
Abstract
Based on the results of research in 1995–2020, mass and potentially commercial fish species have been identified in Russian waters of the Chukchi Sea. Among the 72 species recorded in trawl catches, 16 species from six families were mass species; fishes of the cod family (Gadidae) were absolutely dominant among them (on average, 92.6% of the total commercial biomass of marine fishes). In the late 1990s–early 2000s, the stocks of this family were represented mainly by polar cod Boreogadus saida. In the late 2010s, its biomass significantly decreased; however, the stocks of Walleye pollock (Gadus chalcogrammus) significantly increased as a result of its migrations from the northern part of the Bering Sea. The size-age and weight characteristics, features of the linear and weight growth, and period, scale, and conditions of spawning and feeding have been analyzed for Walleye pollock, polar cod, saffron cod Eleginus gracilis, and Pacific cod G. macrocephalus. The biological parameters of the fishes from the Chukchi and Bering seas have been compared. It is noted that saffron cod and Pacific cod in the study area are represented mainly by juveniles and immature individuals. In addition to juveniles, a significant number of individuals with a length of over 50 cm at age ≥7 years have been recorded among Walleye pollock individuals in 2018–2020. Polar cod from the Chukchi shelf differed in a smaller body size than polar cod from the northwestern part of the Bering Sea.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
The Russian Arctic with its extremely harsh natural conditions is a strategically important region, primarily because it covers up to a quarter of the world’s undiscovered oil and gas reserves on the Arctic shelf (Kovalev and Filippova, 2010). Renewable biological resources also play an important role in this region. It is significant stocks of marine fishes in the western sector of the Arctic (mainly in the Barents Sea and, to a lesser extent, in the White, Kara, and Laptev Seas) that make it possible to carry out long-term stable fishing here. For instance, it was recommended to catch 56.84 thousand tons of 45 fish stock units in the waters of these seas in 2020, while fishing opportunities in the eastern sector of the Arctic (the East Siberian and Chukchi Seas) were limited to 11 fishing objects with a total volume of 3.91 thousand tons, of which 3.9 thousand tons were represented by one species, polar cod Boreogadus saida, from the Chukchi Sea.
The revival of state interest in the Arctic seas has led to an increase in the number of expeditionary studies, primarily in the Chukchi Sea, the fishing significance of which has so far been poorly studied. Whereas five integrated trawl surveys were performed in Russian waters of the sea from 1990 to 2009, the same number of trawl surveys have been performed in the last decade (three of them in 2018–2020). An important result of the regularity of studies was the discovery of commercial aggregations of Walleye pollock Gadus chalcogrammus in the southwestern part of the Chukchi Sea, which made it possible to recommend 37.2 thousand tons of this species for catching in the total allowable catch regime in 2021Footnote 1.
The appearance of significant stocks of large-sized Walleye pollock in 2018 and, in particular, in 2019–2020 in Russian waters of the Chukchi Sea, which was previously hardly recorded during trawl studies (Orlov et al., 2020), resulted from climatic and oceanological changes in the Bering and Chukchi seas in recent years (Baker et al., 2020; Basyuk and Zuenko, 2020; Danielson and al., 2020; Plotnikov et al., 2020; Siddon et al., 2020). The general warming of the air and sea waters, as well as a decrease in the area of sea ice and the reduction of its melting period and changes in the circulation of currents and composition of food supply, have contributed to the redistribution of part of pollock aggregations to the northern part of the Bering Sea, followed by their penetration into the southwestern part of the Chukchi Sea through the Bering Strait (Eisner et al., 2020; Nishio et al., 2020; Baker, 2021; Orlov et al., 2021). The intensification of the migration activity in the northern direction within the Bering Sea in the late 2010s was also recorded for Pacific cod Gadus macrocephalus, Pacific herring Clupea pallasii, pink salmon Oncorhynchus gorbuscha, capelin Mallotus villosus catervarius, and juvenile sockeye salmon O. nerka (Eisner et al., 2020; Farley et al., 2020; Yasumiishi et al., 2020; Datsky and Samoilenko, 2021). On the whole, climate change in the last decade has led to significant transformations in marine ecosystems, including fish communities, which has been significantly reflected in the distribution of fish resources, biomass, and biology.
The uniqueness of Russian waters of the Chukchi Sea is that its water masses are strongly influenced by warm and saline transformed Pacific waters entering from the Bering Sea through the Bering Strait and, at the same time, by the cold waters of the Arctic Ocean and freshened waters of the East Siberian Sea entering through the Long Strait (Dumanskaya, 2017). It seems quite probable that many representatives of fish communities in the Chukchi Sea may respond to the variability of the habitat and serve as a kind of indicator of possible transformations in the ecosystem of this water body and the adjacent waters. At the same time, there is no up-to-date information on the features of habitat, distribution, and biology (size–age composition, growth rate, age, maturity duration, and spawning period and scale) even for mass fish species living in the water area of the sea.
The purpose of this research is to identify mass and potentially commercial fish species in Russian waters of the Chukchi Sea, compare their biological features with the biological features of fishes from the adjacent waters of the Bering Sea. This publication (Part 1) presents long-term data on the commercial biomass of marine fishes and biology of representatives of the cod family (Gadidae), which dominates in the studied area. In our further research (parts 2 and 3), we will consider similar data on fish species from the flatfish (Pleuronectidae), bullhead (Cottidae), herring (Clupeidae), smelt (Osmeridae), and salmonid (Salmonidae) families and make the final conclusion.
MATERIALS AND METHODS
In this study, we used expeditionary data on the commercial biomass and biology of mass fish species from Russian waters of the Chukchi Sea. Materials of nine integrated trawl surveys (which included 366 trawlings) performed in 1995–2020 on research vessels of the Research Fleet Base of the Russian Federal Research Institute of Fisheries and Oceanography served as a basis for this study. On the whole, our studies in the summer–autumn period covered a significant area of the Russian sector of this Arctic sea, except the coastal shallow part with depths of less than 26 m (Table 1). The resulting data on the biology of fishes from the Chukchi shelf were compared with the previously presented (Datsky and Andronov, 2007; Datsky, 2015b; Datsky, 2016) materials of trawl surveys from the northwestern part of the Bering Sea. The schematic map of Russian waters of the Chukchi Sea and adjacent areas of the Bering Sea, which were covered by the comparison analysis, is shown in Fig. 1. The coordinates of the boundary of the Chukchi Sea fishing zone in the figure correspond to those indicated in the Order of the Ministry of AgricultureFootnote 2.

