
Abstract
Early Cretaceous gastropods occur on the eastern margin of Lut Block in Eastern Iran. The gastropod assemblage reported herein is from the Nimbolook section located in the west of Qayen. The section mostly includes marls and limestones. It was dated as Aptian owing to the findings of foraminifers, nannofossils and ammonites. The exposure provides a relatively well-preserved gastropod fauna from marls and limestones. New species Oligoptyxis khorasanica sp. nov. from the family Nerineidae is the first find of the genus in the Cretaceous of Iran. Finally, the new genus Purpuripullina, gen. nov. was proposed. It was previously included in the family Ampullinidae.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The knowledge on Early Cretaceous marine gastropod faunas from Iran is still very incomplete. Despite reporting some fossil groups, particularly from the Qayen area, gastropod associations from the Eastern Iran marine basins are still poorly documented. Detailed geological studies of the area were performed by Fauvelet and Eftekharnezhad (1990), Berthiaux et al. (1981), Raisossadat et al. (2011, 2014, 2015, 2016) in which they published the aspects of local Cretaceous stratigraphy. Raisossadat and Noori (2016) reported gastropods from the area. The rudists were studied by Raissosadat and Skelton (2005) and Khazaei et al. (2011). Foraminifera and Orbitolina limestones were studied by Babazadeh et al. (2010), Motamedalshariati et al. (2016), Raisossadat et al. (2017), Zarei et al. (2014) and Asadi (2013). Latifi et al. (2018) studied the nannoplankton, and Sharifi et al. (2016) reported ammonites.
However, many aspects of research still have to be undertaken. For instance, although the foraminifera, nannofossils, ammonites were studied more than other faunas from Cretaceous beds in the east of Iran; it is necessary to place more emphasis on Gastropoda to improve our knowledge about systematic paleontology and the assemblage might be used for correlation with other areas of Tethys.
This study establishes Aptian record of gastropods from eastern Iran. The present paper describes a species from Family Acteonellidae, a new genus from Family Ampullinidae, and new species from Family Nerineidae. All collected samples are kept in the Geology Department of Birjand University.
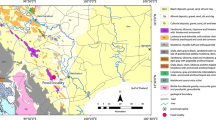
One section from eastern Iran in the area of Qayen was measured and investigated. Qayen area is located in the east of Iran, and the Nimbolook section is measured and studied in this region (Fig. 1). The section is located near Qayen–Gonabad main road and lies at 59°00′49′′ E and 33°52′17′′ N.

Location map of the Nimbolook section.
MATERIALS AND METHODS
In this section, the succession was measured bed-by-bed, the specimens were collected from the surface, and lithology microfauna, and benthic macrofauna were recorded. Samples were collected from marls and limestones at intervals in the case of the Early Cretaceous of Nimbolook section.
Laboratory investigations included cleaning, washing and photographing the macrofaunal elements.
A few specimens are preserved as internal moulds. Some of them occur in shell preservation. Internal moulds show fewer features, hence precise identification was not possible. Specimens were whitened with magnesium oxide dust which accentuates finer ornamentations. Besides, specimens’ axial cross-sections were produced to investigate internal structures such as internal plaits and columellar.
STRATIGRAPHY
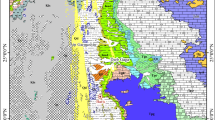
Nimbolook section is situated at the 1 : 100 000 map of Qayen (Berthiaux et al., 1981) (Fig. 2). From the tectonic point of view, this region is one of the most parts in the northern Sistan suture zone (Tirrul et al., 1983) between Afghan and the Lut blocks. Post Neocomian sediments are also deposited in Nimbolook region.

Geological map of the Nimbolook area (redrawn after Berthiaux et al., 1981).
