
Abstract
On the basis of new extensive collections, made by the present authors, a group of unsegmented three-dimensional fossil remains from the Late Precambrian of Namibia and the southeastern White Sea area, including such genera as Protechiurus, Vendoglossa, and Vendoconularia, were restudied. It is established that the fossils belong to two morphologically close genera (Protechiurus and Vendoconularia), composing the new family Protechiuridae. It is possible that they represent the same genus or even species, but the incompleteness of the preservation of the Namibian member of the group prevents their integration. The fossils initially represented conical hollow elastic objects, hexagonal in the cross-section, which were open at the wide end and closed at the pointed end. The sculpture of their surface consisted of longitudinal ridges, both single and double, bounding the rows of the short transverse ridges, scalloped along one margin. Vendoconularia had wide plates which were attached to the cone outwardly along the longitudinal unpaired ridges. In general morphology and several details the fossils resemble the thecae of conulariids and anabaritids, known from Late Precambrian and Paleozoic and classified as the scyphozoan cnidarians. The main difference lays in the theca mineralization, which consists of phosphate in conulariids, carbonate in anabaritids, and is absent in the case of protechiurids. It is proposed that the protechiurids, which appeared in the fossil record first, may belong to the basal group of ancient scyphozoans, ancestral to the conulariids on the one hand, and to the anabaritids on the other.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Fossil remains resembling Paleozoic conulariids are rare in the assemblages of the Late Precambrian macrofossils of Namibia and Russia. Soft-body impressions of some of these macroorganisms are commonly preserved as casts, structures closed at one end and open at the other. In the least deformed specimens, the closed end terminates in a distinct point, and the open end is widely splayed. Preserved in many of these fossils is a sculpture consisting of several longitudinal and many transverse ridges demarking the surface.
One such specimen is in the Geological Museum of the Geological Survey of Namibia (Windhoek) associated with the numerous and diverse macrofossils collected over the past century from the Nama Group. The impression is strongly elongated, slightly flattened laterally and somewhat deformed (Pl. 1, fig. 6). Martin Glaessner described a relatively poorly preserved complex of features visible on this specimen, noted the presence of eight thin longitudinal ridges and weak transverse hatching, and suggested that these features alone were sufficient to distinguish this specimen as a separate species and genus of extinct organism, Protechiurus edmondsi. Glaessner assigned Protechiurus to echiuriids, a group of modern benthic multicellular animals or perhaps to the class of polychaetes (Glaessner, 1979).

Explanation of Plate 1
Protechiurus edmondsi Glaessner, 1979, Geological Museum of the Geological Survey of Namibia (GSN). Figures 1-5, 8a. Specimens coated with ammonium chloride. The white arrows indicate a doubled longitudinal ridge. Scale bars, 1 cm increments.
Fig. 1. Specimen GSN no. F673.
Fig. 2. Specimen GSN no. F661, the holotype of “Vendoglossa tuberculata” (Seilacher, 2007).
Fig. 3. Specimen GSN no. F653: (a) the natural cast, (b) the latex cast from a natural imprint.
Fig. 4. GSN specimen no. F669, side view.
Fig. 5. GSN no. F672, view from the apical (closed) end.
Fig. 6. Holotype GSN no. P5212165.
Fig. 7. Accumulation of small casts, specimen GSN no. F674.
Fig. 8. Specimen GSN no. F668, sandwiched between fragments of Pteridinium simplex, Aarhauser, Farm Aar, southern Namibia.
In 1993, Adolph Seilacher, while preparing the materials for his exhibition “Fossil Art”, made a cast of a large slab with imprints of typical Nama fossils collected from Farm Aar in rocks of the same age as the source of Protechiurus. There were several specimens of another fossil organism, along with numerous remains of Pteridinium simplex, on that slab. Seilacher later described these unusual fossils as Vendoglossa tuberculata (Seilacher, 2007). The casts that Seilacher prepared from his molds were designated as the holotype and the paratypes of this new species, since the original samples were left in place of their discovery on Farm Aar. According to Seilacher, Vendoglossa has an oval in cross-section, slightly flattened body with a surface covered by transverse rows of small tubercles (Pl. 1, fig. 2). Seilacher found it difficult to determine the position of this new taxon in the then current system of extinct organisms.