Schematic map of Russian waters of the Chukchi Sea and separate water areas in the northwestern part of the Bering Sea: (⚫) scientific trawling sites in 1995–2020, (– –) boundary of the Chukchi Sea fishing zone, (–·–) state maritime border of Russia. Designations of the areas: CS, Chukchi Sea, CB, Chirikov basin; NA, CA, and WA, the northern, central, and western parts of the Gulf of Anadyr, respectively; KA, Koryak area.
Unlike the Bering Sea, where the biological characteristics of fishes were recorded using various fishing gear throughout almost the entire calendar year (Datsky and Andronov, 2007; Datsky, 2016, 2017a, 2017b; Datsky and Maznikova, 2017), material in the Chukchi Sea was collected in a limited time (August–September) exclusively with trawls due to harsh natural conditions and the absence of fishing activity. During integrated surveys, we used bottom and pelagic trawls with a fine mesh (10 mm) in the cod end for effective catching of juvenile fish; the duration of each trawling was 30 min. Trawl surveys were pelagic in 2003, 2007, and 2008 and bottom in the other years. To clarify the prospects for Walleye pollock fishing in the southern part of the sea, five trawlings were also carried out with the above-mentioned pelagic trawl (but without a fine-meshed insert) in early September 2020, each trawling with a duration of 3 hours.
Using data from cruise reports, in which the total fish biomass was determined by the area method (Aksyutina, 1968) or modified Voronoi–Dirichlet method of mesh strata (Borisovets et al., 2003) based on the average catch value per 1 km2 of the studied water area with differentiated trawl catchability coefficients (Macrofauna…, 2012), we identified mass and potentially commercial fish species. For this purpose, we calculated their commercial biomass, i.e., part of the total stocks of hydrobionts consisting mainly of mature individuals that reached commercial size and, accordingly, were of interest within the organization of industrial and coastal fisheries. It should be noted that, since there is insufficient information on the mass maturation of fish upon reaching a certain body length and age and there are no appropriate biological justifications for determining the minimum commercial size of fish resources within the Chukchi Sea, our analysis involved the available materials on other water areas of the Far Eastern fishery basinFootnote 3. Therefore, in addition to data on mass species in catches (which, however, are often represented by noncommercial individuals), new data on potentially commercial fish were also obtained. It should be noted that fish with an occurrence of more than 50% in trawl catches were considered mass species (Sheiko and Fedorov, 2000) (if they met this criterion based on the results of studies for at least one year); species that are currently not used by fishery, but can be involved in fishery under certain conditions without prejudice to their reproduction were considered potentially commercial.
The fish biology was described using data from 53 945 thousand measurements of body length, 7367 thousand biological analyzes (complete or only with dissection), and 1343 thousand age determinations. For size–weight characteristics, the fork length (FL) of all fish individuals was measured with an accuracy of 1 cm and the total body weight was determined. Complete biological analysis was performed not only for modal size groups but also for large and small individuals of both sexes. For this purpose, five to ten male specimens and the same number of females were purposefully selected per 1 cm of the size range. During most of the cruises, representatives of ichthyofauna were weighted on a Marel electronic balance (Marel, Iceland) designed for operation under sea rolling conditions, at an error of 0.1 g. The sex ratio, size–age composition, and distribution of fish by gonad maturity stages are represented using the arithmetic means of relative values by year.
The age was determined by otoliths of 1343 fish individuals (polar cod, Walleye pollock, Bering flounder Hippoglossoides robustus, and Pacific cod: 684, 478, 177, and 4 specimens, respectively). The age of Bering flounder was determined under laboratory conditions by whole otoliths (regardless of their size) using an Olympus SZX 12 stereomicroscope (Olympus, Japan) with Cell image analysis against the otolith transmitted light (Lear and Ritt, 1975; Bowering and Nedreaas, 2001; ICES, 2017). For large otoliths of flounders and all cod otoliths, we used the “break and burn” method, which is widely used in studies of both cod fishes and flounders (Chilton and Beamish, 1982; Brodziak and Mikus, 2000; Buslov and Varkentin, 2001; Zolotov, 2006; Fey and Weslawski, 2017). According to this technique, the otolith was broken in half and burned in the flame of an alcohol lamp and then viewed in reflected light to count annual rings as a combination of light and dark zones.
RESULTS AND DISCUSSION
Commercial Biomass of Mass Fishes in Russian Waters of the Chukchi Sea
Previous studies (Datsky, 2015a) showed that only 42 species of the ichthyofauna of Russian waters of the Chukchi Sea (38.2% of the recorded fish species) constantly occurred in fishing gear. Materials supplemented with the up-to-date results of trawl surveys increased the number of these species to 72, mainly based on the expansion of research works in the northern part of the sea (Table 1). At the same time, only 16 species from six families were relatively abundant and formed a significant biomass in different years; among them, cod fishes were absolutely dominant (Table 2) (on average, 92.6% of the total commercial biomass of marine fishes). It should be noted that, whereas in the late 1990s–early 2000s, the stock of this family was formed mainly from polar cod, its biomass significantly decreased in the late 2010s; however, the stock of Walleye pollock sharply increased by that time, which was determined (as already mentioned above) by its migrations to the study area from the northern part of the Bering Sea.
Fishes of other families are not so abundant; however, in a number of years, the biomass of some of them (capelin, Bering flounder, and shorthorn sculpin Myoxocephalus verrucosus) exceeded the stocks of cod fishes, including polar cod and Walleye pollock. One can also mention another most abundant representative of the ichthyofauna of the Chukchi Sea, namely, the Arctic staghorn sculpin Gymnocanthus tricuspis, which cannot be of interest to fisheries because of its small size.
As an explanation of the data in Table 2, it should be noted that Pacific cod, saffron cod, and flatfishes (Pleuronectidae), which are part of bottom and near-bottom communities, were not predictably recorded in catches of midwater trawls in 2003, 2007, and 2008. Conversely, salmonids (Salmonidae), living mainly in surface water masses, were not caught in bottom trawls. At the same time, Walleye pollock, polar cod, herring, and capelin, which form significant aggregations in the pelagic zone, were recorded in commercial quantities even near the bottom under shallow water conditions of the Chukchi Sea shelf in some years. This feature is also typical for these species in other areas of their habitat (Shuntov et al., 1993; Datsky and Andronov, 2007; Nikolaev et al., 2008; Kuznetsov et al., 2013; Loboda and Zhigalin, 2017; Velikanov, 2018).
On the whole, it can be said with high probability that the above-mentioned species of marine fishes are among the species that form the basis of the ichthyofauna of Russian waters of the Chukchi Sea. In addition, the known values of the biomass of some of them (primarily saffron cod Eleginus gracilis, capelin, rainbow smelt Osmerus mordax dentex, keta salmon O. keta, sockeye salmon, and starry flounder Platichthys stellatus) may increase as a result of an increase in the water area, research period, and amount of fishing gear. It is also necessary to note that polar cod, saffron cod, Bering and starry flounders, herring, capelin, smelt, and shorthorn sculpin can most likely be considered permanent inhabitants of the Chukchi Sea. At the same time, Walleye pollock, Pacific cod, black halibut, other flounder species, and Pacific salmons are temporary dwellers of the Chukchi shelf; their abundance and biomass in this area largely depend on abiotic and biotic environmental factors and the intensity of their migration from the Bering Sea. Based on this assumption, the raw material source for marine fishes in this water body, apparently, cannot be considered stable for potential fishery use, although it can be very significant and highly demanded for different types of fishery in some years.
Features of the Biology of Fishes in Russian Waters of the Chukchi Sea. Cod Family (Gadidae)
Walleye pollock. Information on the occurrence and biology of Walleye pollock in the Chukchi Sea was almost completely absent until the early 2000s. Anecdotal reports indicated an insignificant amount of juveniles (mainly young-of-the-year) in the southwestern part of the sea, which entered here with warm currents through the Bering Strait from the Bering Sea, where the main spawning grounds and feeding areas for groups of this species are located (Wolotira et al., 1977; Shuntov et al., 1993; Stepanenko, 1997; Gritsai, 2003; Stepanenko and Gritsai, 2016, 2018). Climate changes in the 2000s–2010s (particularly significant in 2017–2019) led to a redistribution of Walleye pollock: its lower abundance than the historical average values was recorded in the southern areas of the Bering Sea shelf with a simultaneous increase in the number of mature individuals in the north of the sea (Eisner et al., 2020; Baker, 2021). Apparently, the adaptation of Walleye pollock to changing environmental conditions, coupled with the search for a more accessible food supply, led to the migration of some individuals of this species to the shelf of the Chukchi Sea, as evidenced by our data (Fig. 2).

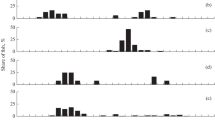
Size ((a)–(f)) and age ((g)–(l)) composition of Walleye pollock Gadus chalcogrammus in trawl catches from Russian waters of the Chukchi Sea: (a), (g) August 2003 (M = 57.9 cm, mean age 9.4 years, n = 63 ind.); (b), (h) August 2007 (M = 9.6 cm, 0.6 years, n = 39 ind.); (c), (i) September 2008 (M = 10.8 cm, 0.7 years, n = 32 ind.); (d), (j) September 2018 (M = 39.8 cm, 6.0 years, n = 944 ind.); (e), (k) August 2019 (M = 51.6 cm, 8.1 years, n = 3957 ind.); (f), (L) August–September 2020: (◼) and (◻) bottom scientific trawlings (M = 43.4 cm, 6.7 years; n = 5080 ind.), (– ○ –) pelagic commercial trawlings (M = 54.1 cm, 8.5 years, n = 886 ind.).
Thus, Walleye pollock individuals with FL > 40 cm were recorded in the area of the Chukchi shelf for the first time in 2003; they were dominated by individuals with FL 50–60 cm (58.7% of all catches) at the age of 7–10 years (Figs. 2a, 2g). With rare exception, individuals with FL 7–14 cm, slightly dominated by 1-year-olds, were recorded in trawl catches in 2007–2008 (Figs. 2b, 2c, 2h, 2i). In 2018–2020, mature Walleye pollock, dominated by individuals with FL 50–65 cm, appeared again in catches: in different years, their proportion was 52.8–74.4%. Its age was 1–16 years; individuals at the age of 8–10 years (the 2007–2012 generation) prevailed (Figs. 2d–2f, 2j–2l). Taking into account the shallow depths of the study area, the size–age characteristics of Walleye pollock captured near the bottom and in the water column were significantly similar, except a smaller number of juveniles and extremely large individuals with FL > 65 cm at the age of 13–15 years in pelagic trawl catches (Figs. 2f, 2l). The data on the appearance of mature Walleye pollock within the Chukchi shelf in certain years are quite consistent with the data indicating the alternation of warm (2002–2005 and 2014–2019) and cold (2006–2013) years (Andrews et al., 2016; Baker and al., 2020; Danielson et al., 2020; Yasumiishi et al., 2020; Baker, 2021).
On the whole, individuals with FL 9–14 and 50–65 cm at the age of 2.8–12 years were dominant in Russian waters of the Chukchi Sea throughout the study period; their proportion was more than 82% (Fig. 3). A distinctive feature of the Walleye pollock feeding here was the almost complete absence of 2–5-year-old individuals with FL 22–45 cm. At the same time, the proportion of this species in the northwestern part of the Bering Sea, adjacent to the Chukchi shelf, was ≥ 69% of all catches, while older Walleye pollock, on the contrary, was recorded in minimum amounts (Figs. 3c, 3f). It should be noted that the averaged size–age fish composition insignificantly differed in the pelagic zone and at the bottom within the Chukchi shelf: the proportion of 1- and 8-year-old individuals was high only in the second biotope (Figs. 3a–3e). The pollock distribution in the water column significantly differed in the adjacent Bering Sea: the pelagic zone was dominated by 2- to 4-year-old individuals with FL 25–40 cm; the mode was 33–35 cm at the age of 2–4 years. Near the bottom, Walleye pollock was concentrated more evenly and the number of individuals with FL 9–18 and 37–43 cm at the age of 2–3 and 5–6 years, respectively, increased here. Apparently, this is explained by the physical and geographical features of the northwestern part of the Bering Sea, where the vast shallow Gulf of Anadyr in combination with a significant continental slope to the south of Cape Navarin create favorable conditions for the feeding of medium-sized Walleye pollock.