In the Nimbolook section, post Neocomian sediments can be divided into two sedimentary sequences, the first sedimentary cycle is transgressive to Cretaceous sea at the highest part of Early Cretaceous, although occurring late, is detectable in most parts of region. The first transgressive cycle starting from Aptian was determined (Berthiaux et al., 1981), and in the Nimbolook section, it starts with \({\text{K}}_{{\text{1}}}^{{{\text{cl}}}}\) clastic rock unit which includes sandstone and red conglomerate deposited after the Neocomian (Fig. 2). The sedimentation continues to \({\text{K}}_{{\text{1}}}^{{{\text{cs}}}}\) containing conglomerate, sandstone, marl and limestone which was studied on this unit. In the Nimbolook section, the time separation between the highest part of the Early Cretaceous and the lowest part of the Late Cretaceous is carried out well. The second sedimentary cycle started at the beginning of Late Cretaceous age in this area, again started with clastic units (red conglomerate) indicated in the map with \({\text{K}}_{{\text{2}}}^{{{\text{cl}}}}\). Hence, there are segregated collections of limestone, marl, sandstone, and conglomerate which are indicated with \({\text{K}}_{{\text{2}}}^{{{\text{fl}}}}\).
The lower boundary of this section is laid on a red conglomerate and sandstone sequence attributed to the Early Cretaceous. The section starts with sandstone, conglomerate, and gray shale (unit 1) (Figs. 3, 4). Then, it continues with gray marl with macrofossils (unit 2) possessing Oligoptyxis khorasanica sp. nov. (samples Qn2, 3, 4, 5, 6) which is recorded from this unit and unit 3. Acteonella sp. (sample Qn5) is recorded from this unit. In the upper part of this section (unit 3), medium to thick bedded, the cream to gray limestone is appeared which contains gastropods, rudists, and other pelecypods, corals, benthic and orbitolinid foraminifera. O. khorasanica (sample Qn6) and Purpuripullina sp. (sample Qn7) are recorded from this unit, and the identified benthic foraminifera species indicate the late Aptian age for this section. The thickness of this section is 55 meters. Based on identified benthic foraminifera an early–late Aptain is suggested. Moreover, the succession above the studied sequence was dated as late Aptian–Albian–Cenomanian using foraminifera (Motamedalshariati et al., 2016), nanofossils (Latifi et al., 2018) and ammonites (Sharifi et al., 2016). Therefore, based on above age determination for the studied sequence, an early–late Aptian age is confirmed.

View of Nimbolook section.

Lithostratigraphic column of the Nimbolook section.
SYSTEMATIC PALEONTOLOGY
The gastropoda samples are not mostly preserved and complete specimens; we use samples that are acceptable for biometric measurements of various parts of shells. To describe gastropods we use terminology provided by Cox (1964). All measurements are linear (using a Vernier caliper) in millimeters and are presented in the tables. Abbreviations and parameters include: Height (H), Diameter (D), Height of last whorl (HL), Height of aperture (HA), Width of aperture (WA), number of whorl (nw), Spire angle (in degrees), (Sa°), the ratio of shell diameter to its height (D/H) and the ratio of the height of the body whorl (the last whorl) to height of the shell (HL/H) (Fig. 5).

Measurements of gastropod shell.
FAMILY ACTEONELLIDAE GILL, 1871
Genus Acteonella d’Orbigny, 1843
Acteonella sp.
Plate 1, figs. 5–8

Cross-sections of Oligoptyxis khorasanica sp. nov.: (a) holotype, University of Birjand, no. 100-GBU-Qn5-5, (b) specimen, number Qn4-1.
Description. Shell involute, ovaly elongated with conically narrowed apical part. Exact axial cross section shows that apical part has a slightly concave shape (Plate 1, fig. 5). Outer surface of collected specimens is strongly eroded and don’t show the real shell form. Whorl section has a lunate section, with maximal breadth anteriorly and gradually narrowing upwards. Three folds are formed anteriorly on columellar side.