In 1997, M.A. Fedonkin discovered a unique fossil in terms of the completeness of its preservation in the deposits of the Late Vendian Ust-Pinega Formation of the southeastern White Sea area, Russia. In general shape and details it closely resembled Paleozoic conulariids. Subsequently, it was called Vendoconularia triradiata (Ivantsov and Fedonkin, 2002). The fossil represents a secondary flattened cast (inner mold) of the cone-shaped structure and two of its counterparts on the rock (Pl. 2, fig. 1). The presence of exclusively plastic deformation demonstrates that in life this organism was composed of an elastic material. Initially, one of the ends of the cone was closed, the other broadly open, and the cross-section was isometric. During the burial, sand penetrated through the wide open end into the cone, but filled only the part near the closed end, the basal end. The part of the cone, adjacent to the aperture, was not filled with sediment and was preserved as an incomplete imprint. Minor details of the sculpture and internal anatomy of Vendoconularia were preserved due to secondary ferrugination, which apparently was deposited in the densest clusters of the organic matter of the buried organism. There are 12 longitudinal ridges, 6 single and 6 double, arranged alternately on the surface of the cast. There are also 12 rows of transverse ridges, scalloped (tuberculate) along the outer margin. On one of the ends the transverse ridges merge in pairs and approach the longitudinal ridge in an alternating order. The symmetry of the cone was six fold—radial; i.e. it consisted of 6 identical elements, or modules. Ivantsov and Fedonkin, by comparing this fossil with the morphology of typical conulariids, suggested that the boundaries of the modules (sides of the theca) were represented by the doubled longitudinal ridges, or rather the groove that separated them (Ivantsov and Fedonkin, 2002). However, H. Van-Iten, specialist on conulariids and other colleagues, disagreed, pointing out that the module boundaries were likely single longitudinal ridges (Van Iten et al., 2005). The holotype also contains traces of internal structures represented by broad, thick, ferruginous longitudinal strips. It seems that the structures which formed these strips were likely located inside the cone in life, underlying each second one of the doubled longitudinal ridges. Only three such strips are preserved on the cast, and this means that the order of the general symmetry of the body of Vendoconularia was not greater than three-fold.

Explanation of Plate 2
Vendoconularia triradiata Ivantsov and Fedonkin, 2002. Figure 1b, 2-6 Specimens coated with ammonium chloride. Scale bars, 1 cm increments.
Fig. 1. The holotype PIN no. 4564/1025, the part of the specimen filled with sediment: 1a, 1d—imprint, dark pattern due to iron hydroxides, apparently, replacing areas with high content of organic matter (1d—the fragment with high magnification); (1b, 1c) the cast (1b—side view, 1c—a view from the apical end, white arrow indicates doubled longitudinal ridge).
Fig. 2. Specimen PIN no. 4564/1076. Specimen PIN no. 4564/1075.
Fig. 4. Specimen PIN no. 4564/1043.
Fig. 5. Specimen PIN no. 4564/1040.
Fig. 6. Small specimen PIN no. 4564/1077.
Thus, these three monospecific genera were represented by few specimens at the beginning of the 21st century—two represented by single specimens, while the originals of the type series of the third taxon were left in the field without an exact reference to the location. During the re-examination of this group, we carried out further collection of fossils in the type localities in Namibia and European North Russia.
CONICAL MACROFOSSILS OF NAMIBIA
The collection sites of the first discoveries of Precambrian macroorganisms forming a core part of the collections housed in the Geological Museum of the Geological Survey of Namibia (Windhoek) are on Farm Plateau and Farm Aar, located in the south of the country, near the village of Aus. Glaessner noted that the type locale for the holotype of Protechiurus was Farm Plateau and referred to the publications of by R. Richter and H. Pflug (Glaessner, 1979). However, in a more detailed figure from Pflug (1966), all collection points were recorded as being on the Farm Aar, but one must keep in mind that the boundaries of these two farms have changed over time. The most productive localities from which the holotypes of Velancorina martina and Ernietta plateauensis were collected in those early years occur on the eastern part of the Farm Aar, near its border with the Farm Plateau. At the beginning of the 20th century, German soldiers were stationed at this isolated spot, a locale known as Aarhauser, and the soldiers were the first ones who discovered strange Precambrian fossils (Vickers-Rich et al., 2007). Most probably the specimen of Protechiurus was also found in this area.
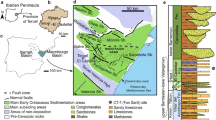
On Farms Aar and Plateau the Late Precambrian sedimentary sequences of the Nama Group are widespread. These sediments include the marine sands, clays, and limestones of the Dabis and Zaris formations, further subdivided into a number of members (Fig. 1). The layers hosting the most abundant macrofossils are confined to the Aar Member, recently distinguished from the Kliphoek Member (Hall et al., 2013). The outcrops of the Kliphoek and Aar members are extensive at Aarhauser. But the layers exposed occur in a variety of slabs, and it is not easy to trace a specific layer along the strike. The local burials of macrofossils are usually confined to specific lenses of sandstone here. These lenses are spindle-shaped, trough-shaped or V-shaped in cross-section and represent paleogutters on the seafloor filled with the sandy sediments. The fossil remains occupy only a part of the lenses and are located in dense accumulations (Pl. 1, figs. 7, 8). Three-dimensional casts in an upright position, with a convex side downward are common among them (Nama-style preservation by G. Narbonne (Narbonne, 2005; Ivantsov, Zakrevskaya, 2018)). The formation of such burials is explained by the transport and burial of the bodies in a flow of suspended sand, with organisms that either had large, widely open cavities into which the sediment penetrated during the transportation, or had life-time accumulations of sand particles within the body (Ivantsov et al., 2016; Vickers-Rich et al., 2013).

Stratigraphic schemes of the Late Precambrian-Early Cambrian deposits of southern Namibia and the northern European Russia (Stankovsky et al., 1990; Vickers-Rich et al., 2016; Grazhdankin, 2016; Linnemann et al., 2019); arrows indicate stratigraphic positions of Protechiurus edmondsi and possible variants along with Russian occurrences of Vendoconularia triradiata.
