
Abstract—
The paper presents characteristics of the species composition, spatial distribution, and trophic interactions of ichthyoplankton and zooplankton in the spring–summer period 2017 dependent on intensity of the warming-up of the upper sea layer against the background of transition in a system of the surface currents from the winter-to summer-type of the circulation. In April–May, successful spawning of temperate-water fish species was prolonged due to low values of the sea surface temperature (8–9°С) on the shelf of Crimean Peninsula between Kerch Strait and the cape Sarych. The maximum abundance of Sprattus sprattus eggs was 162.8 egg/m2 and larvae –116 ind/m2. Warm-water species occurred only in the western sector from Herakleian Peninsula to the Cape Tarkhankut, where the water temperature was at its maximum and peaked at 14–15°С. In June–July the delayed summer hydrological season hampered the warming-up of the upper quasi-homogenous layer, which thickness varied from 1–5 to 10–15 m. Notwithstanding the large number of species in the ichthyoplankton, maximum abundance did not exceed 50 egg/m2 and 14 ind/m2 for larvae. The proportion of larvae of temperate-water fish species averaged 32% and reached 40% in deep water areas. Juvenile stages of copepods of the cold-water complex were dominant in the larval fish diet both in the spring and summer seasons.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Two groups of the marine (saltwater) fish have been established in the Black Sea that authentically reproduce in seas of the temperate zone with well-defined seasonality; that is, the temperate-water Atlantic–boreal relics and warm-water south-boreal subtropical and tropical species (Rass, 1965). Thermohaline structure and the water dynamics significantly determine species composition and abundance of ichthyoplankton in different areas of the sea (Klimova et al., 2019). In the Black Sea, spawning of the warm-water fish species is observed from April to October at the sea surface temperature of 14–27°С and is confined to the upper quasi-homogenous sea layer. For temperate-water species, mass spawning is commonly associated with winter hydrological season and is limited to December–February. Existence of the cold intermediate layer during the summer hydrological season in the Black Sea allows for the presence in ichthyoplankton of eggs and larvae of a number of fish species characterized by multiple spawning, such as sprat Sprattus sprattus, Merlangius merlangus, and Trisopterus luscus, all year round (Aleev, 1958; Rass, 1965; Alonso-Fernández, 2011; Klimova and Podrezova, 2018). The global warming is particularly apparent in marine ecosystems of the temperate zone, featuring sharp seasonal differences, which further determine specific characteristics of phenology of hydrobionts (Visser and Both, 2005; Edwards and Richardson, 2004). Changes in phenology of spawning in both the thermophilic (warm-water) and cryophilic (cold-water) fish have been shown to lead to simultaneous occurrence of larvae of the warm-water and temperate-water complexes in ichthyoplankton (Visser and Both, 2005; Edwards and Richardson, 2004; Auth et al., 2017). Currently, mass spawning of sprat in the Black Sea is observed in November (Klimova and Podrezova, 2018). The spawning season of thermophilic fish species has extended. While the successful spawning of the abundant warm-water European anchovy Engraulis encrasicolus was previously confined to June–September, currently the anchovy larvae occur in the plankton from May to October (Dekhnik, 1970; Klimova et al., 2019).
The goal of the present work is to study peculiarities of the species diversity, spatial distribution of ichthyoplankton, and trophic relationship of fish larvae in planktonic community of the Black Sea off the Crimean Peninsula during the spring and early summer hydrological seasons of 2017 depending on the hydrological regime and state of the fish forage base.
MATERIALS AND METHODS
The material was collected on April 22–May 6 and June 14–July 6, 2017 in the shelf and deep water areas of the Black Sea off the coast of Crimean peninsula during cruises 94 and 95 of R/V Professor Vodyanitskii. Ichthyoplankton was collected with a Bogorov–Rass reverse cone-shaped net (BR 80/113, the mouth opening 0.5 m2, mesh 400 µm). Zooplankton was sampled using the Judey net (diameter 38 cm, mesh 140 µm) concurrently with the ichthyoplankton catches. Ichthyoplankton and zooplankton were collected in the total vertical haul regime. The sampling was done in the layer from the surface to the sea floor at shallow-water stations and down to a boundary of the hydrogen sulfide zone determined based on isopycnal δt = 16.2 relative units in the deep water areas based on Sea-Bird STDplus data. Samples were preserved in 4% formaldehyde solution for further laboratory processing. The total of 159 ichthyoplankton samples (59 in cruise 94 and 100 in cruise 95) and 40 zooplankton samples were collected during two cruises.
Ichthyoplankton was identified using the methods described earlier (Dekhnik, 1973; Russell, 1976). Species names of hydrobionts are given according to a World Register of Marine Species (WoRMS, 2019). Diversity indices were calculated (Simpson, 1949; Pielou, 1966; Odum, 1983; Shannon and Weaver, 1949). Abundance is presented below 1 m2 of the sea surface for ichthyoplankton and in 1 m3 for zooplankton. Nutrition of fish larvae was analyzed using the Duka and Sinyukova method (1976). Size–weight relationships known for the Black Sea species were used to change over from the size characteristics of zooplankton to the biomass units (Petipa, 1957; Chislenko, 1968). The analysis of hydrological regime and surface circulation employed the cruises’ materials (Artamonov et al., 2019), satellite infrared (IR) images of the sea surface, and altimetry data for the period from mid-April to mid-July 2017 (Morskoi portal, 2019), as well as the monitoring data of monthly temperature and salinity in Sevastopol coastal waters (Troshchenko et al., 2019).