Plate 1 . (fig. 1) Purpuripullina sp., specimen no. Qn7-1, shell height 30 mm; (figs. 2–4) Oligoptyxis khorasanica sp. nov.: (2) specimen no. Qn2-1, shell height 31 mm; (3) specimen no. Qn4-1, shell height 24 mm; (4) holotype no. Qn5-5, shell height 43 mm. (figs. 5–8) Acteonella sp.: (5) specimen no. Qn5-1, shell height 30 mm; (6) specimen no. Qn5-4, shell height 40 mm; (7) specimen no. Qn5-3, shell height 39 mm; (8) specimen no. Qn5-2, shell height 47 mm.
Discussion. Although our specimens don’t demonstrate the character of umblicus from recrystallization, the shell form is common for species of Acteonella (Sogdianella) with rudimentary umbilical pockets visible in their cross sections. For example, A. (S.) suрernata Pčelincev, 1953 (Cenomanian) (Pčelincev, 1953, pl. 44, fig. 7; Hacobjan, 1976, pl. 64, fig. 4) is a typical species of subgenus Sogdianella. Our form differs from A. (S.) supernata in concave and higher apical part. The same apical part has A. ornata Pčelincev, 1953 from the Cenomanian of Transcaucasia. They also have the same form in young shell.
Shells from Nimbolook section present a new type of acteonellas that were already unknown from the Lower Cretaceous outside of Europe. Before this, two types of Acteonella shell were described. Firstly, it is a high spired acteonellas: A. delgadoi Choffat, 1901 from the upper Albian of Portugal (Choffat, 1901, p. 109), A. gracilis H. Douvillé, 1916 from the upper Albian of Egypt (Douvillé, 1916, p. 147), Acteonella sp. (Delpey, 1940, p. 231) from the Albian of Lebanon, and A. baconica Benkö-Czabalay, 1962 from the Aptian of Hungary (Benkö-Czabalay, 1962, pp. 221, 270, pl. 7, figs. 8–11 only: different species under one name). Such forms were proposed to be included in separate subgenus Pchelincevella Dzhalilov, 1972. Secondly, it is Acteonella described from the lower Aptian of neighboring section near Hajiabad, Iran (Raisossadat and Noori, 2016, p. 298, as Sogdianella sp.). It has less slender shell than in Nimbolook’s Acteonella, with obtusely conical apical part. Acteonella from Nimbolook and A. baconica (Benkö-Czabalay, 1962, pl. 7, figs. 6, 7, 12–16, including holotype) makes the third type, occupying an intermediate position in morphology between the two first types. So Acteonella was sufficiently diverse group even before the Late Cretaceous.
Measurements:
Number | H | D/H |
Qn5-1 (figs. 5a, 5b, 5c) | 30 | 0.57 |
Qn5-2 (figs. 8a, 8b, 8c ) | 47 | 0.51 |
Qn5-3 (figs. 7a, 7b, 7c ) | 39 | 0.46 |
Qn5-4 (figs. 6a, 6b) | 40 | 0.52 |
Material. Four specimens from sample Qn5.
?FAMILY AMPULLINIDAE COSSMANN, 1919
Genus Purpuripullina Guzhov and Raisossadat, gen. nov.
Name origin: named using combination of generic names Purpuroidea and Ampullina.
Type species: Natica? scalaris Conrad, 1852 from the Upper Cretaceous of Lebanon.
Diagnosis. Large and enough high spired strongly gradate anomphalous shells with keel-like shoulder at the whorl top and commonly with rounded basal-palatal bent between whorl side and base. Upper bent straight or nodose. The rest whorl surface covered by more or less prominent growth lines and sometimes also by round plicae, beginning from shoulder. Whorl side is more or less concave between shoulder and bent. Whorl base convex is without sculpture. Whorls thick-walled is oval in inner cross section. Growth lines are slightly prosocline having rounded aperture below.
Composition. Type species, Chemnitzia aptiensis Lànderer, 1872 (Aptian), Coronatica purpuroidea Blanckenhorn, 1927 (Cenomanian).
Comparison. Purpuripullina differs from other genera of family in strongly gradate shells with concave whorl side.
