
Abstract
The state of gonads of natural juvenile pink salmon Oncorhynhus gorbuscha, which was caught in five rivers during its downstream migration from spawning grounds, and hatchery juvenile pink salmon with different rearing periods, which was taken from 22 fish hatcheries in Sakhalin oblast, have been studied. The germ cells in the testes of all studied fish were represented by a few gonia. A single generation of oocytes of the previtellogenic period had already been formed in the ovaries of all females; the germ cells of earlier development stages (gonia and meiocytes) were hardly present.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
The gametogenesis of juvenile pink salmon Oncorhynhus gorbuscha (including gametogenesis during its rearing at fish hatcheries) has been studied in detail, although the main data on this issue were obtained mainly in the 1960s–1970s (Persov, 1960, 1965, 1966; Pakhomova and Hlevnaya, 1977; 1978; Zelennikov, 2003; Zelennikov et al., 2007). At the same time, fish farming has undergone radical changes in Sakhalin oblast, the only region in Russia where juvenile pink salmon are currently reproduced. Over the past 25 years, all 52 hatcheries were either built for the first time or rebuilt in a modern form and acquired new technical characteristics. The purpose of our research was to find out how the new growing conditions influenced the gametogenesis of juvenile pink salmon.
MATERIALS AND METHODS
Juvenile pink salmon was collected in May–June 2016 and 2017 in five rivers and at all 22 Sakhalin salmon hatcheries (SHs) where it was reproduced during these years. Natural juveniles were caught at night during their downstream migration; hatchery fish were taken from the first batch at each hatchery, immediately before their release into the wild. Fifty specimens were fixed in Bouin or Serra’s fluids in each case.
The histological processing was carried out according to traditional methods (Mikodina et al., 2009); 80 to 120 serial transverse sections of both gonads were prepared from each specimen and were then stained with iron hematoxylin by Heidenhain. A total of 375 fish gonads were prepared and treated. To assess the state of ovaries, we determined the area of the transverse section of both gonads of each female on three sections that were taken at an interval of ten serial sections, followed by the calculation of the number of germ cells of all development periods and measurement of the major and minor diameter of the ten largest oocytes in each gonad (the half sum of the gonads was assumed to be their diameter). In total, this analysis covered preparations of 225 individuals.
The significance of differences in the average values was determined using Student’s t-test. The strength of the relationship between the total amount of heat received by juveniles and fish weight, as well as between the fish weight and parameters of gonad state, was estimated using the Pearson’s pairwise correlation coefficient.
RESULTS AND DISCUSSION
The weight of natural juvenile pink salmon was significantly lower than that of the hatchery one, which is noticeable even during the analysis of relatively small female samples (Table 1): on average, approximately 200 vs. 300–500 mg (at two hatcheries (Bukhta Olya and Kitovy), small batches of pink salmon were grown in the production experiment regime to an average weight of ~900 mg.
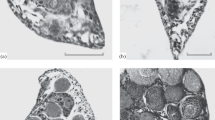
The gonads of all males represented poorly differentiated glands that had an average of five to ten gonia on the transverse sections. All females (both those that were caught in the river and those taken at hatcheries) (Fig. 1) had previtellogenic oocytes with a similar size and state in their ovaries; cytoplasm of their oocytes contained elements of the circumnuclear complex (Fig. 1a). Germ cells of the earlier state (gonia and oocytes in early meiosis prophase) occurred as an exception (see Fig. 1c). Among fry from natural spawning, we found only three individuals with an average of five or more of these cells on the ovary sections; there were no such individuals among hatchery fish. The completed formation of the single generation of previtellogenic oocytes is indicated by the fact that the average number of cells of this period per section was similar in wild and hatchery fry (9.6–12.6, with the only exception of 14.6) and did not change with increase in the body weight. In contrast, an increase in the weight of fish consistently led to an increase in the volume of their ovaries (judging by their area on the transverse sections (r = 0.92)) and in the diameter of oocytes (r = 0.87). The area of gonads on sections and diameter of oocytes also consistently increased (r = 0.80 and r = 0.84, respectively) in fish at different hatcheries with increase in the sum of degree days accumulated by fry.

State of ovaries in females of salmon Oncorhynhus gorbuscha before their release from hatcheries: (a, b) a single generation of oocytes of the previtellogenic period was formed under (a) standard and (b) long-term rearing conditions; (c, d) rare (c) or numerous (d) oocytes in early meiosis prophase. Scale: 0.1 mm.
It is interesting to note that the state of gonads was almost the same in the microanatomical structure and composition of germ cells in natural and hatchery juveniles (both ordinary and those grown to a larger weight) (Figs. 1a, 1b). The larger fish differed only in larger oocytes and, as a consequence, larger ovaries. A female with a weight of 760 mg (Kitovy SH) was the only exception; its germ cell pool was only represented by gonia and oocytes in the early meiosis prophase mainly in the zygotene state (Fig. 1d). It is possible that it is these rarely occurring fish with a slow oogenesis rate that explain the detection of 3-year-old salmon spawners (Ivanov, 1996; Wagner and Stauffer, 1980).
