits in dairy cows, Trop. Anim. Health. Prod., 2021, vol. 53, no. 2, pp. 1—7.
Luthfi, M., Ratnawati, D. and Soewandi, B.D.P., Detection of growth hormone (GH| MspI, GHR| AluI, Pit1| HinfI) genes polymorphism and its association with body weight of Grati-Bali Cattle (Bos sondaicus), IOP Conf. Ser., Environ. Earth Sci., 2021, vol. 788, no. 1, p. 012010.
Doosti, A., Arshi, A., and Momeni, B., Molecular study of PIT1 gene polymorphism in Holstein and Iranian native cattle, Afr. J. Agric. Res., 2011, vol. 6, no, 19, pp. 4467—4470.
Pfaffle, R.W., DiMattia, G.E., Parks, J.S., et al., Mutation of the POU-specific domain of Pit-1 and hypopituitarism without pituitary hypoplasia, Science, 1992, vol. 257, no. 5073, pp. 1118—1121.
Aytekin, İ. and Boztepe, S., Associations of PIT1gene polymorphism with milk yield and composition traits in Brown Swiss cattle, J. Anim. Plant. Sci., 2013, vol. 23, no. 5, pp. 1281—1289.
Thuy, N.T.D., Thu, N.T., Cuong, N.H., et al., Polymorphism of PIT-1 and prolactin genes and their effects on milk yield in Holstein Frisian dairy cows bred in Vietnam, Russ. J. Genet., 2018, vol. 54, no. 3, pp. 346—352.
Zabeel, A.K., Al-Bazi, W.G.M., and Muhammed, H.A., Study the association of PIT1 gene polymorphism with milk yield and body weight traits of local breed Iraqi cattle in Kerbala province, Biochem. Cell. Arch., 2018, vol. 18, no. 2, pp. 1867—1871.
Cañizares-Martínez, M.A., Parra-Bracamonte, G.M., Segura-Correa, J.C., and Magaña-Monforte, J.G., Effect of leptin, pituitary transcription factor and luteinizing hormone receptor genes polymorphisms on reproductive traits and milk yield in Holstein cattle, Braz. Arch. Biol. Technol., 2021, vol. 64.
Seyfert, H., Pitra, C., Meyer, L., et al., Molecular characterization of STAT5A and STAT5B-encoding genes reveals extended intragenic sequence homogeneity in cattle and mouse and different degrees of divergent evolution of various domains, J. Mol. Evol., 2000, vol. 50, pp. 550—561.
Liongue, C. and Ward, A.C., Evolution of the JAK-STAT pathway, JAKSTAT, 2013, vol. 2. e22756.
Arun, S.J., Thomson, P.C., Sheehy, P.A., et al., Analysis reveals an important role of JAK-STAT-SOCS genes for milk production traits in Australian dairy cattle, Front. Genet., 2015, vol. 6, no. 342, pp. 1—7.
Khan, M.Z., Khan, A., Xiao, J., et al., Role of the JAK-STAT pathway in bovine mastitis and milk production, Animals., 2020, vol. 10, 11, pp. 1—16.
Xin, P., Xu, X., Deng, C., et al., The role of JAK/STAT signaling pathway and its inhibitors in diseases, Int. Immunopharmacol., 2020, vol. 80, p. 106210.
Tian, Q., Wang, H.R., Wang, M.Z., et al., Lactogenic hormones regulate mammary protein synthesis in bovine mammary epithelial cells via the mTOR and JAK-STAT signal pathways, Anim. Prod. Sci., 2016, 56, pp. 1803—1809.
Meredith, B.K., Kearney, F.J., Finlay, E.K., et al., Genome-wide associations for milk production and somatic cell score in Holstein—Friesian cattle in Ireland, BMC Genet., 2012, vol. 13, no. 21.
Zadworny, D. and Kuhnlein, U., The identification of the kappa-casein genotype in Holstein dairy cattle using the polymerase chain reaction, Theor. Appl. Genet., 1990, vol. 80, no. 5, pp. 631—634.
Peakall, R. and Smouse, P.E., GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research, Mol. Ecol. Notes., 2006, vol. 6, pp. 288—295.
Wang, T., Fernando, R.L., and Kachman, S.D., Matvec User’s Guide, Version 1.03, 2002.
SAS Institute, SAS/STAT.user’s Guide: Release 9.4, SAS Institute Inc., Cary, N.C., 1990.