RESULTS
Characteristics of Hydrological Regime
In 2016–2017 the winter hydrological season commenced in December 2016 and continued to mid-April of 2017 as shown by the analysis of accessible information in regard to thermal regime of surface water in the Black Sea water area adjoining the Crimean Peninsula over the period, preceding the present survey. Slow rise in sea surface temperature (SST) with gradients >0.1°С/day (3–3.5°С/mo.), corresponding to the spring hydrological seasons, was observed between mid-April and end of May. Intense warming-up of the water upper layer, which caused SST to increase sharply with gradients ~0.2°С/day and characterized the onset of the summer hydrological season, began as late as in the beginning June.
The spring hydrological season is characterized by transition of surface current systems from the winter- to spring-type circulation. According to geostrophic calculations and instrumental data (Artamonov et al., 2019), in April–May, main elements of surface water circulation were the northern periphery of the Black Sea Rim Current (BRC) and two quasi-permanent anticyclonic eddy swirls (ACE) characteristic of this time of the year; specifically, the Sevastopol (SevACE) above the Large Topographic Trough (LTT) west of the Crimean Peninsula and Crimean (CrACE) south of Cape Meganom (Ivanov and Belokopytov, 2011; Oguz et al., 1993; Korotaev et al., 2003; Klimova et al., 2019).
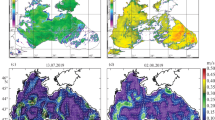
With the measured SST values, ranging from 8–8.5 to 14.5–15.5°С, the BRC waters proved to be the most warmed-up in the western sector (Cape Tarkhankut–Herakleian Peninsula) and SevACE, whereas the least warmed-up waters were those of the coastal shelf zone between Kerch Strait and cape Sarych (Artamonov et al., 2019) (Fig. 1a). Surface waters of the northern periphery of BRC in the central (cape Sarych –Alushta) and eastern (Alushta–Kerch Strait) sectors were warmed up to as low as 11.0–11.5°С. Thermal structure of the water column was characterized by quasi-homogenous vertical distribution of the winter type at the early stage of the spring warming-up.

Distribution of (a), (b) temperature and (c), (d) salinity across the Black Sea water surface layer off the Crimean peninsula based on the 2017 survey data: (a), (b) April; (c), (d) June (after Artamonov et al., 2019 modified); (⚫) stations.
In zones influenced by BRC stream waters, the upper sea layer of up to 5–10 m was occupied by interlayer of relatively warm waters (>10°С) with the maximum vertical gradients of temperature. Cold intermediate layer (CIL) in its classical type (Ivanov and Belokopytov, 2011) was observed across the larger portion of the examined water area in the form of local pools of increased and decreased temperature values in its core. In this setting, regions with the minimum temperature values (7–7.3°С) corresponded to the maximum penetration of the CIL core (down to 60–80 m) and were associated with zones of possible convergence along outer margin of the BRC stream above continental slope and northern periphery of SevACE.
Salinity of the surface waters varied between 18.2 and 18.7‰. The larger portion of the water area had salinity at 18.2–18.4‰, while maximum values (up to 18.5–18.7‰) were recorded in nearshore zone southwesterly of Feodosiya Gulf and in southwestern part of the polygon in the northern periphery of the western core of the main cyclonic gyre (MCG) (Fig. 1b).
Period of conducting the summer survey (June 14–July 6, 2017) coincided with an initial phase of the summer hydrological season. Seasonal weakening of BRC and its active meandering along the continental slope, as well as intense warming-up of the sea surface layer led to a substantial transformation in circulation and thermohaline structure of waters. While the most significant dynamic formations recorded during the spring survey sustained, activation of cyclonic meander of BRC in the northwestern part of the sea along the western slope of LTT led to the SevACE strengthening and its displacement to the trough’s east. Simultaneous northerly displacement of the western MCG core was noted. In the eastern sector, the CrACE displaced southwesterly along the shelf boundaries toward the Alushta area.
The most warmed-up (to 25–26°С) waters occupied southeastern part of the studied water area. The waters less warmed-up (to 23–24°С) corresponded to confinement zone of SevACE. Nearshore zone, stretching along the entire Crimean Peninsula from Kerch Strait to Tarkhankut Peninsula was characterized by a drop in SST (down to 16–18°С) values. Minimum SST values (11–12°С) were observed off cape Sarych (Artamonov et al., 2019) against this background (Fig. 1c). The SST distribution similar to this is indicative of quasi-permanent coastal upwelling (CU). The analysis of IR images allowed us to reveal an important element of the CU retention that is, active mushroom-shaped structure termed vortex dipole (VD). The VD originated from the northeastern part of the sea to the south of Taman Peninsula on June 5–6 as a result of sharp increase of northeastern wind of the bora-type. The VD performed transitional move from the nearshore zone toward the open sea along the continental slope and could be observed on satellite IR images until its entire transformation on the beam of the Yalta Bay on June 25–26. The VD development and transformation process followed the classical pattern in line with the records of analogical phenomena (Ginzburg and Fedorov, 1984; Grishin and Subbotin, 1992). At its rear end, VD provided for compensation rise of subsurface waters in the nearshore zone between Anapa and Kerch Strait and strengthened the CU along the Crimean southeast shoreline. In this setting, SST in VD jet differed from the surrounding waters by 3–4°С.