Discussion: Generic origin is unclear. Most probably it is derived from its ancestor with rounded whorls, so keel-like bent was appeared before than basal-palatal bent. Purpuripullina is more like (excluding sculptural features) the thick-walled and large-sized representatives of Ampullinidae from simultaneous deposits, which is placed in family with some degree of conventionality.
Distribution. Lower Cretaceous, Aptian–lower Upper Cretaceous of Southern Europe and the Middle East.
?Purpiripullina sp. ind.
Plate 1, fig. 1
Description. Sole bed preserved specimen with strongly reworked shell surface. But the presence of prominent bent near whorl top, concavity of whorl side and poorly preserved and weakly visible basal-palatal bent is visible. Axial shell section shows thick shell wall, thick solid columella and inner oval whorl sections. This specimen is most close to Purpuripullina in mentioned preserved features.
Measurements:
Number | H | D | HL | HA | WA | Sa° | nw | D/H | HL/H |
Qn7-1 (figs. 1a, 1b) | 80 | 53 | 50 | 19 | 14 | 80 | 4 | 0.66 | 0.62 |
Material. One specimen from sample Qn7.
FAMILY NERINEIDAE ZITTEL, 1873
SUBFAMILY DIPTYXIDINAE PČELINCEV, 1965
Genus Oligoptyxis Pčelincev, 1953
Type species. Oligoptyxis turricula Pčelincev, 1953 from Cenomanian–lower Turonian, Armenia.
Description. High-spired shell with low, more or less convex whorls. Periphery of whorls is rounded. Interior of whorls has moderately concave columellar portion. Very small parietal plait is in some whorls.
Discussion. The Diptyxidinae, consisting of the genera Diptyxis, Cylindroptyxis and Oligoptyxis, was erected by Pčelintsev (1965). The primary two genera are well-characterized, the main difference between them is in the overall shell shape (Cylindroptyxis shells are semicylindrical while Diptyxis shells are described as conical), and the solid columella of Diptyxis is compared to a narrow closed umbilicus in Cylindroptyxis (Vaughan, 1988). Phaneroptyxis (Family Itieriidae Cossmann, 1896) shows similarity in morphology, but has always concave whorls; in contrast, in Oligoptyxis whorls are concave, flat or convex. In the preserved specimens a long narrow siphonal channel is indicated by the growth lines. The other important morphological feature of Oligoptyxis is the small parietal plait, which does not occur in Phaneroptyxis.
Genus Oligoptyxis is widely spread in Transcaucasia, Middle Asia (mostly in Tadzhikistan), the southern Soviet Union (Hacobjan, 1976). Moreover, the genus is known from the Austria (Kollmann, 1976, pl. 4, figs. 29–37). The stratigraphic distribution is the late Albian to Turonian. Hacobjan (1976) included O. velesensis Hacobjan, 1976 and O. macedonica Hacobjan, 1976 in this genus. It is also proposed for shells that determined by Ćirić (1952) as Cerithium rotulare Stoliczka, 1863 and C. sturi Stoliczka, 1863 respectively from the Upper Cretaceous of the Republic of Macedonia. But in our opinion the Macedonian material differs from weakly plicated Oligoptyxis by entirely absenting signs of plication and doesn’t have the same oblong rhomboid whorl sections (see also shells defined as C. sturi in Stefanoff (1931, pl. 6), Tzankov and Motekova (1981, pl. 14, figs. 7–10). It is most probably poor preserved non-nerineid taxa. Also several poorly preserved shells were assigned to Oligoptyxis by Rossi Ronchetti (1967) from the Upper Cretaceous of Pakistan. Her O. aralensis is not nerineid gastropod, so as O. cylindrica and O. turricula have so terrible preservation that it is impossible to give a generic identification. Rossi Ronchetti (1967) stated in description that she couldn’t recognize plication in those specimens.
Oligoptyxis khorasanica is the first report of the genus in territory of Iran and the first finding of genus in the Lower Cretaceous.
