
Abstract
A partial skeleton of a Mesozoic mammal has been discovered from the fourth member (tuffaceous sandstone) of the Lower Cretaceous Sinuiju Formation, Paekto-dong, Sinuiju city, North Phyongan Province, Democratic People’s Republic of Korea (DPRK). This specimen exhibits several characters common to all multituberculates. The femoral head is fully spherical and inflected, and sets off from the shaft by a long neck. The tibia has the pronounced, hook-like dorsolateral process in the proximal part and the medial and lateral articulating condyles on the tibia. The astragalus and the calcaneus are in juxtaposition, as preserved. The metatarsal V is offset from the cuboid. Hence this specimen belongs to multituberculate mammals. But this specimen has also the derived characters. The parafibula is not present. The calcaneocuboid facet is transverse with respect to the length of the calcaneus. In these regards, we suggest that the preserved postcranial skeleton of the new specimen has a mosaic of primitive and derived characters. This interpretation must be corroborated by additional mammal specimens from this locality.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
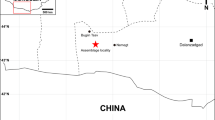
The Mesozoic lacustrine sedimentary rocks of the Sinuiju Formation from Paekto-dong, Sinuiju city are well known for a spectacular biota characterized by mixed terrestrial and lacustrine origins (e.g. fossil fish: Lycoptera davidii; conchostracans: Eosestheria middendorfii, Yanjiestheria kyongsangensis; fossil insect: Ephemeropsis trisetalis; ostracods: Cypridea concina, C. ritimensis, Darwinula tubiformis, Ziziphocypris costata; bivalves: Limnocyrena ovalis, L. sibirica, Ferganoconcha sp.; gastropods: Viviparus sp., Valvata sp., Corbicula sp.; plant fossils: Equisetites ushimarensis, Cladophlebis denticulata, Coniopteris burejensis, C. hymenophylloides, Ruffordia goepperti, Sphenopteris nitidula, Ginkgo sibirica, Ginkgoites jampolensis, Czekanowskia rigida) (Fig. 1) (Pak, 1996; Jon et al., 2019). Of them, an exciting Mesozoic mammal was first discovered in 2014 by the geological faculty from Paleontological Department, the Kim Il Sung University. The fossiliferous bed is confined to the fourth member, Sinuiju Formation of Paekto-dong, Sinuiju City.

Geographic map and outcrop showing the fossiol locality. (a) Map pertaining to the fossil locality; (b) photograph of the studied fossil locality.
GEOLOGICAL BACKGROUND AND METHOD
The Sinuiju Basin is distributed on the Amnokgang Fault Zone of the northeast. The Sinuiju Formation in Sinuiju City is distributed mainly in the Sinuiju basin, which is structurally separated by early Mesozoic or older strata along a southwest-northeast direction and extends over an area of about 250 km2. It comprises, in ascending order, the first to eighth members and the geologic age is the Early Cretaceous (Barremian–Aptian) (Won et al., 2019). The first member consists of sandstone and mudstone containing abundant invertebrate and plant fossils, overlying unconformably the Precambrian basement (Pak, 1996). The lower part of the first member consists of sandstone and conglomerates. The middle and upper parts comprise sandstone, siltstones and mudstone intercalated with mudstones. Invertebrate and plant fossils have been recovered from the mudstones. The second member is composed of volcanic rocks with sedimentary intercalations. The third member is distributed on Paekto-dong area, and consists of greyish-green sandstone, mudstone and siltstone. It is 200–300 m in thickness. The fourth to eighth members are covered with the Quaternary strata. But the lowermost part of the fourth member is distributed in Paekto-dong and Tosong-ri. The present mammal fossil comes from the tuffaceous sandstone, the lower part of the fourth member. It suffered diagenetic metamorphosis and preserved in lacustrine tuffaceous sandstone of Sinuiju Formation. Images of the pes and others are based on the outlines by microscope Leica tracing. The figured specimens are deposited in the collections of the Paleontological Department, Geological Faculty, Kim Il Sung University.