Delayed start of the summer hydrological season resulted in the thickness of the upper quasi-homogenous layer (UQL) not exceeding 10–15 m during the survey. In this setting, values were observed to be the maximum in the zones of SevACE and CrACE, while the minimum (1–5 m) corresponded to the zones of the water rise in CU across the larger portion of the Crimean nearshore. Parameters of the CIL, such as thickness, depth of core penetration, and minimum temperature values likewise corresponded to positions of the main dynamic formations; specifically, the highest temperatures (7.8−8°С) and minimum parameters of thickness and depth of the core occurrence (40–45 m) were associated with zones of the water rise in the eastern and western cyclonic gyres; while minimum temperatures (7–7.3°С) and maximum depths of the core occurrence (85–90 m) were recorded in the form of local spots along the convergence zone at the outer BRC margin and in the SevACE and CrACE regions.
No significant changes occurred in the field of distribution of surface salinity compared with the spring survey. Major portion of the examined water area was occupied by the waters of the Black Sea upper water masses with a salinity level of 18–18.7‰ (Ivanov and Belokopytov, 2011). The lowest salinity (17.8–17.9‰) was observed in the northwestern part of the polygon southwest of Cape Tarkhankut and in the eastern part of the polygon in the form of a chain of plumes of freshened surface water along axis of the VD transition (Fig. 1d).
Species Composition and Distribution of Ichthyoplankton
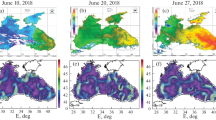
Six species of eggs and fish larvae represented ichthyoplankton during the spring survey (April 22–May 6, 2017). The mean numbers were 13.7 egg/m2 and 14.4 ind/m2 for larvae. Eggs of the spring-spawning Scophthalmus maeoticus were encountered occasionally exclusively on the shelf no deeper than 150 m between the Kara-Dag beam and Cape Tarkhankut with the eggs reported alive only near Cape Tarkhankut. Larvae of the warm-water Syngnathus schmidti and Pomatoschistus minutus were reported only in the western sector on the beam of the Kalamita Bay and nearshore station at Balaklava. Ichthyoplankton was missing from one station at Cape Tarkhankut, as well as deep-water sites of the southwestern and central sectors of the survey (Fig. 2a).

Distribution of ichthyoplankton off the Crimean peninsula in (a) April–May and (b) June–July 2017: (◼) empty samples, (⚪) eggs, egg/m2; (⚫) larvae, ind/m2.
Spawning of the temperate-water S. sprattus, Merlangius merlangus, and T. luscus continued (Klimova and Podrezova, 2018). The mean numbers constituted 12.2 egg/m2 for S. sprattus and did not exceed 1.1 egg/m2 for M.merlangus (Table 1). Larvae of S. sprattus and T. luscus of different ages occurred in all study areas. The sprat was dominant in the ichthyoplankton; its eggs and larvae accounted for 89 and 96%, respectively. The maximum numbers of S. sprattus eggs were 162.8 egg/m2 and 116 ind/m2 for larvae at stations at the southern shore of Crimea (Fig. 2a).
Eggs and larvae of 24 fish species from 18 families represented ichthyoplankton during the period of the summer survey (June 14–July 6, 2017). The mean numbers were 11.9 egg/m2 and 2 ind/m2 for larvae (Table 1, Fig. 2b). Eggs of 13 species from 13 families were represented in the ichthyoplankton. The warm-water E.encrasicolus eggs were dominant; it accounted for 82.5% of the total number of eggs in all encountered species; Trachurus mediterraneus accounted for approximately 9%; and eggs of other fish species occurred singly. Share of eggs of temperate-water fish species (S. sprattus and M. merlangus) totaled 3.4%. Single sprat eggs were reported above depths of 86–900 m; M. merlangus eggs were singly encountered down to 100-m isobath.
The larvae were represented by 16 fish species from 10 families (Table 1). Larvae of the warm-water species from demersal eggs in total did not exceed 27%. Larvae of S. schmidti accounted for 20%. Share of larvae from pelagic eggs added up to 21%; E. encrasicolus (13%) was dominant. Share of larvae of temperate-water fishes, such as S. sprattus, M. merlangus, and T. luscus, summed up to 32%. Sprat larvae were dominant (28%); the larvae were recorded above depths of 70–2000 m. M. merlangus occurred exclusively on the continental slope on the beam of Sevastopol and Yalta; while T. luscus larvae were found above the depths of 1400–1800 m on the beam of the Kara-Dag and Feodosiya Gulf.