Oligoptyxis khorasanica Guzhov and Nazemi, sp. nov.
Plate 1, figs. 2–4
Name origin: after historical province Khorasan.
Holotype. University of Birjand, no. 100-GBU-Qn5, Iran, Southern Khorasan, near village of Nimbolook: Aptian. Figured: pl. 1, fig. 4.
Diagnosis (Fig. 6). Enough large highly turriculate anomphalous shells, with height more than 10 cm, up to 13 cm, teleoconch angle 8°–15°. Whorls are concave, forming sutural thickened ridge, with spiral row of tubercles in the middle of whorl side. Whorl section rhombic has siphonal projection anteriorly. Inner whorl section with medial well developed parietal fold, which are broad enough and with rudimentary columellar fold is formed anteriorly. Basal side of inner whorl section is straight or sometimes has low elevation.
Comparison. O. khorasanica differs from O. glabra Pčelincev, 1953 in larger size, concave sculptured later whorls, less developed columellar fold; from O. turricula Pčelincev, 1953 in sculptured and strongly concave adult whorls, better developed parietal fold. It differs from the other species in well-developed parietal fold and concave sculptured whorls.
Measurements:
Number | H | D | HL | HA | HW | Sa° | nw | D/H | HL/H |
Qn2-1 (figs. 2a, 2b) | 31 | 11 | 9 | 6 | 5 | 15 | 12 | 0.35 | 0.29 |
Qn4-1 (figs. 3a, 3b) | 24 | 12 | 8 | 6 | 6 | 8 | 5 | 0.5 | 0.33 |
100-GBU-Qn5-5 (Figs. 4a, 4b) | 43 | 13 | 10 | 6 | 5 | 11 | 11 | 0.30 | 0.23 |
Material. Three specimens from samples Qn2, Qn4, Qn5.
CONCLUSIONS
Nimbolook section with 55 meters thickness was studied in the east of Iran. The lithology at the base consists of conglomerates and sandstone. The lithology presents shale and sandstone intercalations, marls and limestones. Acteonella sp., Oligoptyxis khorasanica sp. nov. and Purpuripullina, gen. nov. are reported from this section. Based on benthic foraminifera such as orbitolinids Aptian attributed to this section. Nerineoidea and Acteonellidae are typical for Tethys.
REFERENCES
Asadi, A., Biostratigraphy and paleoenvironment of Lower Cretaceous units based on Orbitolina of Nimbolook Section (Northwest of Qayen), East of Iran, MSc Thesis, Payam Noor University of Birjand, 2013.
Babazadeh, S.A., Raisossadat, S.N., and Ahrari, F., Biostratigraphy and evolutionary trend of the Cretaceous orbitolinids in the sedimentary succession of east Lut Block, south west of Qayen, Sedimentary Facies, 2010, vol. 3, no. 1, pp. 1–10.
Benkö-Czabalay, L. and Bakonyhegység, A., Apti, Albai és Cenomán gastropodái, Geol. Hungarica, Ser. Palaeontol., 1962, vol. 31, pp. 181–291.
Berthiaux, A., Christmann, P., Fauvelet, E., Hatrival, J.N., Maurizot, P., and Vaslet, D., Geological Map of the Qayen, Scale 1/100 000, Tehran: Geol. Surv. Iran, 1981.
Choffat, P., Recueil d'études paléontologiques sur la faune Crétacique du Portugal, Espèces diverses, Sect. Trav. Geol. Portugal, Sér. 4, 1901, pp. 107–171.
Ćirić, M.B., Faune Crétacée des environs de Titov Veles, Bull. Mus. Hist. Nat. Pays Serbie, Ser. A, 1952, no. 5, pp. 249–276.
Cox, L.R., General characteristics of gastropods, in Treatise on Invertebrate Paleontology, Pt. I. Mollusca 1, Kansas: Univ. Kansas Press, 1964, pp. 169–331.