To determine the dependence of the parameters of gonad state on the weight of fish, we analyzed three juvenile samples from the Lesnoy SH (10 days before the release and during the release) and Bukhta Olya SH with a weight of 270.9 (154–369), 396.0 (173–514), and 903.4 (297–1682) mg, respectively (Table 1). An increase in the body weight determined some (insignificant) trend towards an increase in the area of gonads on sections (r = 0.59, r = 0.63, and r = 0.47) and in the oocyte diameter (r = 0.41, r = 0.45, and r = 0.18). It can be assumed that this relationship, which has been recorded for juveniles of other salmon fishes (Zelennikov, 1997; Khristoforov and Murza, 1998), would be significant at a larger sample size.
Therefore, the state of gonads of juvenile pink salmon before its release from hatcheries providing modern bioengineering conditions for its rearing proved to be significantly standardized. Among all studied females, we did not identify any individual with a significant proportion of germ cells at early development periods (gonia and meiocytes). On the contrary, an older and actually single generation of oocytes of the previtellogenic period had already been formed in all females. The abundance of this generation was not recruited in the course of further fish growth, and only the growth of oocytes determined an increase in the volume of gonads in this period. Among natural juveniles, we found only three individuals with a significant proportion of gonia and meiocytes. However, it is quite possible that the proportion of these fish can be higher in years with less favorable temperature conditions (Zelennikov and Fedorov, 2005).
REFERENCES
Ivanov, O.A., A case of catch of two-year pink salmon Oncorhynchus gorbuscha in a sea in winter 1994–1995, Vopr. Ikhtiol., 1996, vol. 36, no. 5, pp. 716–720.
Khristoforov, O.L. and Murza, I.G., Reproductive function and its control in Atlantic salmon, in Atlanticheskii losos’ (The Atlantic Salmon), Kazakov, R.V., Ed., St. Petersburg: Nauka, 1998, pp. 75–126.
Mikodina, E.V., Sedova, M.A., and Chmilevskii, D.A., et al., Gistologiya dlya ikhtiologov: opyt i sovety (Histology for Ichthyologists: Experience and Recommendations), Moscow: VNIRO, 2009.
Pakhomova, N.A. and Khlevnaya, A.S., Effect of water temperature on gametogenesis of pink salmon during gender inversion, in Biologiya severnykh morei Evropeiskoi chasti SSSR (Biology of Northern Seas of European Part of USSR), Apatity: Kol’sk. Fil., Akad. Nauk SSSR, 1977, pp. 54–60.
Pakhomova, N.A. and Khlevnaya, A.S., Morphology of pink salmon ovaries in the marine period of life, in Voprosy funktsional’noi morfologii morskikh zhivotnykh (Functional Morphology of Marine Animals), Apatity: Kol’sk. Fil., Akad. Nauk SSSR, 1978, pp. 102–108.
Persov, G.M., Characteristics of the early ontogenesis of the sexual glands of pink salmon related with its use as naturalization object, Materialy soveshchaniya po voprosam rybovodstva (Proc. Conf. on Fishery Problems), Mileiko, I.V., Ed., Moscow: Rybnoe Khozyaistvo, 1960, pp. 86–92.
Persov, G.M., The re-differentiation of gonads of pink salmon (Oncorhynchus gorbuscha Walbaum) as normal development, Nauchn. Dokl. Vyssh. Shk., Biol. Nauki, 1965, no. 1, pp. 26–30.
Persov, G.M., Early gametogenesis in migratory salmons, in Vosproizvodstvo i akklimatizatsiya lososei v Barentsevom i Belom moryakh (Reproduction and Naturalization of Salmons in the Barents and White Seas), Moscow: Nauka, 1966, pp. 7–44.
Wagner, W.C. and Stauffer, Th.M., Three-year-old pink salmon in Lake Superior tributaries, Trans. Am. Fish. Soc., 1980, vol. 109, no. 4, pp. 458–460.
Zelennikov, O.V., The influence of water acidification on the ontogenesis of the reproductive system of fish in early ontogenesis, Extended Abstract of Cand. Sci. (Biol.) Dissertation, St. Petersburg: State Sci. Res. Inst. Lake River Fish., 1997.
Zelennikov, O.V., Comparative analysis of the state of the ovaries in juveniles of Pacific salmon related to the problem of the monocyclicity, Vopr. Ikhtiol., 2003, vol. 43, no. 4, pp. 490–498.
Zelennikov, O.V. and Fedorov, K.E., Early ontogenesis of pink salmon Oncorhynhus gorbuscha Walbaum in natural reproduction and farming in Sakhalin and Iturup islands, Vopr. Ikhtiol., 2005, vol. 45, no. 5, pp. 653–664.
Zelennikov, O.V., Sabanova, E.V., and Mishchenko, O.V., Effect of water acidification on the oogenesis of the pink salmon Oncorhynchus gorbuscha,J. Ichthyol., 2007, vol. 47, no. 3, pp. 254–257.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by D. Zabolotny
Rights and permissions
About this article

Cite this article
Zelennikov, O., Yurchak, M.I. Gametogenesis of Pacific Salmons: 1. The State of Gonads of Juvenile Pink Salmon Oncorhynhus gorbuscha under the Conditions of Its Natural and Hatchery Reproduction in Sakhalin Oblast. J. Ichthyol. 59, 966–969 (2019). https://doi.org/10.1134/S003294521906016X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S003294521906016X