Size ((a)–(c)) and age ((d)–(f)) composition of Walleye pollock Gadus chalcogrammus in cumulative samples from pelagic ((a), (d)), bottom ((b), (e)), and mixed ((c), (f)) trawl catches from Russian waters in 1996–2020: (◼), (◻) Chukchi Sea; (–○–) Bering Sea.
Taking into account that Walleye pollock appears in the study area as a result of migrations from the Bering Sea through the Bering Strait, we compared the size–age composition of individuals from different water areas of the northwestern part of the Bering Sea with materials from the Chukchi Sea (Figs. 1, 4). It was noted that the size–age composition of Walleye pollock was most similar in the Chukchi waters and central part of the Gulf of Anadyr, which is largely determined by the features of the bottom topography and pattern of currents in these water areas. In both cases, there is a relatively gently sloping shallow shelf (Udintsev et al., 1959; Moiseev and Grinfeld, 2019) with favorable feeding conditions for Walleye pollock with FL > 50 cm at age above 7+, while the warm Navarin Current contributes to the formation of aggregations of young-of-the-year and one-year-old individuals here (Datsky and Andronov, 2007; Kuznetsov et al., 2013). This current also contributes to the accumulation of euphausiids (Euphasiidae) and copepods (Copepoda), which are the basic items of the diet of Walleye pollock juveniles (Shuntov et al., 1993; Orlov et al., 2019). The occurrence of large-sized individuals in the central part of the gulf is determined by the presence of Pacific sand lance Ammodytes hexapterus and humpy shrimp Pandalus goniurus, available for food (Zgurovskii and Khen, 1988; Chikilev and Datsky, 2000; Andronov and Datsky, 2019); in Chukchi waters, their occurrence is explained by the presence of numerous small sculpin species of the genus Gymnocanthus and slender eelblenny Lumpenus fabricii in addition to the above-mentioned species. On the whole, the migration of mature Walleye pollock to the Russian waters of the Chukchi Sea in 2018–2020 was facilitated by the general warming of air and water masses and a decrease in the area of sea ice and its melting period, which are characteristic of the late 2010s (Baker et al., 2020; Danielson and al., 2020; Carvalho et al., 2021), as well as by a sufficient available food supply in this part of the sea shelf (Sirenko and Gagaev, 2007; Slabinskii and Figurkin, 2014; Grebmeier et al., 2015; Kuznetsova and Gorbatenko, 2021). For instance, according to trawl surveys in 2017 and 2019, large Walleye pollock was also recorded in the US sector of the sea; however, its abundance was minimal here (Farley et al., 2017; personal communication of I.V. Grigorov).

Size ((а)–(e)) and age ((f)–(j)) composition of Walleye pollock Gadus chalcogrammus in bottom trawl catches from Russian waters in 2001 and 2019: (- ‑), (◻) Chukchi Sea (M = 51.6 cm, mean age 8.1 years, n = 3957 ind.); (—), (◼) Bering Sea: (a), (f) Chirikov basin (M = 15.8 cm, 1.3 years, n = 409 ind.); (b), (g) northern (M = 48.4 cm, 8.2 years, n = 4567 ind.); (c), (h) the central part of the Gulf of Anadyr (M = 23.5 cm, 2.9 years, n = 7912 ind.); (d), (i) the western part of the Gulf of Anadyr (M = 33.7 cm, 4.2 years, n = 14 167 ind.); (e), (j) Koryak area (M = 20.3 cm, 1.7 years, n = 51 159 ind.).
Immature individuals in trawl catches from the shelf of the Chukchi Sea had FL 7.5–24.0 cm and a body weight of 1–10 g. Among mature individuals, females were larger than males; their FL was 25.3–82.0 cm and weight was 106–4950 g. The number of males with FL 25.0–76.1 cm and a weight of 97–2506 g was lower (by two–three times) than that of females (Table 3). On the whole, the greater size of females and their predominance among older Walleye pollock individuals are typical for the species under consideration, which is confirmed by data on its other habitats (Shuntov et al., 1993; Datsky, 2016). The dependence of the weight (W, g) of Walleye pollock on its length (FL, cm) in the study area is well described by the equation W = 0.004FL3.1335 (r = 0.99, n = 606 ind.). This ratio is closest to the value for the fish from the central part of the Gulf of Anadyr: W = 0.0031FL3.1802 (r = 0.94, n = 52 ind.).
The growth of Walleye pollock is uneven throughout its life. According to 2018–2019 data, the greatest increase in the body length of the fish on the shelf of the Chukchi Sea was observed in the 2nd and 3rd years of its life: 8.0 and 3.9 cm per year, respectively. Quite high rates of linear growth were also recorded in the 8th year of life (3.7–4.8 cm), followed by their decrease in extremely large individuals with FL > 70 cm at the age of 13 years or older. The weight gain is maximal in 8- and 10–11-year-old individuals: 240–250 and 237–521 g, respectively (Table 4).
On the whole, the results confirm the life strategy of Walleye pollock for the greatest linear increase in the first two–three years and further gradual decrease by the age of seven years. This is determined by the period of sexual maturity, when a significant part of energy is spent on the maturation of reproductive products rather than on growth, as in the first three years of life. At the age of 8–9 years, the range of the diet of Walleye pollock significantly changes; it begins to feed mainly on fish objects, which undoubtedly contributes to an increase in its linear and weight gain. As pollock reaches the age limit, all the functions of its body are aimed at maintaining the necessary metabolism (Datsky, 2016); this allows Walleye pollock to reach FL 91 cm, a weight of 5.2 kg, and the age of 28 years within its range (Eschmeyer et al., 1983; Munk, 2001; Datsky, 2016). It should also be noted that Walleye pollock captured on the shelf of the Chukchi Sea had a slower rate of growth length than the individuals from the northwestern part of the Bering Sea at the age below 13 years (Fig. 5), which probably indicates less favorable conditions for its habitat on the northern periphery of its range.

Linear growth of Walleye pollock Gadus chalcogrammus from Russian waters of the Chukchi Sea and northwestern part of the Bering Sea in 1995–2019: (- -) Chukchi Sea (FL = 11.199t0.7322, r = 0.98, n = 478 ind.), (—) Bering Sea (FL = 17.812t0.5447, r = 0.96, n = 1783 ind.).
There is no information on Walleye pollock reproduction in the Chukchi Sea. The closest spawning area to the studied water area was recorded in the north of the Gulf of Anadyr (Datsky, 2004). The spawning period within the Bering Sea is seven months (from February to September) (Datsky and Andronov, 2007). At the same time, a relatively large proportion of Walleye pollock individuals with gonads at maturity stages VI–II indicates their recent spawning. This can be indirectly confirmed by the presence of its larvae in the northeastern part of the Chukchi Sea in August–September (Logerwell et al., 2020), which might result from their removal to this water area by currents. The largest number of post-spawned individuals was recorded in August; their proportion naturally decreased by September (Table 5); the number of post-spawned females was higher than that of males. Post-spawned females actively fed and matured: in August, up to 18% of females and 48% of males had gonads at maturity stages I–II; in September, the proportions of these individuals increased to 49 and 71%, respectively.
Polar cod is a cryopelagic species that constantly lives and reproduces in Arctic waters, unlike Walleye pollock (Manteifel, 1943; Borkin et al., 1987; Novikov et al., 1992; Karamushko, 2020). Climate changes towards warming led to a significant reduction in the range of this species, as well as to a general decrease in commercial fish biomass, in the northern part of the Bering Sea and in the Chukchi Sea in the late 2010s (Savin, 2021; Baker, 2021) (Table 1); however, polar cod is still one of the mass species among the fish communities on the Chukchi shelf.
Individuals with FL 3–29 cm at ages 0+ to 6+ were recorded in Russian waters of the Chukchi Sea. At the same time, catches of polar cod during the period of its high abundance in 1995, 1997, and 2003 were represented mainly (about 88.4–92.5%) by 2–3-year-old individuals with FL 11–19 cm (Figs. 6a–6c). The abundance of these individuals then decreased to 74.1–78.0%; a large number of young-of-the-year with FL 4–7 cm were recorded in the pelagic zone (Figs. 6d–6i), which is also generally typical of the northeastern part of the sea (De Robertis et al., 2017). The total size range of polar cod was characterized by the presence of two maxima: fish with FLs 5–7 and 9–15 cm. As a result, comparison of the summarized data for a number of years revealed a greater size of individuals recorded near the bottom (Fig. 7). This is also confirmed by earlier studies of polar cod in the Chukchi Sea (Nikolaev et al., 2008; Helser et al., 2017; Savin, 2021).

Size composition of polar cod Boreogadus saida in trawl catches from Russian waters of the Chukchi Sea: (a) August 1995 (M = 15.9 cm, n = 816 ind.), (b) August–September 1997 (M = 14.9 cm, n = 958 ind.), (c) August 2003 (M = 11.8 cm, n = 2261 ind.), (d) August 2007 (M = 8.7 cm, n = 4589 ind.), (e) September 2008. (M = 6.7 cm, n = 1734 ind.), (f) September 2010 (M = 11.4 cm, n = 3910 ind.), (g) September 2018 (M = 11.7 cm, n = 3943 ind.), (h) August 2019 (M = 13.3 cm, n = 436 ind.); (i) August–September 2020: (◼) bottom scientific trawlings (M = 11.8 cm, n = 3264 ind.), (–○–) pelagic commercial trawlings (M = 11.1 cm, n = 41 ind.).