Citek, J., Rehout, V., and Neubauerova, V., Allele frequency at PRL (prolactin) and LGB (lactoglobulin beta) genes in Red cattle breeds from Central Europe and in other breeds, Czech. J. Anim. Sci., 2001, vol. 46, no. 10, pp. 433—438.
Sorensen, P., Grochowska, R., Holm, L., et al., Polymorphism in the bovine growth hormone gene affects endocrine release in dairy calves, J. Dairy. Sci., 2002, vol. 85, pp. 1887—1893.
Woollard, J., Schmitz, C.B., Freeman, A.E. and Tuggle, C.K., HinfI polymorphism at the bovine Pit-1 locus, J. Anim. Sci., 1994, vol. 72, p. 3267.
Renaville, R., Gengler, N., and Vrech, E., Pit-1 gene polymorphism, milk yield, and conformation traits of Italian Holstein—Friesian bulls, J. Dairy Sci., 1997, vol. 80, no. 12, рр. 3431—3438.
Zwierzchowski, L., Krzyzewski, J., Strzalkowska, N., et al., Effects of polymorphism of growth hormone(GH), pit_1 and leptin (LEP) genes on cow ages lactation stage and somatic cell count on milk yield and composition of Polish Black and White cows, Anim. Sci. Pap. Rep., 2002, vol. 20, no. 4, pp. 213—227.
Ozdemir, M., Topal, M., and Aksakal, V., The relationships between performance traits and the bGH/Alu I and Pit-1/Hinf I polymorphisms in Holstein cows, Indian J. Anim Res., 2017, vol. 52, no. 2, pp. 186—191.
Dario, C. and Selvaggi, M., Study on the STAT5A/AvaI polymorphism in Jersey cows and association with milk production traits, Mol. Biol. Rep., 2011, vol. 38, pp. 5387—5392.
Dybus, A., Grzesiak, W., and Kamieniecki, H., Association of genetic variants of bovine prolactin with milk production traits of Black-and-White and Jersey cattle, Arch. Anim. Breed., 2005, vol. 48, no. 2, pp. 149—156.
Dudule, P.D., Kale, D.S., and Sonwane, A., PCR-RFLP polymorphism at exon 3 and promoter region of prolactin gene in Gaolao Cattle, Indian J. Anim. Res., 2020, vol. 54, no. 5, pp. 523—528.
Ozdemir, M., Sonmez, Z., and Aksakal, V., Associations between PRL/RsaI polymorphism and some performance traits in Holstein cattle reared under organic condition, J. Anim. Plant Sci., 2021, vol. 31, no. 3, pp. 900—905.
Dybus, A., Association of growth hormone (GH) and prolactin (PRL) genes polymorphism with milk production trait in Polish Black-and-White cattle, Anim. Sci. Paper Rep., 2001, vol. 20, pp. 203—212.
Oprządek, J., Flisikowski, K., Zwierzchowski, L., et al., Associations between polymorphism of some candidate genes and growth rates, feed intake and utilisation, slaughter indicators and meet quality in cattle, Arch. Tierz., 2005, vol. 48, pp. 81—87.
Singh, U., Deb, R., Alyethodi, R.R., et al., Molecular markers and their applications in cattle genetic research: a review, Biomark Genome Med., 2014, vol. 6, no. 2, pp. 49—58.
Hartatik, T., Kurniawati, D., and Adiarto, A., Associations between polymorphism of growth hormone gene with milk production, fat and protein content in Friesian Holstein Cattle, J. Indonesian Trop. Anim. Agric., 2015, vol. 40, no. 3, pp. 133—137.
Molee, A., Poompramun, C., and Mernkrathoke, P., Effect of casein genes—beta-LGB, DGAT1, GH, and LHR—on milk production and milk composition traits in crossbred Holsteins, Genet. Mol. Res., 2015, vol. 14, no. 1, рр. 2561—2571.
Çınar, M.U., Akyüz, B., Kıyıcı, J.M., et al., Effects of GH-AluI and MYF5-TaqI polymorphisms on weaning weight and body measurements in Holstein young bulls, Kafkas. Univ. Vet. Fak. Derg., 2018, vol. 24, no. 6, pp. 873—880.