Delpey, G., Les gastéropodes Mésozoïques de la region Libanaise, Notes et Mémoires. Haut-Commissariat de la République Française en Syrie et au Liban. Service des Travaux Publics-Section d'Études Géologiques, 1940, vol. 3, pp. 5–305.
Douvillé, H., Les terrains secondaires dans le Massif du Moghara à l’Est de l’Isthme de Suez, d’après les explorations de M. Couyat-Barthoux. Paléontologie, Mém. Acad. Sci. Inst. France, 1916, vol. 54, no. 1, pp. 1–184.
Dzhalilov, M.R., To systematics of Acteonellidae (Gastropoda), Paleontol. Zh., 1972, no. 1, pp. 16–23 [in Russian].
Fauvelet, E. and Eftekharnezhad, J., Explanatory Text of the Qayen, Quadrangle Map 1: 250 000 Scales, Tehran: Geol. Surv. Iran, 1990.
Hacobjan, V.T., Pozdnemolovye gastropody Armyanskoi SSR (Late Cretaceous Gastropods of the Armenian SSR), Erevan: Akad. Nauk Armyanskoi SSR, 1976 [in Russian].
Khazaei, A., Raisossadat, S.N., and Asadi, Sh., Rudist bivalves (Requieniidae family) in Early Cretaceous sediments from SW Qayen, Eastern Iran, paleobiogeographic aspects, Sedimentary Facies, 2011, vol. 3, no. 2, pp. 52–67.
Kollmann, H.A., Gastropoden aus den Losensteiner Schichten der Umgebung von Losenstein (Oberösterreich) 1. Teil: Euthyneura und Prosobranchia 1, Ann. Naturhist. Mus. Wien, 1976, vol. 80, pp. 162–206.
Latifi, Z., Foroughi, F., Motamedalshariati, M., and Raisossadat, S.N., Calcareous nannofossils biostratigraphy of Lower Cretaceous deposits at the east of Iran, NW of Qayen (Nimbolook stratigraphic section), Sci. Quart. J., Geosci. (Geol. Surv. Iran), 2018, vol. 27, no. 106, pp. 41–50.
Motamedalshariati, M., Raisossadat, S.N., Moloudi, D., and Mortazavi, M., Foraminifera biozonation and morphogroups from Nimbolook section, east margin of Lut block, Iran, Arabian J. Geosci., 2016, vol. 9, no. 19. Iss. 720, pp. 1–11. https://doi.org/10.1007/s12517-016-2709-y
Pčelincev, V.F., Fauna bryukhonogikh verchnemelovykh otlozenii Zakavkaz’ya i Srednei Azii (Gastropods of the Upper Cretaceous deposits of Transcaucasia and the Middle Asia), Leningrad: Izd. Akad. Nauk SSSR, 1953 [in Russian].
Pčelincev, V.F., Murchisoniata mezozoya Gornogo Kryma (Mesozoic Murchisoniata of the Crimean Mountains), Moscow, Leningrad: Nauka, 1965 [In Russian].
Raisossadat, S.N. and Noori, H., Lower Cretaceous gastropods from the Qayen area, Eastern Iran, Geobios, 2016, vol. 49, no. 4, pp. 293–301.
Raisossadat, S.N. and Skelton, P.W., First record of rudist fauna from the Qayen area, Eastern Iran, in Proc. 7th Int. Cretaceous Symp., Neuchatel, Switzerland, 2005, pp. 177–178.
Raisossadat, S.N., Mosavinia, A., Khazaei, A.R., and Asadi, Sh., Stratigraphy of Cretaceous deposits in southwest of Qayen area (East of Iran), in Proc. 5th Symp. Iranian Paleontol. Soc., Mahan, Kerman, 2011, pp. 216–221.
Raisossadat, S.N., Asadi, Sh., Zarei, H., Khazaei, A.R., Mortazavi, M., Motamedalshariati, M., Mosavinia, A., Moloudi, D., and Yazdi Moghadam, M., Stratigraphy, paleoecology and paleobiogeography of Cretaceous deposits in the east of Lut Block (Qayen area), in Proc. 18th Symp. Geol. Soc. Iran, Tarbiat Modarres Univ., December 24–25, 2014, pp. 600–610.