Size composition of polar cod Boreogadus saida in cumulative samples from pelagic (a), bottom (b), and mixed (c) trawl catches from Russian waters of the Chukchi (◼) and Bering (◻) seas in 1996–2020.
In comparative terms, polar cod from the Chukchi Sea is smaller than that living in the northwestern part of the Bering Sea. In addition to the above-mentioned size groups, large-sized individuals with FL 17–21 cm, represented mainly by 4-year-olds, were found among the individuals from the northwestern part of the Bering Sea, with the greatest differences being recorded between individuals captured in the pelagic zone (Fig. 7). According to other studies, polar cod with the maximum FL of 32 cm was encountered in the central part of the Gulf of Anadyr of the Bering Sea; 20, 28, 31, 40, and 40 cm (Shleinik, 1979; Coad and Reist, 2004; Antonov et al., 2016; Glebov et al., 2016; Gorbatenko and Kiyashko, 2019) in the East Siberian, Laptev, Kara, Beaufort, and Barents seas, respectively (Savin, 2021). In the latter case, it was a fish weighing 430 g at the age of 10 years (Shleinik, 1979).
It is believed (Bondarev, 1970) that polar cod forms a single Chukchi–Bering-Sea group and feeding migrations of predominantly older individuals from the Chukchi Sea to the Bering Sea can be observed under the condition of its high abundance (as a rule, in cold years) (Fadeev, 1986; Nikolaev et al.., 2008). Indeed, polar cod is somewhat larger in the waters of the northwestern part of the Bering Sea than on the Chukchi shelf, in particular, in the southwestern part of the Gulf of Anadyr and off the Koryak coast (Fig. 8). At the same time, small-sized individuals with FL 5–10 cm were also recorded in significant numbers in the northern part of the gulf (Fig. 8b). Taking into account that the polar cod biomass has significantly decreased and its aggregations have almost completely disappeared in the area of the Bering Strait since 2007 (Nikolaev et al., 2008; Orlov et al., 2019), it can be assumed that a relatively independent group of the species lives in winter cold waters in the northern part of the Gulf of Anadyr. In years with high abundance, it can be recruited by spawners from the Chukchi Sea and it spawns in the Kresta Bay in the northwestern part of the Bering Sea. The confinement of polar cod aggregations to this water area was also noted according to distribution maps in different study years (Datsky and Andronov, 2007). The scale of migrations to this area is more dependent on the yield of separate generations and total abundance of the species.

Size composition of polar cod Boreogadus saida in bottom trawl catches from Russian waters in 2001 and 2019: (- -) Chukchi Sea (M = 13.3 cm, n = 4360 ind.); (—) Bering Sea: (a) Chirikov basin (M = 16.9 cm, n = 242 ind.); (b)–(d) the northern (M = 15.9 cm, n = 1097 ind.), central (M = 18.2 cm, n = 224 ind.), and western (M = 17.6 cm, n = 30 ind.) parts of the Gulf of Anadyr, respectively; (e) Koryak area (M = 19.3 cm, n = 161 ind.).
The size of immature individuals in trawl catches on the shelf of the Chukchi Sea was 3.4–12.9 cm at a body weight of 1 to 11 g. Females were larger than males during all observation years, reaching the maximum length and weight of 28.2 cm and 149 g (Table 6). This feature is typical for the species within its range: all the individuals with the maximum size and age (7–8 years) were represented exclusively by females (Borkin et al., 1987; Karamushko, 2020). Another pattern was that the size–weight values of the body were higher for polar cod from bottom catches than for the individuals from pelagic aggregations. This was observed during almost all observation years both in mixed catches and separately in females and males. The length–weight dependence for polar cod within the Chukchi shelf is as follows: W = 0.0056FL3.0774 (r = 0.98, n = 3052 ind.).
Polar cod grows slowly compared to other representatives of the cod family; it reaches FL 18–20 cm at the age of 5 years and 25–27 cm at the age of 7 years (Benko and Ponomarenko, 1972). A high rate of linear growth was recorded in younger age groups from the Chukchi and Bering seas, which decreases during the onset of sexual maturity in the 3rd year of life (Cherkasova, 1976). The slower growth of polar cod at age above 2–3 years is also recorded for the Beaufort Sea (Craig et al., 1982) and Allen Bay in the Canadian Arctic (Matley et al., 2013). A progressive decrease in the annual absolute increase in length after reaching sexual maturity was also recorded for polar cod from the Barents Sea (Shleinik, 1979).
Unlike the linear growth, the absolute growth in the body weight of polar cod consistently increases with age, reaching the maximum value after the onset of its sexual maturity, which is undoubtedly an important condition for the production of the maximum amount of reproductive products and an increase in population fecundity. For instance, in the Laptev Sea, the increase in the relative rate of weight growth was particularly significant for individuals with FL 20–23 cm at the age of 5–6 years: from 7 to 16 g per year (Karamushko, 2020). Within the Russian waters of the Chukchi shelf, the greatest weight increase was observed in individuals with FL 19–24 cm (when their weight increased by 9–11 g per 1 cm of their body length).
Polar cod is a fast maturing fish; it reaches sexual maturity at the age of 2–3 years at FL ~13 cm (Moskalenko, 1964; Borkin et al., 1987). Individuals that live in the southeastern coastal part of the Barents Sea and in the Kara and Laptev seas mature at the age of 2 years at FL 8–14 cm (Moskalenko, 1964; Karamushko, 2020). In the Chukchi Sea, the seasonal maturation of gonads in August–September is observed in both sexes starting from the size class of 8–9 cm and reaches 50% of maturation in males with FL 16 cm and in females with FL 19 cm or more (Savin, 2021). As in all cod fishes, there are more males among small-sized individuals and females with a greater size than that of males begin to prevail with age. The number of females in aggregations is also higher than that of males: the sex ratio is 1.1–1.3 : 1.0 (Table 6).
In the Chukchi and Beaufort seas, polar cod spawns for the first time at FL ~12.5 cm at the age of 3 years (Frost and Lowry, 1983) near the coast, mainly under ice in January–February (Manteifel, 1943; Musienko, 1970). The opinion about earlier spawning of the fish in the Chukchi Sea and northern part of the Bering Sea (in October–December) was not confirmed (Cherkasova, 1976; Savin, 2021). According to our data, polar cod spawners in August were characterized by the presence of gonads predominantly at maturity stage II and their mass maturity (more than 50% of the studied individuals) (stage III) started only in September. There were more males among these individuals (Table 7), which only confirms their earlier readiness for breeding: in polar cod in particular and in cod fishes in general.
There is no information on the reproduction of this species in the Russian part of the Chukchi Sea; however, polar cod larvae were recorded in the northeastern part of the sea in August–September (Logerwell et al., 2020). In the Bering Sea, the spawning sites of the species are mainly in the northern part of the Gulf of Anadyr, in the Kresta Bay, and near the Bering Strait (Bondarev, 1970). Larvae and juveniles (with FL 8–19 and 20–40 mm, respectively) are also concentrated here above the depths of 37–63 m at temperatures from –1.9 to 0.4°C (Bulatov, 1986).
Saffron cod has a relatively small body size among the cod fishes in the Far Eastern seas and adjacent water areas and occupies the intermediate position between the small polar cod and larger Walleye pollock and Pacific cod. The maximum length, weight, and age of this species generally do not exceed 57 cm, 1.3 kg, and 15 years, respectively (Datsky and Andronov, 2007). Off the Arctic coast of Canada, saffron cod reaches FL 63 cm (Coad and Reist, 2004). The maximum size was recorded for individuals from waters off the western coast of Kamchatka and in the northwestern part of the Bering Sea, while the FL of saffron cod in the Chukchi Sea did not exceed 37 cm, its weight was not more than 420 g, and age not more than 15 years (Semenenko, 1965).
Indeed, saffron cod in trawl catches from the Chukchi shelf was represented mainly by young-of-the-year and 1-year-old individuals with FL 5–16 cm (Fig. 9). According to the data of 2020, young-of-the-year with FL 5–9 cm preferred the water column, while 1-year-old individuals with a larger body size were concentrated near the bottom. In this year, individuals with FL 16–25 cm were recorded in catches for the first time; however, their occurrence was sporadic (Fig. 9d).

Size composition of saffron cod Eleginus gracilis in trawl catches from Russian waters of the Chukchi Sea: (a) September 2008 (M = 7.7 cm, n = 74 ind.), (b) September 2018 (M = 9.4 cm, n = 43 ind.), (c) August 2019 (M = 10.3 cm, n = 3 ind.); (d) August–September 2020: (◼) bottom scientific trawlings (M = 12.7 cm, n = 157 ind.); (–○–) pelagic commercial trawlings (M = 7.2 cm, n = 51 ind.).
The difference is even greater in the size between saffron cod from the southwestern part of the Chukchi Sea and saffron cod from the northwestern part of the Bering Sea (Fig. 10). In addition to the low abundance of this species, bottom trawl catches from the Chukchi shelf contained almost exclusively juveniles with FL 5–19 cm at ages 0+ to 2+ (99.7%), the proportion of which was insignificant in the more southern water area (10.8%) and which was dominated by individuals with FL 29–39 cm (69.8%) at the age of 4–6 years (Datsky, 2016). At the same time, according to 2012 data, bottom and midwater trawl catches from the southeastern part of the Chukchi Sea contained saffron cod individuals with FL 3–34 cm at ages 0+ to 7+ (with a high occurrence (up to 50% near the bottom) of individuals with FL > 19) (Helser et al., 2017).