-
SYSTEMATIC PALAEONTOLOGY
-
CLASS MAMMALIA
-
SUBCLASS ALLOTHERIA
-
Order Multituberculata Cope, 1884
-
Multituberculata indet.
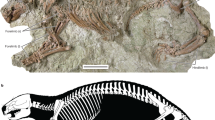
Description. Figures 1–7. The first Mesozoic mammal fossil from Paekto-dong, Sinuiju City is preserved on a slab of the tuffaceous Sandstone and the counter-slab is not preserved (Fig. 3, Table 1). In the fossil specimen, part of the vertebral column (lumbar and sacral vertebrae), pelvis, right hind limb, and left femur are preserved largely in articulation. But the lumbar ribs are absent.

Stratigraphic column measured at the Sinuiju Formation.

Mammal fossil from Sinuiju Formation (no. 2015.9-A).

Pelvic girdles (no. 2015.9-A). (a) Photo of pelvic girdles. The pelvic acetabular dorsal emargination is closed. The pubis and epipubis is not preserved. The inflected head of the femur is spherical and sets off from the shaft by a long neck. The acetabulum bears an inverted U-shaped region for the femoral head; (b) Line drawing of reconstruction of Fig. 4a.

Proximal part of the tibia and the fibular (no. 2015.9-A). (a) Photo of proximal part of the tibia and the fibular. The tibia has a proximolateral process. The patella is not preserved. The fibula doesn’t contacts the distal femur; (b) Line drawing of reconstruction of Fig. 5a.

Astragalus and calcaneus (no. 2015.9-A). (a) Photo of astragalus and calcaneus. The Navicular and mesocuneiform bones are obscure. The entocuneiform is elongate, and its joint with metatarsal 1 is offset anteriorly from the joint of the mesocuneiform and metatarsal 2 (arrow). Metatarsal 5 is offset from the cuboid. The calcaneus is well developed. The calcaneocuboid facet is transverse with respect to the length of the calcaneus; (b) Line drawing of reconstruction of Fig. 6a.

Metatarsals and phalanges (no. 2015.9-A). (a) Photo of metatarsals and phalanges. Pedal digit III is elongate relative to the digit IV; Line drawing of reconstruction of Fig. 7a. All scale bars are 1 mm.
Although there are several lumbar vertebrae but only one of the sacral vertebrae is preserved; their preservation is very poor.
The ilium, ischium and pubis are fused to form the pelvic girdle (Figs. 4a, 4b). The pelvic acetabular dorsal emargination is closed, with a smooth, unbroken and semicircular rim. The pubis and the epipubis are not preserved, but the pubic symphysis is preserved. The suture and internal aspect of the acetabulum is blocked from view by the head of the femur. The ilium is short and likely incomplete, and the preserved part is not rod-like. The anterior end of it is sharp and narrower than the rest of it. The posterior part of it shows the antero-dorsal margin of the acetabulum (Figs. 4a, 4b). The ischium is not well preserved, but what is preserved is distinctive, and shows a triangular outline. The acetabulum bears an inverted U-shaped region for the femoral head, though the part of it is preserved (Figs. 4a, 4b).
The femur shows relatively broader proximal and distal ends than the shaft. The length of it is relatively long, compared to the tibia and the fibula. The femoral head is spherical and inflected, and sets off from the shaft by a fairly long neck (Figs. 4a, 4b). The greater trochanter is robust and extends vertically from the shaft and beyond the femoral head. The distal end of the femur bears a clear epiphyseal suture between the shaft and the distal epiphysis (Figs. 5a, 5b). The two distal articulating condyles are formed by the epiphysis, and are separated by an intercondylar fossa in the posterior aspect of the femur (Figs. 5a, 5b). The femur lacks a distinct patellar groove, also there is no patella preserved, both suggestive of an absence of the patella.
The tibia has a proximolateral process, a primitive feature that is strongly developed and hooklike (Figs. 5a, 5b). The distal extremity of it shows a distinctive suture between the epiphysis and the diaphysis (shaft). The distal end of the tibial epiphysis has a medial malleolus, called the medial tibial condyle (sensu Krause and Jenkins 1983).