Raisossadat, S.N., Wang, P.L., Mortazavi Mehrizi, M., Motamedalshariati, M., and Sharifi, J., Biostratigraphy and chemostratigraphy of Upper Albian–Lower Cenomanian deposits in southwest Qayen area of eastern Iran, Berichte des Institutes für Erdwissenschaften der Karl-Franzens-Universität Graz, 2015, vol. 21.
Raisossadat, S.N., Zarei, H., Mortazavi Mehrizi, M., and Yazdi Moghadam, M., Urgonian facies in south west Qayen, in Proc. 10th Symp. Iranian Paleontol. Soc., 24 April, Farhangian Univ., Nyshabour, Iran, 2016, pp. 69–73.
Raisossadat, S.N., Motamedalshariati, M., and Erfani, M., Foraminifera biostratigraphy of Albian–Cenomanian deposits in southwest of Qayen, East of Iran, Berichte Geol. Bundesanst., 2017, vol. 120, p. 221.
Ronchetti, C., Mollusks from the Upper Cretaceous at Burji La (Baltistan, Central Asia), Riv. Ital. Paleontol. Stratigr., 1967, vol. 73, no. 3, pp. 811–832.
Sharifi, J., Raisossadat, S.N., Mortazavi, M., and Motamedalshariati, M., Albian and Cenomanian ammonites of the eastern margin of the Lut block (East Iran), Carnets Geol., 2016, vol. 16, no. 25, pp. 591–613.
Stefanoff, A., Le Crétacé supérieur dans les versants occidentaux de la montagne Golo-Bǎrdo en Bulgarie (études stratigraphiques et paléontologiques), Z. Bulgar. Geol. Ges., 1931, vol. 3, no. 2, pp. 3–32.
Tirrul, R., Bell, I.R., Griffis, R.J., and Camp, V.E., The Sistan suture zone of eastern Iran, Geol. Soc. Am. Bull., 1983, vol. 94, no. 1, pp. 134–150.
Tzankov, V. and Motekova, N., Gastropoda, in Les Fossiles de Bulgarie. V. Crétaceé Supérieur. Grandes Foraminifères, Anthozoaires, Gastéropodes, Bivalvia, Sofia: Imprimerie de l’Académie Bulgare des Sciences, 1981, pp. 52–72.
Vaughan, P.G., Cretaceous Nerineacean Gastropods: Systematic, Affinities and Palaeoecology, PhD Thesis, London: Open Univ., 1988.
Zarei, H., Raisossadat, S.N., Yazdi Moghadam, M., and Mortazavi Mehrizi, M., Lower Cretaceous paleoecology in the Qomenjan section, southwest Qayen, based on benthic Foraminifera, in Proc. 8th Symp. Iranian Paleontol. Soc., May 21–22, 2014, Zanjan Univ., Zanjan, Iran, pp. 184–188.
ACKNOWLEDGMENTS
The authors are grateful to Prof. S.V. Popov (Paleontological Institute, Russian Academy of Sciences) and Prof. V.A. Zakharov (Geological Institute, Russian Academy of Sciences) for their valuable comments and suggestions. Logistical support for field works and laboratorial facilities was partly financed by University of Birjand (Iran) and is appreciated. Thanks go to MSc students of University of Birjand, Mr. Kamerani and Miss Afrasiabian for their help in the field.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Reviewers S.V. Popov and V.A. Zakharov
Rights and permissions
About this article

Cite this article
Seyed Naser Raisossadat, Guzhov, A., Arzhaneh, R.N. et al. Gastropoda from Lower Cretaceous Deposits of Nimbolook Area, Eastern Iran. Stratigr. Geol. Correl. 30, 65–74 (2022). https://doi.org/10.1134/S0869593822010063
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0869593822010063