Size composition of saffron cod Eleginus gracilis in bottom trawl catches from Russian waters in 1996–2020: (◼) Chukchi Sea (M = 12.0 cm, n = 203 ind.), (◻) Bering Sea (M = 31.3 cm, n = 4179 ind.).
The size composition of saffron cod significantly changed in the northern direction (Fig. 11). Whereas fish aggregations in the Olyutor–Navarin area were formed mainly from individuals with FL 30–44 cm (about 87% of all catches) and the abundance of juveniles and individuals maturing for the first time was low, there were almost no individuals with FL > 30 cm in the western part of the Gulf of Anadyr, while individuals with FL 18–25 cm were dominant here (93.3%). In the north of the gulf, saffron cod had an even lower body size and young-of-the-year with FL 5–7 cm were absolutely dominant here (>66%); the occurrence of other size groups was insignificant in the catches. The occurrence of juveniles in this area can be explained by the close proximity of the Kresta Bay, the bottom topography, banks, and hydrological regime of which are probably favorable for the reproduction of saffron cod and further habitation of its young-of-the-year. It is quite possible that some of young-of-the-year will be carried by currents from the northern part of the Gulf of Anadyr to the area of the Chirikov basin and some of them to the Chukchi Sea in the future. On the whole, the trend towards a decrease in the size of saffron cod in catches towards the north is also confirmed by data from the eastern parts of the Bering and Chukchi seas (De Robertis et al., 2017).

Size composition of saffron cod Eleginus gracilis in bottom trawl catches from Russian waters in 2001 and 2018–2020: (- -) Chukchi Sea (M = 12.0 cm, n = 203 ind.); (—) Bering Sea: (a) and (b) the northern (M = 10.8 cm, n = 213 ind.) and western (M = 22.1 cm, n = 208 ind.) parts of the Gulf of Anadyr, respectively; (c) Koryak area (M = 35.3 cm, n = 3413 ind.).
Taking into account that juveniles prefer shallow water areas and the proportion of large individuals increases with depth (Datsky, 2016), it can be assumed that the Chukchi shelf within Russian waters is a feeding area mainly for immature fish. For adult representatives of the species, there are probably insufficient shallow bays and gulfs that would be favorable for spawning (except the Neshkanskaya Lagoon and Kolyuchin bay, at the output of which small aggregations of saffron cod were recorded); harsh environmental conditions in the study area are also an important limiting factor.
The greatest linear growth of saffron cod was recorded in the first four years of its life, followed by an annual increase in its body length at the rate of not more than 2–3 cm. Its weight significantly increases at the age of 5–8 years; its further growth is insignificant (Datsky, 2016). Saffron cod grows more slowly in the Chukchi Sea than in the northern part of the Bering Sea, especially at a younger age (up to age 3+). Its average FL value in the eastern part of the Chukchi shelf at ages 0+, 1+, 2+, 3+, and 4+ does not exceed 5–7, 13, 20, 24, and 28 cm, respectively, while it is not more than 9, 17, 23, 25, and 28 cm in the north of the Bering Sea within American waters (Helser et al., 2017). An even higher rate of linear growth was recorded for saffron cod from the Olyutor–Navarin area in the Bering Sea: at ages 3+ and 4+, it reaches FL 33 and 38 cm, respectively (Datsky, 2016).
The Far Eastern saffron cod reaches sexual maturity at the end of the second year of its life at FL 18–26 cm. The latest sexual maturation in the Chukchi Sea was recorded for individuals from the Neshkanskaya Lagoon with a low growth rate: mostly in the 4th–5th year of life after reaching FL > 21 cm (Safronov, 1986). The gonads of saffron cod mature from August to winter; males prepare for the reproduction process more rapidly than females (Datsky, 2016). In our case, all the studied individuals had gonads at maturity stage II in August–September; the number of females was higher than that of males and their body size was larger (Table 8). The length–weight dependence for saffron cod is described by the equation: W = 0.0019FL3.5346 (r = 0.98, n = 15 ind.).
In December–February, saffron cod spawns at negative water temperatures: –1.0 to –1.9°C (Bulatov, 1986; Safronov, 1986). The number of spawning individuals in the bays and gulfs of the Chukchi Peninsula within the Bering Sea waters was maximal in early December. Single captures of larvae of this species were recorded in the eastern part of the Chukchi Sea (Logerwell et al., 2020).
Pacific cod is a commercial fish species that reaches FL 120 cm, a weight of 22.7 kg, and an age of 25 years within its range (Morrow, 1980; Lamb and Edgell, 1986; Munk, 2001) and occurs in fishery catches from Russian waters mainly with FL 30–60 cm at the age of 3–6 years (Bogdanov, 2006). It forms aggregations in shallow areas of the Bering Sea shelf, including bays and gulfs; as a rule, these aggregations consist of juveniles and individuals maturing for the first time with FL < 40 cm and, to a lesser extent, middle-aged and older individuals preferring depths over 10 m) (Datsky and Andronov, 2007). Probably for this reason, the features of the Chukchi shelf (vast bottom areas with shallow depths, a weak indentation of the coastline, and severe oceanological conditions) do not contribute to the formation of aggregations of mature cod in this area.
As with saffron cod, Pacific cod recorded in the study area at the bottom and in the pelagic zone is mainly immature, with FL 10–17 and 5–9 cm at ages 0+ to 1+, respectively (Fig. 12). The appearance of a small number of juveniles is apparently facilitated by warm currents through the Bering Strait from the Bering Sea, from which they are carried to the shelf of the Chukchi Sea. At the same time, unlike Walleye pollock, the general warming in recent years has had little effect on the scale of migration of mature Pacific cod to the periphery of the range, although its main aggregations in the Bering Sea also shifted north of its common range (Stevenson and Lauth, 2019; Baker, 2021). Whereas Pacific cod was absent in trawl catches from the Chukchi shelf in the early 2000s both according to our data and according to the materials of other researchers (Mecklenburg et al., 2016), juveniles with FL 10–18 cm that appeared here in 2018–2020 were accompanied by only a few individuals with FL 55–78 cm (Table 2, Fig. 12). This can be observed even more clearly if we compare the size ranges of individuals from the shelves of the Chukchi and Bering seas (Fig. 13). It is clearly seen that there are Pacific cod individuals of all sizes in the Bering Sea, which indicates more favorable living conditions for this species here, unlike the study area. The greatest concentrations of these fish groups were recorded along the Koryak coast and in the western part of the Gulf of Anadyr; a decrease in the abundance of medium-sized Pacific cod in catches with increase in the proportion of immature individuals was recorded in the northeastern direction (Fig. 14). In addition to juveniles, individuals with the maximum size generally migrate to the Chukchi shelf, mostly from the adjacent bays of the Chukchi Peninsula, where they probably lead a sedentary bottom lifestyle, which is generally typical of cod and pollock (Shuntov et al., 1993).). In this case, it can be assumed that significant migrations of biomass of older Walleye pollock to the study area in 2018–2020 hinder the feeding of Pacific cod in this area, which is limited not so much by the interannual variability in the temperature of bottom water masses as by the availability of food supply (Baker, 2021). At the same time, the compositions of food for Walleye pollock and Pacific cod with FL > 50 cm are quite similar in the northwestern part of the Bering Sea: in addition to decapods (Decapoda) and bivalve mollusks (Bivalvia) (only in Pacific cod), their main food items include fish objects, such as polar cod, capelin, Pacific sand lance Ammodytes hexapterus, and Walleye pollock juveniles (Datsky and Andronov, 2007).

Size composition of Pacific cod Gadus macrocephalus in trawl catches from Russian waters of the Chukchi Sea: (a) August 2007 (M = 6.6 cm, n = 23 ind.), (b) September 2018 (M = 24.4 cm, n = 31 ind.), (c) August 2019 (M = 24.0 cm, n = 16 ind.); (d) August–September 2020: (◼) bottom scientific trawlings (M = 13.1 cm, n = 6 ind.), (–○–) pelagic commercial trawlings (M = 7.0 cm, n = 2 ind.).

Size composition of Pacific cod Gadus macrocephalus in bottom trawl catches from Russian waters in 2001–2020: (◼) Chukchi Sea (M = 20.0 cm, n = 53 individuals), (◻) Bering Sea (M = 44.6 cm, n = 593 ind.).