Abundance and species diversity of ichthyoplankton changed as distance between the stations and shore increased. Ichthyoplankton was represented by 20 fish species (13 species of eggs and 10 species of larvae) on the shelf of the Crimean Peninsula (depths ≤200 m). The mean numbers were 25 egg/m2 and 3.2 ind/m2 for larvae. Eggs of warm-water fish species were dominant in the samples (94.4%). In this setting, E. encrasicolus eggs accounted for 77%; T. mediterraneus eggs accounted for 12%; and eggs of the remaining nine species were encountered occasionally. Eggs of temperate-water fishes were represented by S. sprattus and M. merlangus. Larvae of warm-water fish species were dominant; among them, larvae from the demersal and pelagic eggs accounted for 46.7 and 36.3%, respectively. Larvae of temperate-water fishes were represented only by sprat; their proportion in the total number of larvae on the shelf was sufficiently high (16%). The mean abundance and number of species in ichthyoplankton were comparable with the parameters of July 1989–1992, when the number of species had fallen short of 15, while the number of eggs and larvae had not exceeded 22 egg/m2 and 3 ind/m2, respectively, during the period of mass spawning of warm-water fish species on the shelf of the Crimean Peninsula. The latter period, however, was characterized by degradation of the planktonic community due to introduction and large-scale development of the ctenophore Mnemiopsis leidyi (Klimova, 1998).
In the deep water areas (depths >200 m) ichthyoplankton was collected mostly in the last days of June and early July. There were 12 species of fish eggs and larvae identified in these areas. The mean numbers were 7.3 egg/m2 and 1.4 ind/m2 for larvae. Warm-water species were represented by eggs of E. encrasicolus (94%) and T. mediterraneus (4%), while temperate-water species by eggs of S. sprattus and M. merlangus (2%). Larvae of warm-water species were dominant among larvae in samples (60%). Larvae from demersal eggs accounted for 51%; they all appear to have been drifted away from the nearshore shelf water areas. The proportion of larvae of the temperate-water fish species constituted 40% (S. sprattus accounted for 35% among them).
Diversity indices were calculated to evaluate a state of ichthyoplankton complexes above various depths (Simpson, 1949; Odum, 1983; Pielou, 1966; Shannon and Weaver, 1949). Shelf of the Crimean Peninsula featured the most favorable conditions for embryonic and postembryonic fish development (Table 2). In the deep-water areas, the number of species in ichthyoplankton was half of the number on the shelf; species richness index declined from 13.84 to 10.63. The intense spawning of the European anchovy was observed in the area; proportion of its eggs in samples exceeded 95% (dominance index 0.72), which contributed to a decrease in parameters of the species diversity and evenness indices.
Zooplankton and Fish Larvae Diet
The work reports data on zooplankton as forage base for fish, for which the term fodder zooplankton was proposed by Kusmorskaya (1950). Herewith, the use of this term implies all zooplankton species consumed by fish. The fodder zooplankton was represented in April–May by the common Black Sea species. Six copepod species were detected, including Calanus euxinus, Pseudocalanus elongatus, Acartia clausi, Paracalanus parvus, Oithona similis, and recent invader O. davisae; singular occurrence of cladoceran Pleopis polyphemoides was noted. Larvae of the benthic animals were represented by mollusks (Bivalvia and Gastropoda), polychaetes (Polychaeta), and cirripedians (Cirripedia). All of them were few in number in plankton. Meroplankton (bentic larvae), appendicularian Oikopleura (Vexillaria) dioica and chaetoghaths Parasagitta setosa were consistently present; whereas rotifers of the genus Synchaeta were also present in the samples but extremely rare. Quantitative characteristics of zooplankton during the spring–summer period of 2017 is shown in Table 3.
Copepods accounted for 93.2% of the abundance and 70.3% of biomass of fodder zooplankton in the inshore waters. Their contributions changed little on the continental slope in terms of abundance (94.0%), but dropped to 60.7% in terms of biomass. In the offshore waters, proportion of copepods in fodder zooplankton based on abundance remained almost unchanged (93.5%), but appreciably decreased based on biomass (34.8%) for the benefit of large-size chaetoghaths present in samples. Abundance of copepods declined above the continental slope and remained at the same level in offshore waters. The biomass of copepods was minimal on the continental slope and increased in the inshore and offshore waters. Cold-water species, such as С. euxinus and P. elongatus, were dominant among copepods with the first being dominant by biomass and the second by abundance across the entire examined water area.
Species composition of zooplankton changed little in offshore in June–July 2017. The warm-water copepod Centropages ponticus had emerged, which was encountered singly and, similar to cyclopoid O. davisae, accounted for fractions of a percentage of copepod abundance. Similar to spring, a considerable portion of fodder zooplankton included copepods with 95.5% of abundance and 44% of biomass. Cold-water species, such as С. euxinus and P. elongatus, remained dominant among copepods with the first species being dominant based on biomass and the second by abundance as it was in spring. Chaetoghaths (large specimens predominantly) accounted for more than half of the fodder zooplankton biomass (55.8%). Overall, abundance and biomass of copepods and all the fodder zooplankton in the offshore proved to be lower in summer compared with spring. Low abundance of small-size fraction of crustacean zooplankton in summer is noted. Thus, Calanoida nauplii accounted for 30.2% of copepod abundance in spring, whereas their portion dropped to 19% in spring.
Sprat larvae dominated in ichthyoplankton samples in April–May 2017. Data that had been published in regard to size–weight composition and peculiarities of sprat diet on the shelf and deep-water part of the Black Sea at Crimean Peninsula dates to March–April 1986 (Tkach et al., 1991). The larvae were allocated by us to three size groups, analogous to the ones reported in the aforementioned work to perform the comparative analysis of the size composition and diet of larval sprat in spring 2017 and 1986 (Table 4). In the size groups, mean length of larvae proved to be greater in April–May 2017 compared with spring 1986. According to Dekhnik (1973), sprat commonly finishes spawning in March–April. In 1986, proportion of the larvae of younger age group (3.5–10 mm) was only 5%. In April–May 2017, intense spawning of the sprat continued, which was evidenced by a high proportion (80%) of larvae with TL ≤ 10 mm in the total number of the larvae.