Sönmez, Z., Özdemir, M., Bayram, B., and Aksakal, V., Holstein ineklerde GH/AluI polimorfizmi ile bazı süt verim özellikleri arasındaki ilişkiler, TURJAF, 2018, vol. 6, no. 5, pp. 602—606. https://doi.org/10.24925/turjaf.v6i5.602-606.1838
Dybus, A., Associations between Leu/Val polymorphism of growth hormone gene and milk production traits in Black-and-White cattle, Arch. Tierz. (Dummerstorf), 2002, vol. 45, no. 5, pp. 421–428.
Akcay, A., Daldaban, F., Celik, E., et al., Meta analysis of allele and genotype frequency of growth hormone (bGH) gene AluI polymorphism, which is effective on milk yield in Holstein cattle, Kafkas. Univ. Vet. Fak. Derg., 2020, vol. 26, no. 5, pp. 687—695.
Pozovnikova, M.V., Rotar, L.N., Kudinov, A.A., and Dementieva, N.V., The linkage of polymorphic variants of genes Gh, Prl, and Pit-1 and milk productivity of cows with morphology of cumulus-oocyte complex sampled post mortem, Cytol. Genet., 2020, vol. 54, pp. 212—219.
Dybus, A., Szatkowska, I., Czerniawska-Piątkowska, E., et al., PIT1-HinfI gene polymorphism and its associations with milk production traits in polish Black-and-White cattle, Arch. Anim. Breed., 2004, vol. 47, pp. 557–563.https://doi.org/10.5194/aab-47-557-2004
Garcia, A.B., Angeli, N., Machado, L., et al., Relationships between heat stress and metabolic and milk parameters in dairy cows in southern Brazil, Trop. Anim. Health Prod., 2015, vol. 47, pp. 889—894.
Anggraeni, A., Sumantri, C., Saputra, F., and Praharani, L., Association between GH (g. 1456_1457insT), GHRH (g. 4474 C>A), and Pit-1 (g. 244G>A) polymorphisms and lactation traits in Holstein Friesian cattle, Trop. Anim. Sci. J., 2020, vol. 43, no. 4, pp. 291—299.
Gorbani, A., Prnemat Khanegah, S., Maheri Sis, N., and Taghinejad Rodbaneh, M., Relationship of PIT1 gene polymorphism with dairy traits of Iranian Holstein bulls North West and Karaj breeding centers, J. Anim. Environ., 2020, vol. 12, no. 1, pp. 59—64.
Daniela, E.I. Mazeanschi, A.E., Mhali, C.V., et al., The effect of genetic variations of Pitutary transcription factor 1 (Pit1) on milk performance traits in Romanian cattle, Rev. Rom. Med. Vet., 2021, vol. 31, no. 1, pp. 87—92.
Carșai, T.C., Bâlteanu, V.A., Vlaic, A., and Coșier, V., The polymorphism of pituitary factor 1 (PO U1F1) in cattle, Sci. Pap.: Anim. Sci. Biotech., 2012, vol. 45, no. 1, pp. 142—146.
Bao, B., Chunlei, Zh., Fang, X., Runfeng, Zh., Gu, Ch., Lei, Ch., and Chen, H., Association between polymorphism in STAT5A gene and milk production traits in Chinese Holstein cattle, Anim. Sci. Pap. Rep., 2010, vol. 28, no. 1, pp. 5—11.
Cosier, V., Vlaic, A., and Gaboreanu, I., HindI polymorphism of k-casein and pit 1 genes in Romanian Simmental cattle, Zooteh. Biotehnol., 2006, vol. 40, no. 1, pp. 59—64.
He, X., Chu, M.X., Qiao, L., et al., Polymorphisms of STAT5A gene and their association with milk production traits in Holstein cows, Mol. Biol. Rep., 2012, vol. 39, pp. 2901—2907.
Al-Azzawi, Z.M. and Al-Dulaimi, M.R., Relationship of STAT5a gene (exon 7) polymorphism with milk production traits in Holstein cows in Iraq, Plant Arch., 2020, vol. 20, no. 1, pp. 983—988.
ACKNOWLEDGMENTS
The data and samples were provided by the Animal Breeding Center of Iran.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest. This article does not contain any studies involving animals or human participants performed by any of the authors.
Rights and permissions
About this article

Cite this article
Sadeghi, M., Mokhber, M. & Shahrbabak, M.M. Genetic Variation in Hypothalamic-Pituitary Axis Candidate Genes and Their Effects on Milk Production Traits in Iranian Holstein Cattle. Russ J Genet 58, 1393–1400 (2022). https://doi.org/10.1134/S1022795422110096
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1022795422110096