Size composition of Pacific cod Gadus macrocephalus in bottom trawl catches from Russian waters in 2001, 2018, and 2019: (- -) Chukchi Sea (M = 23.0 cm, n = 53 ind.); (—) Bering Sea: (a)–(c) the northern (M = 48.9 cm, n = 58 ind.), central (M = 49.3 cm, n = 12 ind.), and western (M = 41.9 cm, n = 93 ind.) parts of the Gulf of Anadyr; (d) Koryak area (M = 44.3 cm, n = 431 ind.).
Among the few cod representatives, immature individuals had FL 6–18 cm at a weight of 1–48 g (e.g., fish with FL 14 and 18 cm at age 1+ had a weight of 22 and 48 g, respectively). According to 2018–2019 data, older individuals were represented by females with FL 75 and 77 cm and a weight of 5305 and 5318 g and males with FL 55, 71, and 78 cm with a weight of 270, 4790, and 4110 g, respectively. All of them were in the 7th year of life except the individual with FL 55 cm. The sex ratio was approximately equal. The size–weight dependence of Alaska cod within the Chukchi shelf is as follows: W = 0.0053FL3.1915 (r = 0.99, n = 23 ind.).
Cod females reach sexual maturity for the first time at FL 55–85 cm and males at FL 48–80 cm at the age of 3–8 and 4–9 years, respectively (Kim Sen Tok, 1998; Fadeev, 2005). Mass sexual maturation of the fish (≥50% of individuals) is observed at average FL 70 cm at the age of 6 years (Fadeev, 2005). In the Bering Sea, mass Pacific cod maturation in the closest water area to the study zone is recorded for individuals at the age of 5–7 years when the FL of females and males is 60–65 and 55–65 cm, respectively (Vershinin, 1987; Witherell, 2000; Vinnikov et al., 2013).
The spawning of Pacific cod and further development of its eggs and larvae within its range cover the period from January to May, usually during ice formation (Moiseev, 1953). In the north of the Bering Sea, the spawning period shifts to spring (from March to May); however, it may be completed even in the summer period when the year is cold with respect to hydrological conditions (Datsky, 2016). Apparently, this is also true for the Chukchi shelf, where most of the Pacific cod females and males had gonads indicating the recent end of the spawning period (maturity stages VI–II). In single individuals (females), the reproductive products were at the stage of maturity onset.
The spawning sites of Pacific cod in the Chukchi Sea have not yet been found; however, taking into account the absence of a great number of spawners in this water area, it can be assumed that the species spawns in small quantities only in the bays and gulfs of the Chukchi Peninsula south of the Bering Strait (St. Lawrence, Mechigmen, Tkachen, Pengingei, and Abolishev bays). The absence (or insignificant areas) of spawning sites in the Chukchi Sea and adjacent water areas is also confirmed by studies in the eastern part of the sea, when Pacific cod was not found in numerous collections of eggs, larvae, and young-of-the-year among fish species from 11 families, including cods (Logerwell et al., 2020). In the Bering Sea, the main spawning grounds of Pacific cod are outside the coastal waters at depths of 10–250 m; the temperature in its spawning sites is 0–3°C. In June, Pacific Cod larvae occur at depths of 160–130 m at a water temperature of 1.7–2.2°C. The quantitative indicators are low: the maximum density of larvae is 6 ind./m2 in the Navarin area and 5 ind./m2 in the eastern part of the sea. In June, the size of larvae varies from 10.0 to 15.6 mm (Bulatov, 1986).
Notes
Order no. 601 of the Ministry of Agriculture of the Russian Federation dated October 9, 2020 “On the Approval of the Total Allowable Catch of Aquatic Biological Resources in the Inland Sea Waters of the Russian Federation (in the Territorial Waters of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economic Zone of the Russian Federation, and in the Caspian Sea as of 2021.” (https://rulaws.ru/acts/Prikaz-Minselhoza-Rossii-ot-09.10.2020-N-601)
Order no. 267 of the Ministry of Agriculture of the Russian Federation dated May 23, 2019 (amended of June 27, 2019) “On the Approval of the Fishing Rules for the Far Eastern Fishery Basin”. (https://fish.gov.ru/wp-content/uploads/documents/ otraslevaya_deyatelnost/lubitelskoe_rybolovstvo/Pravila_Dalnevostochnogo_2.pdf)
Order of the Ministry of Agriculture of the Russian Federation dated May 23, 2019 No. 267.
REFERENCES
Aksyutina, Z.M., Elementy matematicheskoi otsenki rezul’tatov nablyudenii v biologicheskikh i rybokhozyaistvennykh issledovaniyakh (Elements of mathematical evaluation of the results of observations in biological and fishery research), Moscow: Pishch. prom-st’, 1968.
Andrews, A.G.III., Strasburger, W.W., Farley, E.V.Jr., et al., Effects of warm and cold climate conditions on capelin (Mallotus villosus) and pacific herring (Clupea pallasii) in the eastern Bering Sea, Deep-Sea Res. II: Top. Stud. Oceanogr., 2016, vol. 134, pp. 235–246. https://doi.org/10.1016/j.dsr2.2015.10.008
Andronov, P.Yu. and Datsky, A.V., Benthos, in Ekosistemy Beringova proliva i faktory antropogennogo vozdeistviya (Ecosystems of the Bering Strait and factors of anthropogenic impact), Moscow: World Wildlife Fund, 2019, pp. 101–111.
Antonov, N.P., Kuznetsov, V.V., Kuznetsova, E.N., et al., Arctic cod Boreogadus saida (Gadiformes, Gadidae) as a key species and potential fishery object in the Kara Sea, Vopr. Rybolovstva, 2016, vol. 17, no. 2, pp. 203–212.
Baker, M.R., Contrast of warm and cold phases in the Bering Sea to understand spatial distributions of arctic and sub-Arctic gadids, Polar Biol., 2021, vol. 44, pp. 1083–1105. https://doi.org/10.1007/s00300-021-02856-x
Baker, M.R., Kivva, K.K., Pisareva, M.N., et al., Shifts in the physical environment in the pacific arctic and implications for ecological timing and conditions, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vol. 177, Article 104802. https://doi.org/10.1016/j.dsr2.2020.104802
Basyuk, E. and Zuenko, Y., Extreme oceanographic conditions in the northwestern Bering Sea in 2017–2018, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vols. 181–182, Article 104909. https://doi.org/10.1016/j.dsr2.2020.104909
Benko, Yu.K. and Ponomarenko, V.P., Osnovnye promyslovye ryby Barentseva, Norvezhskogo i Grenlandskogo morei (biologiya, ulovy) (Main Commercial Fish of the Barents, Norwegian and Greenland Seas (Biology, Catches)), Murmansk: Izd. Polar. Inst. Rybn. Khoz. Okeanogr., 1972.
Bogdanov, G.A., Tikhookeanskaya treska. Voprosy biologii i zapasy (Pacific Cod. Biology and Stocks), Moscow: IPF “Sashko,” 2006.
Bondarev, B.M., Description of the Arctic cod, in Saika (Arctic Cod), Petropavlovsk-Kamchatskii: Kamchat. Otd. Dal’nevost. Kn. Izd., 1970, pp. 3–9.
Borisovets, E.E., Vdovin, A.N., and Panchenko, V.V., Estimated stocks of sculpins based on accounting trawl surveys in Peter the Great Bay, Vopr. Rybolovstva, 2003, vol. 4, no. 1 (13), pp. 157–170.
Borkin, I.V., Ponomarenko, V.P., Tret’yak, V.A., and Shleikin, V.N., Arctic cod Boreogadus saida (Lepekhin)— fish of the polar seas (stocks and use), in Biologicheskie resursy Arktiki i Antarktiki (Biological Resources of the Arctic and Antarctic), Moscow: Nauka, 1987, pp. 183–207.
Bowering, W.R. and Nedreaas, K.H., Age validation and growth of Greenland halibut (Reinhardtius hippoglossoides (Walbaum)): a comparison of populations in the Northwest and Northeast Atlantic, Sarsia, 2001, vol. 86, no. 1, pp. 53–68. https://doi.org/10.1016/j.fishres.2016.01.016
Brodziak, J. and Mikus, R., Variation in life history parameters of Dover sole (Microstomus pacificus), off the coasts of Washington, Oregon, and northern California, Fish. Bull., 2000, vol. 98, pp. 661–673.
Bulatov, O.A., Distribution of codfish eggs and larvae in the Pacific waters of Kamchatka and the western part of the Bering Sea, in Treskovye dal’nevostochnykh morei (Codfishes of the Far East Seas), Vladivostok: Izd. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1986, pp. 89–101.
Buslov, A.V. and Varkentin, A.I., Comparative characteristics of estimates of age and some population parameters of walleye pollock using scales and otoliths, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2001, vol. 128, pp. 164–176.
Carvalho, K.S., Smith, T.E., and Wang, S., Bering Sea marine heatwaves: patterns, trends and connections with the Arctic, J. Hydrol., 2021, vol. 600, Article 126462. https://doi.org/10.1016/j.jhydrol.2021.126462
Cherkasova, L.D., Size composition, growth and fertility of the Far Eastern Arctic cod, in Issledovaniya po biologii ryb i promyslovoi okeanografii. Vyp. 7 (Research in Fish Biology and Commercial Oceanography. Issue 7), Vladivostok: Izd. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1976, pp. 137–143.
Chikilev, V.G. and Datsky, A.V., Far Eastern multivertebral gerbil Ammodytes hexapterus (Ammodytidae) in the Gulf of Anadyr and adjacent waters, Vopr. Ikhtiol., 2000, vol. 40, no. 6, pp. 772–779.
Chilton, D.E. and Beamish, R.J., Age determination methods for fishes studied by the groundfish program at the Pacific Biological Station, Can. Spec. Publ. Fish. Aquat. Sci., 1982, vol. 60, p. 102.