In spring 2017, the main prey items of sprat larvae included juvenile stages of copepods, primarily Calanoida, which were observed thoroughly digested in larval sprat guts. The number of copepods in one gut varied within a range of 1–21 individuals with their size being 0.15–0.35 mm. In addition, eggs of hydrobionts were encountered in food bolus in small quantities (1–4 eggs per one gut). In size groups II and III, proportion of larvae with food items was larger than in 1986 (Table 4). Low proportion of larvae with food items in size group I in 2017 can be attributed to the large number in samples of larvae with TL ≤ 4.3 mm, which mouth opening had not been yet developed according to data by Dekhnik (1973).
Species composition of diet items of larval sprats was slightly different in 1986 and 2017. They fed on copepods mostly, such as Oithona nana, P. elongatus, Calanus helgolanticus (presently С. euxinus), and P. parvus, at juvenile and adult stages, with the nauplii of О. nana accounting for between 14.3 and 75% of the number of all the consumed items in March–April 1986 (Tkach et al., 1991). Species from the family Oithonidae were not detected in larval sprat guts in 2017, which is associated with disappearance from plankton of the abundant year-round species О. nana (Zagorodnyaya and Skryabin, 1995), while O. davisae introduced in the 2000s (Zagorodnyaya, 2002) became abundant during the warm season of the year (Temnykh et al., 2012). During the spring survey, O. davisae was detected on a few stations only; its abundance was extremely low and averaged 0.33 and 1.70 ind/m3 in inshore and offshore waters, respectively.
In June–July 2017 the larval fish diet was examined in larvae (70 ind) of 12 species. Sprat larvae (19 ind) were represented by a wide size range (TL 1.7–20 mm); their mean length was comparable with the one of April–May 2017, but greater than in spring 1986 (Table 4). The larval sprat guts were noted to contain copepods (Calanoida) at juvenile stages from 1 to 7 ind per one gut. Among 18 larvae of Syngnathus schmidti captured in the deep-water part of the sea (depth >200 m), two individuals (TL 8.2 and 20 mm) did not have any food in the guts; other larvae (TL 14.2–20 mm) had copepods Calanoida 0.15–0.58 mm in size identified in bolus; 1–10 ind per one gut. Six individuals of copepods (Calanoida) of 0.2–0.4 mm were identified in the gut of S. schmidti TL 14.6 mm in size caught at shallower depths. Diplodus annularis (five ind of TL 3.5–4.4 mm) caught on the shelf (<200 m) were found to have copepods at juvenile stages 0.12–0.35 mm in size and occasional eggs of hydrobionts. Along with copepods (Calanoida), ten larvae of the family Gobiidae (TL 3.7–10 mm) were noted to have in the bolus the cladocera and eggs of hydrobionts; no food items were detected in six larvae of TL 2.6–5 mm. Larvae of the remaining fish were represented by single individuals, which qualitative composition of the gut contents did not essentially differ from larvae of other species dominant in the samples and was largely represented by copepods at juvenile stages.
DISCUSSION
The first survey (April 22.04–May 6, 2017) corresponded to the end of spawning of the temperate-water and onset of the spawning season of warm-water fish species; period of the active spawning of warm-water fish species coincided with the second survey (June 14–July 6, 2017). The analysis of series of sequential satellite IR images (Morskoi portal, 2019) revealed the dynamics of the surface water warming-up at various stages of hydrological seasons and served to explain the established peculiarities of species composition and spatial distribution of ichthyoplankton.
The SST variability ranged 8–12°С at the early stage of the spring survey: values of SST were observed to be the minimum (8–9°С) in the nearshore zone of Crimea from Kerch Strait to Herakleian Peninsula and maximum (11–12°С) on the water area of Kalamita Bay. Warming-up of the upper water layers led to spatial restructuring of the SST field by the completion of the survey. Under the SST range of variability of 9–15°С, the most warmed-up waters proved to be in Kalamita Bay and in BCR waters in the western sector with an abrupt cut-off southwesterly toward the sea from the Tarkhankut Cape, while coldest waters were recorded in the eastern sector.
During the spring ichthyoplankton survey, the warm-water species were detected only in the most warmed-up waters on the beam of Kalamita Bay. The shelf and deep-water sea areas of the Crimean Peninsula between Herakleian Peninsula and Kerch Strait displayed the water temperature favorable for spawning of the temperate-water fish. Eggs and larvae of sprat were dominant in the samples. The maximum abundance of sprat was 162.8 egg/m2 and 116 ind/m2 for larvae on the beam of the southern coast of Crimea. Large proportion of larvae of the young age group (80%) confirmed the successful mass spawning of sprat.
State of the forage base in the studied area supported a survival of larval sprats, which was confirmed by a small proportion of larvae with empty guts. Copepods were dominant in inshore and offshore waters (>90% of the fodder zooplankton abundance). Among them, the dominance belonged to cold-water species, such as C. euxinus and P. elongatus, which generally form the basis of the sprat diet. Proportion of small-size fraction in the fodder zooplankton exceeded 50%. The larval sprat guts predominantly contained copepods at juvenile stages, primarily Calanoida, which nauplii accounted for 30% of the small-size fraction of fodder zooplankton. In offshore waters, a comparatively high biomass of big predatory chaetoghaths did not have a significant effect on abundance of crustacean zooplankton, inasmuch as chaetoghaths are actively consumed by adult sprat. According to Chayanova (1958), proportion of chaetoghaths in sprat guts approached 90%.