Apparently the fibula is not fused with the tibia in the specimen (Figs. 5a, 5b). It also does not contact the distal femur. But the lateral part of the fibular head bears a hooklike process for articulation with the hooklike process of the tibia. The fibula is narrower and shorter than the tibia.
Although the astragalus is not well preserved, it does not overlap over the calcaneus (lower ankle joint) (Figs. 6a, 6b). The navicular bone is incomplete (Figs. 6a, 6b). The entocuneiform is elongate, and it’s joint with metatarsal I is offset anteriorly from the joint of the mesocuneiform and metatarsal II (Figs. 6a, 6b). The ectocuneiform has an oblong and curved outline. The mesocuneiform is the shortest and smallest of the three cuneiform bones although poorly preserved.
The calcaneus is oblong outline. It has well-developed and laterally compressed tuber calcanei, a calcaneocuboid facet that is transverse and facing anteriorly, and a sustentacular facet that is medially facing (Figs. 6a, 6b). The base of the peroneal process is offset from anterior end of calcaneus. The sustentacular process forms an oblong outline (Figs. 4a, 4b). The cuboid is relatively large and is wider distally than proximally. The Metatarsal V is offset from the cuboid and not perfectly aligned laterally (Figa. 3a, 3b).
The metatarsals are elongated (Figs. 6a, 6b). Metatarsals I–IV have similar outline. The shafts are narrower and appear to be rod-like. The metatarsal V is very different. It is much stouter overall and broader distally. Only the proximal phalanges on digit III, and IV are complete (Figs. 7a, 7b). Based on the preserved proximal phalanges, they have similar shapes with their ends slightly wider than the rod-like shafts. The intermediate phalanges are incomplete on every digit. The terminal phalanges are very poor (Figs. 7a, 7b) (Table 1).
REMARKS
Now there are hundreds mammal skeletons from Jurassic and early Cretaceous of China, most of which consist of fragmentary jaws and teeth, which largely explain the intense emphasis that paleontologists place on the evolution of the mammalian dentition (Luo et al., 2011; Williamson et al., 2014). But in the present fossil from Paekto-dong, the dentition, the cranium is not preserved and the postcranial skeleton is partly preserved.
On the basis of the preserved postcranial skeletons, the present fossil is similar to multituberculates in many cruro-pedal characters.
Especially, the femoral head of our specimen is spherical, inflected and sets off from the shaft by a long neck (Figs. 4a, 4b), similar to the condition in multituberculates (Kielan-Jaworowska et al., 2004). The greater trochanter is long and directed proximally as in multituberculates, Zhangheotherium, Maotherium, and Henkelotherium (Krebs, 1991; Kielan-Jaworowska and Gambaryan, 1994; Chen and Luo, 2013). But cynodonts and Yanoconodon have the femoral head which are not spherical and have no neck; the greater trochanter has no vertical process (Luo et al., 2007; Chen et al., 2017).
The tibia possesses a large, hooklike proximolateral tuberosity (Figs. 5a, 5b). It is a plesiomorphic similarity to multituberculates, monotremes, morganucodontids, and docodonts (Luo et al., 2002; Martin, 2005; Luo et al., 2015; Meng et al., 2015). A large proximolateral tibial process is also present in many eutriconodonts, such as Jeholodens, Gobiconodon, and Spinolestes (Ji et al., 1999; Martin et al., 2015).
The shaft of the tibia in our specimen is relatively straight (Fig. 3), and is bowed medially as seen in the Late Cretaceous Chulsanbaatar, Nemegtbaatar, and Catopsbaatar (Kielan-Jaworowska and Gambaryan, 1994; Hurum and Kielan-Jaworowska, 2008). And the fibula has a straight shaft (Fig. 3), slightly bowed as those of several Cretaceous multituberculates (Kielan-Jaworowska and Gambaryan, 1994; Hurum and Kielan-Jaworowska, 2008).