Coad, B.W. and Reist, J.D., Annotated list of the arctic marine fishes of Canada, Can. MS Rep. Fish. Aquat. Sci., 2004, no. 2674, p. 112.
Craig, P.C., Griffiths, W.B., Haldorson, L., and McElderry, H., Ecological studies of Arctic cod (Boreogadus saida) in Beaufort Sea coastal waters, Can. J. Fish. Aquat. Sci., 1982, vol. 39, no. 3, pp. 395–406. https://doi.org/10.1139/f82-057
Danielson, S.L., Ahkinga, O., Ashjian, C., et al., Manifestation and consequences of warming and altered heat fluxes over the Bering and Chukchi Sea continental shelves, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vol. 177, Article 104781. https://doi.org/10.1016/j.dsr2.2020.104781
Datsky, A.V., Reproduction of pollock Theragra chalcogramma (Gadidae) in the Gulf of Anadyr, Vopr. Rybolovstva, 2004, vol. 5, no. 4(20), pp. 597−617.
Datsky, A.V., Fish fauna of the Chukchi Sea and perspectives of its commercial use, J. Ichthyol., 2015a, vol. 55, no. 2, pp. 185–209. https://doi.org/10.1134/S0032945215020022
Datsky, A.V., Ichthyofauna of the Russian exclusive economic zone of the Bering Sea: 1. taxonomic diversity, J. Ichthyol., 2015b, vol. 55, no. 6, pp. 792–826. https://doi.org/10.1134/S0032945215060065
Datsky, A.V., Biological features of the common fish species in Olyutorsky-Navarin region and the adjacent waters of the Bering Sea: 1. Gadidae (cods) family, J. Ichthyol., 2016, vol. 56, no. 6, pp. 868–889. https://doi.org/10.1134/S0032945216050039
Datsky, A.V., Biological features of the common fish species in Olyutorsky-Navarin region and the adjacent waters of the Bering Sea: 2. Families Macrouridae, Clupeidae, and Osmeridae, J. Ichthyol., 2017a, vol. 57, no. 1, pp. 106–121. https://doi.org/10.1134/S0032945217010027
Datsky, A.V., Biological features of the common fish species in Olyutorsky-Navarin region and the adjacent waters of the Bering Sea: 4. family sculpins (Cottidae), J. Ichthyol., 2017b, vol. 57, no. 3, pp. 341–353. https://doi.org/10.1134/S0032945217030031
Datsky, A.V. and Andronov, P.Yu., Ikhtiotsen verkhnego shel’fa severo-zapadnoi chasti Beringova morya (Ichthyocene of the Upper Shelf of the Northwestern Part of the Bering Sea), Magadan: Izd. Severo-Vost. Nauch. Tsentra Dal’nevost. Otdel. Ross. Akad. Nauk, 2007.
Datsky, A.V. and Maznikova, O.A., Biological features of common fish species in Olyutorsky-Navarin region and the adjacent areas of the Bering Sea: 3. Righteye flounders (Pleuronectidae), J. Ichthyol., 2017, vol. 57, no. 2, pp. 228–256. https://doi.org/10.1134/S0032945217020060
Datsky, A.V. and Samoilenko, V.V., Raw material base of aquatic biological resources in the Russian waters of the Bering Sea and its value, Vopr. Rybolovstva, 2021, vol. 22, no. 1, pp. 64–99. https://doi.org/10.36038/0234-2774-2021-22-1-64-99
De Robertis, A., Taylor, K., Wilson, C.D., and Farley, E.V., Abundance and distribution of Arctic cod (Boreogadus saida) and other pelagic fishes over the U.S. Continental Shelf of the Northern Bering and Chukchi Seas, Deep-Sea Res. II: Top. Stud. Oceanogr., 2017, vol. 135, pp. 51–65. https://doi.org/10.1016/j.dsr2.2016.03.002
Dumanskaya, I.O., Ledovye usloviya morei aziatskoi chasti Rossii (Ice Conditions of the Seas of the Asian Part of Russia), Moscow; Obninsk, IG-SOTsIN, 2017.
Eisner, L.B., Zuenko, Y.I., Basyuk, E.O., et al., Environmental impacts on walleye pollock (Gadus chalcogrammus) distribution across the Bering Sea shelf, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vols. 181–182, Article 104881. https://doi.org/10.1016/j.dsr2.2020.104881
Eschmeyer, W.N., Herald, E.S., and Hammann, H., A Field Guide to Pacific Coast Fishes of North America, Boston: Houghton Mifflin Co., 1983.
Fadeev, N.S., Bering Sea, in Biologicheskie resursy Tikhogo okeana (Biological Resources of the Pacific Ocean), Moscow: Nauka, 1986, pp. 389–405.
Fadeev, N.S., Spravochnik po biologii i promyslu ryb severnoi chasti Tikhogo okeana (Handbook of Biology and Fisheries of the North Pacific), Vladivostok: Izd. TINRO-tsentr, 2005.
Farley, E., Cieciel, K., Vollenweider, J., et al., Arctic Integrated Ecosystem Survey Cruise Report to the Arctic Integrated Research Program, Anchorage: NPRB, 2017.
Farley, E.V.Jr., Murphy, J.M., Cieciel, K., et al., Response of pink salmon to climate warming in the northern Bering Sea, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vol. 177, Article 104803. https://doi.org/10.1016/j.dsr2.2020.104830
Fey, D.P. and Weslawski, J.M., Age, growth rate, and otolith growth of polar cod (Boreogadus saida) in two fjords of Svalbard, Kongsfjorden and Rijpfjorden, Oceanologia, 2017, vol. 59, no. 4. https://doi.org/10.1016/j.oceano.2017.03.011
Frost, K.J. and Lowry, L.F., Demersal fishes and invertebrates trawled in the northeastern Chukchi and western Beaufort seas, 1976–77, in NOAA Tech. Rept. NMPS SSRF, no. 764, U.S. Department of Commerce; National Oceanic and Atmospheric Administration; National Marine Fisheries Service, 1983.
Glebov, I.I., Nadtochii, V.A., Savin, A.B., et al., Results of comprehensive studies in the East Siberian Sea in August 2015, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2016, vol. 186, pp. 81–92. https://doi.org/10.26428/1606-9919-2016-186-81-92
Gorbatenko, K.M. and Kiyashko, S.I., Zooplankton composition and trophic status of aquatic species in the Laptev and East Siberian Seas, Oceanology (Engl. Transl.), 2019, vol. 59, no. 6, pp. 893–902. https://doi.org/10.1134/S0001437019060055
Grebmeier, J.M., Cooper, L.W., Ashjian, C.A., et al., Pacific Marine Arctic Regional Synthesis (PacMARS) Final Report, Anchorage: NPRB, 2015.
Gritsai, E.V., Interannual variability of spatial differentiation and functioning of the East Bering Sea walleye pollock populations, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2003, vol. 133, pp. 80–93.
Helser, T.E., Colman, J.R., Anderl, D.M., and Kastelle, C.R., Growth dynamics of saffron cod (Eleginus gracilis) and Arctic cod (Boreogadus saida) in the northern Bering and Chukchi Seas, Deep-Sea Res. II: Top. Stud. Oceanogr., 2017, vol. 135, pp. 66–77. https://doi.org/10.1016/j.dsr2.2015.12.009
ICES, Report of the workshop on age reading of Greenland halibut 2 (WKARGH2) in ICES CM 2016/SSGIEOM:16, Reykjavik: ICES, 2017. https://doi.org/10.2960/S.v41.m1
Karamushko, L.I., Bioenergetika i rost morskikh vidov ryb Arktiki (Bioenergy and Growth of Arctic Marine Fish Species), Apatity: Izd-vo Fed. Issled. Tsentra Kol. Nauch. Tsentra Ross. Akad. Nauk, 2020. https://doi.org/10.37614/978.5.91137.438.9
Kim, Sen Tok, Features of biology and abundance of Pacific cod in the waters of the western coast of Sakhalin and the South Kuril Islands, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1998, vol. 124, pp. 212−235.
Kovalev, S.A. and Filippova, S.G., “Arktika-2010”—a new step of Russia, Rybn. Khoz-vo, 2010, no. 5, pp. 16–17.
Kuznetsov, M.Yu., Basyuk, E.O., Chul’chekov, D.N., and Syrovatkin, E.V., Distribution and hydrological conditions of walleye pollock in the northwestern part of the Bering Sea in the summer-autumn period, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2013, vol. 174, pp. 104–124.
Kuznetsova, N.A. and Gorbatenko, K.M., Feeding of Arctic cod, walleye pollock and other pelagic fish and their food supply in the Chukchi Sea in August–September 2019, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2021, vol. 201, no. 4, pp. 765–783. https://doi.org/10.26428/1606-9919-2021-201-765-783
Lamb, A. and Edgell, P., Coastal Fishes of the Pacific Northwest, Madeira Park: Harbour Publ., 1986.
Lear, W.H. and Ritt, N.K., Otolith age validation of Greenland halibut (Reinhardtius hippoglossoides), J. Fish. Res. Board Can., 1975, vol. 32, no. 2, pp. 289–292. https://doi.org/10.1139/f75-027
Loboda, S.V. and Zhigalin, A.Yu., Results of studies of Pacific herring in the northwestern part of the Bering Sea in 2010–2015, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2017, vol. 188, pp. 125–139. https://doi.org/10.26428/1606-9919-2017-188-125-139
Logerwell, E.A., Busby, M., Mier, K.L., et al., The effect of oceanographic variability on the distribution of larval fishes of the northern Bering and Chukchi seas, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vol. 177, Article 104784. https://doi.org/10.1016/j.dsr2.2020.104784
Makrofauna pelagiali zapadnoi chasti Beringova morya: tablitsy vstrechaemosti, chislennosti i biomassy, 1982–2009 gg. (Pelagial Macrofauna of the Western Bering Sea: Tables of Occurrence, Abundance, and Biomass, 1982–2009), Vladivostok: Izd. TINRO-tsentr, 2012.
Manteifel’, B.P., Saika i ee promysel (Saika and Its Fishery), Arkhangelsk: OGIZ, 1943.
Matley, J.K., Fisk, A.T., and Dick, T.A., The foraging ecology of Arctic cod (Boreogadus saida) during open water (July-August) in Allen Bay, Arctic Canada, Mar. Biol. (Berlin), 2013, vol. 160, no. 11, pp. 2993–3004. https://doi.org/10.1007/s00227-013-2289-2
Mecklenburg, C.W., Mecklenburg, T.A., Sheiko, B.A., and Steinke, D., Pacific Arctic Marine Fishes. Conservation of Arctic Flora and Fauna, Akureyri: CAFF Internat. Secretariat, 2016.