Period of the summer survey corresponded to a transition from the spring to summer hydrological season and was accompanied by a sharp rise of SST and formation of the summer-type thermal stratification (Artamonov et al., 2019). Spatial–temporal changes in the SST distribution pattern were conditioned by the regional inertia in water column warming and effects of the various quasi-permanent (ACE and MCG) and non-stationary (CU and VD) dynamic structures. In mid-July, the SST variability ranged 18−22°С with the values being the maximum on the larger part of the studied water area, including BRC, SevACE, and CrACE and minimum in the narrow nearshore zone of Crimea, stretching from Kerch Strait to Herakleian Peninsula, as a consequential effect of “latent” CU. The SST had increased to 19–26°С by early July. Waters from the BRC system in eastern part of the polygon proved to be the most warmed-up with the SST rising to 23–24°С in the nearshore zone of Crimea. Northern peripheries of the western and eastern cores of MCG were characterized by the minimum SST values (19–20°С). During the survey period, maximum thickness of CIL did not exceed 10–15 m, while the minimum thickness was reduced to 1–5 m in upwelling zones.
Spawning of warm-water fish species and their early developmental stages are exclusively confined to UQL, where the water temperature is favorable for embryonic and postembryonic fish development. In the ichthyoplankton, 24 fish species were identified in the shelf and deep-water areas of the Black Sea off the Crimean Peninsula. The species composition of the ichthyoplankton corresponded to the onset of the summer spawning season with the mean number of eggs (11.9 egg/m2) and larvae (2 ind/m2) being comparatively low. Thus, in the second ten-day period of June 2016 with the same species composition mean number of eggs was twice and of larvae five times higher (Ignat’ev et al., 2017). The deep-water areas of the sea predominantly featured larvae of the temperate-water (40%) and warm-water fish species from demersal eggs (51%) largely caught in the northeastern sector. Drift of the warm-water fish species larvae (dwellers of the Black Sea nearshore water areas) toward the deep water study areas appear to have occurred due to the active mushroom-shaped structure (VD), which formed during the first ten days of June and was migrating along the continental slope toward the open sea until the complete transformation on the beam of Yalta during the third ten days of June. Its migration was accompanied by compensation rise of subsurface water in the nearshore zone between Anapa and Kerch Strait and strengthening of coastal upwelling along the southeastern shore of Crimea, which led to a decrease in the numbers of eggs and larvae of warm-water fish species, while sprat proceeded with the successful spawning.
The number of larvae proved to be appreciably lower during summer period than in spring, which appears to be related to specifics of the sea thermal regime due to the delayed summer hydrological season. June 2017 was cold for embryonic development of warm-water fish. Cold-water species, such as С. euxinus and P. elongatus, continued to dominate in fodder zooplankton. Species of the cold-water complex are known to take precedence during the spring period and, in some instances, in the summer as well (Greze and Fedorina, 1979), exactly which was observed by us in June 2017. Apparently, the sprat was already finishing spawning and was migrating to the nearshore sea water areas for the summer fattening (Aleev, 1958), inasmuch as its eggs and larvae occurred singly. In the absence of predators and feeding competitors, proportion of big chaetoghaths grew up to 55.8% in biomass of fodder zooplankton biomass. When compared with the spring season, abundance and biomass of crustacean zooplankton in deep water areas decreased by a factor of 1.5, while nauplii of copepods accounted for as little as 19% of the Calanoida abundance. Reduction of small-size fraction in fodder zooplankton appears to have had a negative effect on survival of the larvae of nearshore fish species drifted off toward the deep water areas.
CONCLUSIONS
The SST variability ranged 8–15°С in April–May 2017. The most warmed-up proved to be the waters of Kalamita Bay and BRC in the western sector with the submeridional boundary starting from the Cape Tarkhankut. The colder waters were reported from the nearshore zone of Crimea, from the Herakleian Peninsula to Kerch Strait. Active spawning of temperate-water sprat was observed under low water temperature along the Crimean Peninsula. The warm-water ichthyoplankton species, such as eggs of S. maeoticus and larvae of Gobiidae spp. and S. schmidti, occurred only in the western sector, where the sea surface temperature reached 15°С.
In June–early July 2017 the upper quasi-homogenous layer did not exceed 10–15 m on average due to a delayed summer hydrological season and was reduced to 1–5 m in the nearshore zones exposed to “latent” upwelling. The mean number of eggs and larvae of the warm-water fish species was considerably lower compared with June 2016. Proportion of larvae of the temperate-water fish species averaged 32% and reached 40% in the deep water areas. During the first ten days of June 2017 abrupt strengthening of northeastern wind led to the formation of active mushroom-shape structure (VD) in the nearshore zone southerly of the Taman Peninsula and drift of the nearshore fish species larvae from demersal eggs away to the deep water areas, where their proportion reached 51% in the total larval fish abundance.