The astragalus–calcaneus juxtaposition is a variable feature in multituberculates. In the most primitive known multituberculate with preserved ankle joint (Sinobaatar), the astragalus and the calcaneus are in juxtaposition, as preserved. The superposition of these two bones is not developed (Hu and Wang, 2002a, b). Ankle joint is preserved in Jurassic Rugosodon, in which the calcaneus and the astragalus are slightly dislocated, exposing the calcaneo-astragalar contact, which was interpreted to be capable of translational movement for the calcaneus relative to the astragalus (Yuan et al., 2013). In the Late Cretaceous djadochtatherian multituberculates from Mongolia, the astragalus is oriented obliquely relative to the calcaneal body, and does not overlap the calcaneus, according to the detailed restoration by Kielan-Jaworowska and Gambaryan (1994).
In the new specimen the entocuneiform is elongate, and it’s joint with metatarsal I is offset anteriorly from the joint of the mesocuneiform and metatarsal II (Figs. 6a, 6b) as in some multituberculates and eutherians.
In our specimen, the base of the peroneal process is offset from anterior end of calcaneus (Figs. 6a, 6b). In some multituberculates and Zhangheotherium, the peroneal process is also set off from the calcaneal body (Luo et al., 2003).
In the present specimen, Mt V is offset from cuboid (Figs. 6a, 6b) (such as multituberculates, symmetrodonts, and several eutriconodonts) (Kielan-Jaworowska and Gambaryan, 1994; Ji et al., 1999; Chen and Luo, 2013; Chen et al., 2017) in contrast to that in eutherian Asioryctes (Ji et al., 2002).
But the present specimen has also the derived characters.
Relative to the size of the pelvis and the acetabular diameter in our specimen, the pubic symphysis seems to be shorter than those of Zhangheotherium, Henkelotherium (Krebs, 1991) and Vincelestes (Rougier, 1993). Given the short sacral transverse processes and the deep pelvis (Figs. 4a, 4b), it is likely that the pelvis was narrow at the sacro-iliac joint and vertically deep, as hypothesized for Zalambdalestes (Kielan–Jaworowska, 1978), Ukhaatherium (Novacek et al., 1997), multituberculates (Kielan-Jaworowska and Gambaryan, 1994; Krause and Jenkins, 1983) and Eomaia (Ji et al., 2002).
Kielan-Jaworowska and Gambaryan (1994) suggested that parafibula might be characteristic of multituberculates as a whole. But in our specimen, the fibula apparently lacks contact with the femur and the parafibular (Figs. 5a, 5b), as the condition commonly encountered in eutherians.
The calcaneus is developed and similar to those of Asioryctes (Kielan-Jaworowska, 1977), Ukhaatherium (Horovitz, 2000), Zalambdalestes (Kielan-Jaworowska, 1978; Horovitz, 2000) and Eomaia (Ji et al., 2002) (Figs. 6a, 6b).
At the anterior end of the calcaneal body, the calcaneo-cuboid facet is transversely oriented, similar to the condition in Vincelestes (Rougier, 1993) and Cretaceous eutherians (Horovitz, 2000; Ji et al., 2002; Luo et al., 2003) (Figs. 6a, 6b).
Hence our analyses place the new fossil as a multituberculate, noting that it shares several cruro-pedal structure features with multituberculate mammals. However, our specimen has also some derived characters.
This interpretation must be corroborated by additional phylogenetic analyses that include a large swath of Mesozoic mammals and a broad sampling of dental and non-dental characters.
Multituberculates are generally regarded as the evolutionarily most successful and longest-lived (Middle Jurassic to late Eocene) clade of Mesozoic and early Paleogene mammals. First record of the Mesozoic mammal fossil from DPRK broadens the distribution and diversity of the Mesozoic multituberculate mammals. It also provides the new information on the pelvis, the hindlimb and pes, which in turn will help in exploring for other multituberculates in Sinuiju and elsewhere.