Moiseev, A.R. and Grinfel’dt, Yu.S., General information, in Ekosistemy Beringova proliva i faktory antropogennogo vozdeistviya (Ecosystems of the Bering Strait and Factors of Anthropogenic Impact), Moscow: World Wildlife Fund, 2019.
Moiseev, P.A., Cod and flounder of the Far Eastern seas, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1953, vol. 40.
Morrow, J.E., Freshwater Fishes of Alaska, Anchorage: Alaska Northwest Publ. Co., 1980.
Moskalenko, B.K., On the biology of polar cod (Arctic cod) Boreogadus saida (Lepechin), Vopr. Ikhtiol., 1964, vol. 4, no. 3 (32), pp. 433–443.
Munk, K.M., Maximum ages of groundfishes in waters off Alaska and British Columbia and consideration of age determination, Alaska Fish. Res. Bull., 2001, vol. 8, no. 1, pp. 12–21.
Musienko, L.N., Reproduction and development of fishes in the Bering Sea, Tr. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 1970, vol. 70, pp. 166–225.
Nikolaev, A.V., Kuznetsov, M.Yu., and Syrovatkin, E.V., Acoustic studies of Arctic cod (Boreogadus saida) in the Russian waters of the Bering and Chukchi Seas in 1999–2007, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2008, vol. 155, pp. 131–143.
Nishio, S., Sasaki, H., Waga, H., and Yamamura, O., Effects of the timing of sea ice retreat on demersal fish assemblages in the northern Bering and Chukchi Seas, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vols. 181–182, Article 104910. https://doi.org/10.1016/j.dsr2.2020.104910
Novikov, N.P., Snytko, V.A., and Dolgikh, I.P., Promyslovyi atlas Dal’nevostochnykh morei (Fishery Atlas of the Far Eastern Seas), Vladivostok: Izd. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1992.
Orlov, A.M., Benzik, A.N., Vedishcheva, E.V., et al., Fishery research in the Chukchi Sea on the R/V Professor Levanidov in August 2019: some preliminary results, Tr. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 2019, vol. 178, pp. 206–220. https://doi.org/10.36038/2307-3497-2019-178-206-220
Orlov, A.M., Savin, A.B., Gorbatenko, K.M., et al., Biological research in the Russian Far Eastern and Arctic seas in the transarctic expedition of VNIRO, Tr. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 2020, vol. 181, pp. 102–143. https://doi.org/10.36038/2307-3497-2020-181-102-143
Orlov, A.M., Rybakov, M.O., Vedishcheva, E.V., et al., Walleye pollock Gadus chalcogrammus, a species with continuous range from the Norwegian Sea to Korea, Japan, and California: new records from the Siberian Arctic, J. Mar. Sci. Eng., 2021, vol. 9, no. 10, Article 1141. https://doi.org/10.3390/jmse9101141
Plotnikov, V.V., Vakul’skaya, N.M., Mezentseva, L.I., et al., Variability of ice conditions in the Chukchi Sea and their relationship with the Arctic oscillation, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2020, vol. 20, no. 1, pp. 155–167. https://doi.org/10.26428/1606-9919-2020-200-155-167
Safronov, S.N., Pacific navaga, in Biologicheskie resursy Tikhogo okeana (Biological Resources of the Pacific Ocean), Moscow: Nauka, 1986, pp. 201–212.
Savin, A.B., Arctic cod (Boreogadus saida, Gadidae) of the Chukchi Sea and adjacent waters, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2021, vol. 201, no. 4, pp. 810–832. https://doi.org/10.26428/1606-9919-2021-201-810-832
Semenenko, L.I., On the local populations of the Pacific saffron cod and the prospects for its fishing in the northern part of the range, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1965, vol. 59, pp. 136–144.
Sheiko, B.A. and Fedorov, V.V., Class Cephalaspidomorphi – Lampreys. Class Chondrichthyes – Cartilaginous fishes. Class Holocephali – Holocephalians. Class Osteichthyes – Bony fishes, in Katalog pozvonochnykh zhivotnykh Kamchatki i sopredel’nykh morskikh akvatorii (Catalog of Vertebrate Animals of Kamchatka and Adjacent Marine Areas), Petropavlovsk-Kamchatskii: Kamchat. Pechat. Dvor, 2000, pp. 7–69.
Shleinik, V.N., Biological basis for the rational use of Arctic cod in the Barents Sea, Extended Abstract of Cand. Sci. (Biol.) Dissertation, Russ. Fed. Res. Inst. of Fisheries and Oceanography, Moscow, 1979.
Shuntov, V.P., Volkov, A.F., Temnykh, O.S., and Dulepova, E.P., Mintai v ekosistemakh dal’nevostochnykh morei (Walleye Pollock in Ecosystems of Far East Seas), Vladivostok: Vladivostok: Izd. Tikhook. Inst. Rybn. Khoz. Okeanogr., 1993.
Siddon, E.C., Zador, S.G., and Hunt, G.L., Ecological responses to climate perturbations and minimal sea ice in the northern Bering Sea, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vols. 181–182, Article 104914. https://doi.org/10.1016/j.dsr2.2020.104914
Sirenko, B.I. and Gagaev, S.Yu., Unusual abundance of macrobenthos and Pacific invaders in the Chukchi Sea, Biol. Morya, 2007, vol. 3, no. 6, pp. 399–407.
Slabinskii, A.M. and Figurkin, A.L., Structure of the plankton community in the southern part of the Chukchi Sea in summer, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2014, vol. 178, pp. 135–147. https://doi.org/10.26428/1606-9919-2014-178-135-147
Stepanenko, M.A., Interannual variability of spatial differentiation of pollock Theragra chalcogramma and cod Gadus macrocephalus of the Bering Sea, Vopr. Ikhtiol., 1997, vol. 37, no. 1, pp. 19–26.
Stepanenko, M.A. and Gritsai, E.V., State of resources, spatial differentiation and reproduction of walleye pollock in the northern and eastern parts of the Bering Sea, Izv. Tikhook. Inst. Rybn. Khoz. Okeanogr., 2016, vol. 185, pp. 16–30. https://doi.org/10.26428/1606-9919-2016-185-16-30
Stepanenko, M.A. and Gritsai, E.V., Interannual variability of environmental conditions and spatial differentiation of walleye pollock in the Bering Sea, Tr. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 2018, vol. 174, pp. 6–20.
Stevenson, D.E. and Lauth, R.R., Bottom trawl surveys in the northern Bering Sea indicate recent shifts in the distribution of marine species, Polar Biol., 2019, vol. 42, no. 2, pp. 407–421. https://doi.org/10.1007/s00300-018-2431-1
Udintsev, G.B., Boichenko, I.G., and Kanaev, V.F., Relief of the bottom of the Bering Sea, Tr. Inst. Okeanol. Akad. Nauk SSSR, 1959, vol. 29, pp. 17–64.
Velikanov, A.Ya., Far Eastern capelin: distribution, biology, biomass dynamics, problems and prospects of commercial development, Vopr. Rybolovstva, 2018, vol. 19, no. 3, pp. 30–326.
Vershinin, V.G., On the biology and current state of cod stocks in the northern part of the Bering Sea, in Biologicheskie resursy Arktiki i Antarktiki (Biological Resources of the Arctic and Antarctic), Moscow: Nauka, 1987, pp. 207–224.
Vinnikov, A.V., Terent’ev, D.A., and Batanov, R.L., Biological substantiation of establishing a commercial measure for Pacific cod in the Far Eastern fishery basin, in Tikhookeanskaya treska dal’nevostochnykh vod Rossii (Pacific Cod of Russian Far East Waters), Moscow: Izd. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 2013, pp. 263–266.
Witherell, D., Groundfish of the Bering Sea and Aleutian Islands Area: Species Profiles, Anchorage: NPFM. Counc., 2000.
Wolotira, R.J.Jr., Sample, T.M., and Morin, M., Demersal Fish and Shelf Fish Resources of Norton Sound, the Southeastern Chukchi Sea, and Adjacent Waters in the Baseline Year 1976, Seattle: NMFS, 1977.
Yasumiishi, E.M., Cieciel, K., Andrews, A.G., et al., Climate-related changes in the biomass and distribution of small pelagic fishes in the eastern Bering Sea during late summer, 2002–2018, Deep-Sea Res. II: Top. Stud. Oceanogr., 2020, vols. 181–182, Article 104907. https://doi.org/10.1016/j.dsr2.2020.104907
Zgurovskii, K.A. and Khen, G.V., Distribution of shrimp aggregations depending on the hydrological conditions of the northwestern part of the Bering Sea, in Promyslovo-biologicheskie issledovaniya morskikh bespozvonochnykh (Commercial and Biological Research of Marine Invertebrates), Moscow: Izd. Vseross. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 1988, pp. 98–107.
Zolotov, A.O., Comparison of age estimates for yellowfin (Limanda aspera Pallas) and northern bilinear (Lepidopsetta polyxystra Orr et Matareze) soles in the western part of the Bering Sea and the eastern coast of Kamchatka based on scales and otoliths, Issled. Vod. Biol. Resur. Kamchatki i Severo-Zap. Cha. Tikh. Okeana, 2006, no. 8, pp. 198–206.
ACKNOWLEDGMENTS
The authors are sincerely grateful to all members of the Pacific, Kamchatka, and Polar branches and Central Office of the Russian Federal Research Institute of Fisheries and Oceanography, who were involved in scientific research works in the Chukchi and Bering seas in different years, as well as to members of ship crews for active assistance in collecting the primary material used in this article.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interests.
Additional information
Translated by D. Zabolotny
Rights and permissions
About this article

Cite this article
Datsky, A.V., Vedishcheva, E.V. & Trofimova, A.O. Features of the Biology of Mass Fish Species in Russian Waters of the Chukchi Sea. 1. Commercial Fish Biomass. Family Gadidae. J. Ichthyol. 62, 560–585 (2022). https://doi.org/10.1134/S0032945222040051
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0032945222040051