No significant differences have been established in qualitative diet composition among various larval fish species on the shelf and deep water sea area between the spring and summer 2017 study periods. Copepods at juvenile stages of Calanoida were dominant in larval fish diet during both seasons. Adult copepods and the ones at older stages were dominant in zooplankton. Simultaneous presence in samples of a wide size range of larvae of the temperate-water and warm-water fish species; occurrence of larvae with different feeding types; and, in addition, low proportion of larvae with empty guts indirectly suggest conditions favorable for their survival. This confirms the state of the forage base, inasmuch as, notwithstanding a decline in quantitative values of fodder zooplankton in the deep water sea areas, copepods with a relatively high share of nauplii were noted to be dominant in the plankton.
REFERENCES
Aleev, Yu.G., Biology and economic role of the Black Sea sprat Sprattus sprattus phalericus (Risso), Tr. Sevastop. Biol. Stn., 1958, vol. 10, pp. 90–107.
Alonso-Fernández, A., Bioenergetics approach to fish reproductive potential: case of Trisopterus luscus (Teleostei) on the Galician Shelf (NW Iberian Peninsula), PhD Thesis, Vigo: Univ. of Vigo, 2011.
Artamonov, Yu.V., Fedirko, A.V., Skripaleva, E.A., et al., Water structure in the zone of the Main Black Sea Current in spring and summer 2017 (94th and 95th cruises of R/V Professor Vodyanitsky), Ekol. Bezop. Pribrezhnoi Shel’fovoi Zon Morya, 2019, no. 1, pp. 16–28. https://doi.org/10.22449/2413-5577-2019-1-16-28
Auth, T.D., Daly, E.A., Brodeur, R.D., and Fisher, J.L., Phenological and distributional shifts in ichthyoplankton associated with recent warming in the northeast Pacific Ocean, Global Change Biol., 2017, vol. 24, no. 1, pp. 259–272. https://doi.org/10.1111/gcb.13872
Black Sea data archive, Maritime portal, 2019. http://dvs.net.ru/mp/data/main_ru.shtml. Accessed September, 2019.
Chayanova, L.A., Feeding of the Black Sea sprat, Tr. VNIRO, 1958, vol. 36, pp. 106–127.
Chislenko, L.L., Nomogrammy dlya opredeleniya vesa organizmov po razmeram i forme tela (morskoi zoobentos i plankton) (Nomograms for Determination of Weight of Aquatic Organisms by Size and Body Shape (Marine Zoobenthos and Plankton)), Leningrad: Nauka, 1968.
Dekhnik, T.V., Distribution and abundance of pelagic eggs and larvae, in Razmnozhenie i ekologiya massovykh ryb Chernogo morya na rannikh stadiyakh ontogeneza (Reproduction and Ecology of Black Sea Mass Fishes at the Early Ontogenesis Stages), Kiev: Naukova Dumka, 1970, pp. 59–88.
Dekhnik, T.V., Ikhtioplankton Chernogo morya (Ichthyoplankton of the Black Sea), Kiev: Naukova Dumka, 1973.
Duka, L.A. and Sinyukova, V.I., Rukovodstvo po izucheniyu pitaniya lichinok i mal’kov morskikh ryb v estestvennykh i eksperimental’nykh usloviyakh (Handbook on Analysis of Feeding of Larvae and Juveniles of Marine Fishes in Natural and Experimental Conditions), Kiev: Naukova Dumka, 1976.
Edwards, M. and Richardson, A.J., Impact of climate change on marine pelagic phenology and trophic mismatch, Nature, 2004, vol. 430, pp. 881–884. https://doi.org/10.1038/nature02808
Ginzburg, A.I. and Fedorov, K.N., Umbrella-like currents in the ocean according to satellite survey data, Issled. Zemli Kosm., 1984, no. 3, pp. 18–26.
Greze, V.N. and Fedorina, A.I., Abundance and biomass of zooplankton, in Osnovy biologicheskoi produktivnosti Chernogo morya (Fundamentals of Biological Productivity of the Black Sea), Greze, V.N., Ed., Kiev: Naukova Dumka, 1979, pp. 157–164.
Grishin, G.A. and Subbotin, A.A., Analysis of the eddy dipole in the Black Sea using satellite data and on-board ship measurements, Issled. Zemli Kosm., 1992, no. 5, pp. 56–64.
Ignat’ev, S.M., Melnikov, V.V., Klimova, T.N., et al., Macro- and ichthyoplankton of the Crimean coastal area in summer 2016, Sist. Kontr. Okruzh. Sredy, 2017, no. 8 (28), pp. 93–101.
Ivanov, V.A. and Belokopytov, V.N., Okeanografiya Chernogo morya (Oceanography of the Black Sea), Sevastopol: EKOSI-Gidrofizika, 2011.
Klimova, T.N., Dynamics of species composition and numbers of ichthyoplankton of the Black Sea in the area of the Crimea in the summer periods of 1988–1992, J. Ichthyol., 1998, vol. 38, no. 8, pp. 645–651.