REFERENCES
Chen, M. and Luo, Z.-X., Postcranial skeleton of the Cretaceous mammal Akidolestes cifellii and its locomotor adaptations, J. Mamm. Evol., 2013, no. 20, pp. 159–189. https://doi.org/10.1007/s10914-012-9199-9
Chen, M., Luo, Z.-X., and Wilson., G.P., The postcranial skeleton of Yanoconodon allini from the Early Cretaceous of Hebei, China and its implications for locomotor adaptation in eutriconodontan mammals, J. Vertebr. Paleontol., 2017, e1315425. https://doi.org/10.1080/02724634.2017.1315425
Horovitz, I., The tarsus of Ukhaatherium nessovi (Eutheria Mammalia) from the Late Cretaceous of Mongolia: an appraisal of the evolution of the ankle in basal therians, J. Vertebr. Paleontol., 2000, no. 20, pp. 547–560.
Hu, Y.M. and Wang, Y.Q., Sinobataar gen. nov.: First multituberculate from Jehol Biota of Liaoning, Northern China, Chin. Sci. Bull., 2002a, no. 46, pp. 372–386.
Hu, Y.M. and Wang, Y.Q., Sinobataar gen. nov.: First multituberculate from Jehol Biota of Liaoning, Northern China, Chin. Sci. Bull., 2002b, no. 47, pp. 933–938.
Hurum, J.H. and Kielan-Jaworowska, Z., Postcranial skeleton of a Cretaceous multituberculate mammal Catopsbaatar, Acta Palaeontol. Pol., 2008, no. 53, pp. 545–566.
Jenkins, F.A., Jr. and Parrington, F.R., The postcranial skeletons of the Triassic mammals Eozostrodon, Megazostrodon and Erythrotherium, Philos. Trans. R. Soc. London, 1976, no. 273, pp. 387–431.
Ji, Q., Luo, Z.-X., and Ji, S.-A., A Chinese triconodont mammal and mosaic evolution of mammalian skeleton, Nature, 1999, no. 398, pp. 326–330.
Ji, Qiang, Luo, Zhe-Xi, Yuan, Chong-Xi, Wible, John, Zhang, Jianqing, and Georgi, J.A., The earliest known eutherian mammal, Nature, 2002, no, 416, pp. 816–821.
Jon, S.H., Won, C.G., So, K.S., and Nam, T.Y., New Mesozoic insect fossils from the Democratic People’s Republic of Korea, Cretaceous Res., 2019, no. 99, pp. 240–245.
Kielan-Jaworowska, Z., Evolution of the therian mammals in the Late Cretaceous of Asia, Part II: Postcranial skeleton in Kennalestes and Asioryctes, Palaeontol. Pol., 1977, no. 37, pp. 65–83.
Kielan-Jaworowska, Z., Evolution of the therian mammals in the Late Cretaceous of Asia, Part III: Postcranial skeleton in Zalambdalestidae, Palaeontol. Pol., 1978, no. 38, pp. 3–41.
Kielan-Jaworowska, Z. and Gambaryan, P.P., Postcranial anatomy and habits of Asian multituberculate mammals, Fossils Strata, 1994, no. 36, pp. 1–92.
Kielan-Jaworowska, Z., Cifelli, R.L., and Luo, Zhe-Xi, Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure, New York: Columbia Univ. Press, 2004.
Krause, D.W. and Jenkins, F.A., The postcranial skeleton of North American multituberculates, Bull. Mus. Comp. Zool., 1983, no. 150, pp. 199–246.
Krebs, B., Das Skelett von Henkelotherium guimarotae gen. et sp. nov. (Eupantotheria, Mammalia) ausdem Oberen Jura von Portugal, Berl. Geowiss. Abh., 1991, no. 133, pp. 1–110.
Luo, Z.-X., Kielan-Jaworowska, Z., and Cifelli R.L., In quest for a phylogeny of Mesozoic mammals, Acta Palaeontol. Pol., 2002, no. 47, pp. 1–78.
Luo, Z.X., Chen, P.J, Li, G., and Chen, M., A new eutriconodont mammal and evolutionary development in early mammals, Nature, 2007, no. 446, pp. 288–293.
Luo, Zhe-Xi, Ji, Qiang, Wible, J.R., and Yuan, Chongxi, An Early Cretaceous Tribosphenic Mammal and Metatherian Evolution, Science, 2003, no. 302, pp. 1934–1940.
Luo, Z.X, Yuan, C.X, Meng, Q.J., and Ji, Q., A Jurassic eutherian mammal and divergence of marsupials and placentals, Nature, 2011, no. 476, pp. 442–445.
Luo, Z.X., Meng, Q.J., Ji, Q., Liu, D., Zhang, Y.G., and Neander, A.I., Evolutionary development in basal mammaliaforms as revealed by a docodontan, Science, 2015, no. 347, pp. 760–764.
Luo, Z.X., Meng, Q.J., Liu, D., Zhang, Y.G., and Yuan, C.X., Cruro-pedal structure of the paulchoffatiid Rugosodon eurasiaticus and evolution of the multituberculate ankle, Palaeontol. Pol., 2016, no. 67, pp. 149–169.
Martin, T., Postcranial anatomy of Haldanodon exspectatus (Mammalia, Docodonta) from the Late Jurassic (Kimmeridgian) of Portugal, and its bearing for mammalian evolution, Zool. J. Linn. Soc., 2005, no. 145, pp. 219–248.
Martin, T., Marugan-Lobon, J., Vullo, R., Martin-Abad, H., Luo, Z.X., and Buscalioni, A.D., A Cretaceous eutriconodont and integument evolution of early mammals, Nature, 2015, vol. 526, no. 7573, pp. 380–384.
Meng, Q.J., Ji, Q., Zhang, Y.G., Liu, D., Grossnickle, D.M., and Luo, Z.X., An arboreal docodont from the Jurassic and mammaliaform ecological diversification, Science, 2015, no. 347, pp. 764–768.
Novacek, M.J., Rougier, G.W., Wible, J.R., McKenna, M.C., Dashzeveg, D., and Horovitz, I., Epipubic bones in eutherian mammals from the Late Cretaceous of Mongolia, Nature, 1997, no. 389, pp. 483–486.
Pak, I.S. and Kim, Y.N., Mesozoic Era, in Geology of Korea, Paek, R.J., Kang, H.G., and Jon, G.P., Eds., Pyongyang: Foreign Languages Books Publ. House, 1996, pp. 155–188.
Rougier, G.W., Vincelestes neuquenianus Bonaparte (Mammalia, Theria), unprimitivo mammifero del Cretacico Inferior de la Cuenca Neuqina, Thesis, Buenos Aires: Univ. Nac., 1993.
Szalay, F.S., Evolutionary History of the Marsupials and an Analysis of Osteological Characters, Cambridge: Cambridge Univ. Press, 1994.
Williamson, T.E., Brusatte, S.L., and Wilson, G.P., The origin and early evolution of Metatherian mammals: Cretaceous record, Zookeys, 2014, no. 465, pp. 1–76.
Won, C.G., So, K.S., and Jon, S.H., New evidence for a Cretaceous age for a Mesozoic nonmarine bivalve assemblage from Paektho-dong, Sinuiju City, The Democratic People’s Republic of Korea, Acta Geol. Sin. (Engl. Ed.), 2019, vol. 93, no. 6, pp. 1607–1613.
Yuan, C.X., Ji, Q., Meng, Q.J., Tabrum A.R., and Luo, Z.X., Earliest evolution of Multituberculate Mammala revealed by a new Jurassic Fossil, Science, 2013, no. 341, pp. 779–783.
ACKNOWLEDGMENTS
We thank Jo Song Dae for collaborative field work and various help during the fieldwork in Sinuiju in 2015–2016; Han Kum Sik for sharing ideas on early mammal research; Kim In Chan for improving the paper. The authors are extremely grateful to Jin Meng for his contribution to the study of the specimen. Our sincere gratitude is also offered to Marina A. Knorre for editing and anonymous reviewer for the very useful comments on the earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Won, C.G., So, K.S. & Jon, S.H. The First Known Mesozoic Mammal in the Democratic People’s Republic of Korea. Paleontol. J. 57, 826–832 (2023). https://doi.org/10.1134/S0031030123070122
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030123070122