Klimova, T.N. and Podrezova, P.S., Seasonal distribution of the Black Sea ichthyoplankton near the Crimean Peninsula, Reg. Stud. Mar. Sci., 2018, vol. 24, pp. 260–269. .https://doi.org/10.1016/j.rsma.2018.08.013
Klimova, T.N., Subbotin, A.A., Melnikov, V.V., et al., Spatial distribution of ichthyoplankton near the Crimean Peninsula in the summer spawning season of 2013, Morsk. Biol. Zh., 2019, vol. 4, no. 1, pp. 63–80. https://doi.org/10.21072/mbj.2019.04.1.06
Korotaev, G., Oguz, T., Nikiforov, A., and Koblinsky, C., Seasonal, interannual, and mesoscale variability of the Black Sea upper layer circulation derived from altimeter data, J. Geophys. Res.: Oceans, 2003, vol. 108, no. 4. https://doi.org/10.1029/2002JC001508
Kusmorskaya, A.P., Zooplankton of the Black Sea, Tr. Azovo-Chernomorsk. Nauchno-Issled. Inst. Rybn. Khoz. Okeanogr., 1950, no. 14, pp. 177–214.
Odum, E., Basic Ecology, New York: CBS College, 1983, vol. 2.
Oguz, T., Latun, V.S., Latif, M.A., et al., Circulation in the surface and intermediate layers of the Black Sea, Deep Sea Res., Part I, 1993, vol. 40, no. 8, pp. 1597–1612. https://doi.org/10.1016/0967-0637(93)90018-X
Petipa, T.S., On the average weight of the main forms of zooplankton from the Black Sea, Tr. Sevastop. Biol. Stn., 1957, vol. 9, pp. 39–57.
Pielou, E.C., Species-diversity and pattern- diversity in the study of ecological succession, J. Theor. Biol., 1966, vol. 10, pp. 370–383.
Rass, T.S., Rybnye resursy evropeiskikh morei SSSR i vozmozhnosti ikh popolneniya akklimatizatsiei (Fish Resources of European Seas of USSR and Their Possible Naturalization), Moscow: Nauka, 1965.
Russell, F.S., The Eggs and Planktonic Stages of British Marine Fishes, London: Academic, 1976.
Shannon, C.E. and Weaver, W., A Mathematical Theory of Communication, Urbana: Univ. of Illinois Press, 1949.
Simpson, E.H., Measurement of diversity, Nature, 1949, vol. 163, p. 688. https://doi.org/10.1038/163688a0
Temnykh, A.V., Tokarev, Yu.N., Melnikov, V.V., and Zagorodnyaya, Yu.A., Daily dynamics and vertical distribution of pelagic Copepoda in offshore waters in front of the South-Western Crimea (the Black Sea) in autumn 2010, Morsk. Ekol. Zh., 2012, vol. 11, no. 2, pp. 75–84.
Tkach, A.V., Glushchenko, T.I., and Zagorodnyaya, Yu.A., Feeding of larvae and fry of the Black Sea sprat Sprattus sprattus phalericus (Risso) in early spring, Ekol. Morya, 1991, no. 37, pp. 56–61.
Troshchenko, O.A., Subbotin, A.A., and Eremin, I.Yu., Variability of the main limiting environmental factors during growing of bivalve mollusks on a farm in the Sevastopol region, Uch. Zap. Kaliningr. Fed. Univ., Ser. Geogr. Geol., 2019, vol. 5, no. 71-2, pp. 308–321.
Visser, M.E. and Both, C., Shifts in phenology due to global climate change: the need for a yardstick, Proc. R. Soc. London, Ser. B, 2005, vol. 272, no. 1581, pp. 2561–2569. https://doi.org/10.1098/rspb.2005.3356
WoRMS, World register of marine species, 2019. http://www.marinespecies.org/index.php; https://doi.org/10.14284/170.
Zagorodnyaya, Yu.A., Oithona brevicornis in the Sevastopol Bay: accidental case or new invader in the Black Sea? Ekol. Morya, 2002, no. 61, p. 43.
Zagorodnyaya, Yu.A. and Skryabin, V.A., Current dynamics of zooplankton in the coastal regions of the Black Sea, in Issledovaniya shel’fovoi zony Azovo-Chernomorskogo basseina (Study of the Self Zone of the Azov-Black Sea Basin), Sevastopol: Morsk. Gidrofiz. Inst., Nats. Akad. Nauk Ukr., 1995, pp. 87–95.
Funding
This work was conducted as a part of the state assignment to the Institute of Biology of the Southern Seas of the Russian Academy of Sciences: “Regularities of Formation and Anthropogenic Transformation of Biodiversity and Bioresources of the Sea of Azov–Black Sea Basin and Other Areas of the World Ocean”, project no. АААА-А18-118020890074-2 and “Study on the Mechanisms of the Production Processes Management in Biotechnological Complexes Intended to Develop Scientific Foundations for Obtaining Biologically Active Substances of Technical Products of the Marine Genesis”, project no. АААА-А18-118021350003-6. The work was also supported by the grants of the Russian Foundation for Basic Research “Response of the Black Sea Pelagic System to Climate Change in the Region (Case Study on Jellyfish, Comb Jellies, and Small Pelagic Fish)”, project no. 18-44-920022 and “The Role of the Global and Local Factors in the Formation of Ichthyoplankton Communities in the Black Sea”, project no. 18-44-920007.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflicts of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by E. Kuznetsova
Rights and permissions
About this article

Cite this article
Klimova, T.N., Subbotin, A.A., Vdodovich, I.V. et al. Distribution of Ichthyoplankton in Relation to Specifics of Hydrological Regime off the Crimean coast (the Black Sea) in the Spring–Summer Season 2017. J. Ichthyol. 61, 259–269 (2021). https://doi.org/10.1134/S0032945221020077
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0032945221020077