For many years (since 2004) the study of the macrobiota of the Late Precambrian of Farm Aar and the surrounding areas has been carried out by an international working group led by Professor P. Vickers-Rich (Vickers-Rich et al., 2016; UNESCO IGCP Projects 493, 587 and currently 673). In the course of a complete examination of the Aarhauser tract in 2014, we identified 19 sites with concentrations of macrofossils, some of which contained conical fossils. Sometimes these conical forms were located alongside relatively complete specimens of Pteridinium (Pl. 1, fig. 8); sometimes they were intermixed with fragments of bodies of Pteridinium and Ernietta (Ernietta plateauensis). One monospecific accumulation of small specimens of such conical fossils was also discovered (Pl. 1, fig. 7). We managed to find the slab that had been moulded for Seilacher’s exhibition “Fossil Art” at Aarhauser, which contained the originals of the type series of Vendoglossa. The holotype of this taxon was removed from the slab (Pl. 1, fig. 2) and transferred to the Geological Museum of the Geological Survey of Namibia.
Seventeen samples of conical casts with more or less well-preserved morphological details were also collected for further laboratory study. This study clearly demonstrated that the sand casts preserved as fossils vary significantly in thickness, shape, and degree of sharpness of the ends. Usually, a uniform narrowing towards the closed end, sharpened into a horn, occurs for the most part of the fossil length (Pl. 1, figs. 3, 4, 8). But the closed end is sometimes narrower but swollen and rounded, as in the holotype of Protechiurus edmondsi. The open ends are strongly compressed only in the holotype. Deep, wedge-shaped depressions complicating the open end in more or less fully preserved specimens (Pl. 1, figs. 3, 4) may indicate initial folding of the cone “mouth” (open end), but we did not find sufficient evidence for this. The cross-section of some specimens is distinctly hexagonal (Pl. 1, fig. 5), but in most cases the samples are highly flattened and the symmetry is not so clear. All the differences in morphology can be explained by the post-mortem deformation associated with the flexibility of the original organic matter.
There are longitudinal ridges on the surface of all studied specimens. In case of better preservation of the sculpture, in some specimens there is doubling. As already noted, Glaessner counted 8 longitudinal ridges in the holotype. J. Dzik, who studied the plaster cast of this specimen housed at the South Australian Museum in Australia, found that 2 of these ridges resulted from secondary deformation (Dzik, 2003). Thus, there seem to be only 6 single ridges in Protechiurus. This conclusion is also confirmed by our three-dimensionally preserved specimens. Less pronounced doubled ridges are preserved in rare cases and are only partially visible (Pl. 1, figs. 1, 2). However, the symmetrical arrangement of their fragments in the middle between the single ridges suggests that there were also 6 doubled ridges. The transverse ridges are usually preserved more poorly than the longitudinal ones, and are barely noticeable in the holotype (Pl. 1, figs. 6, 8a). But they can be rather sharp and tuberculate, similar to those observed in the holotype of Vendoglossa tuberculata. The spectrum of the shape and sculpture variability of the conical fossils from Aarhauser overlaps the features of Protechiurus and Vendoglossa, which suggests that these taxa should be synonomized with Protechiurus edmondsi.
As can be judged from the newly discovered specimens, the living Protechiurus was an organism with a hollow body made of elastic material. It was conical, open at the wide end and closed at the pointed end, and hexagonal in cross-section. Its lateral surface hosted longitudinal ridges, both single and double, bounding the rows of the short transverse ridges.
VENDOCONULARIA OF THE WHITE SEA REGION
The sand lenses, representing casts of gutters, occur quite commonly in the Late Vendian deposits of the southeastern White Sea region. Burial of fossil remains with Nama-style preservation was discovered in some of these lenses (Ivantsov, 2015). The Yarnema locality hosting a variety of Precambrian macrofossils, in which the holotype of Vendoconularia was found, entirely consists of such lenses (Ivantsov and Fedonkin, 2002). The locality is confined to a small isolated landslide block surrounded by Carboniferous and Quaternary sediments. Its stratigraphic position is debated. A.F. Stankovsky, while referring to the materials recovered from numerous boreholes drilled in the vicinity of the locality, confidently attributed the block to the lower part of the Arkhangelsk Beds of the Ust-Pinega Formation (Stankovsky and Fedonkin, 2000) (Fig. 1). However, D.V. Grazhdankin, with no justification, placed it within the Verkhovka Formation, which he described himself (Grazhdankin, 2004), and later within the Erga Formation. Both formations lie substantially above the level of the Arkhangelsk Beds in the sequence of the Vendian deposits of the White Sea region (Grazhdankin, 2014).
In the summer of 2015, we discovered a sand lens containing a large, monospecific accumulation of Vendoconularia triradiata near the Yarnema locality. Three joint blocks were preserved in situ. Thirty-seven casts and imprints of this fossil were discovered inside these blocks (Fig. 2). Also, more than 40 casts originating from the same lens were found in the scree rock around the discovery site. These Vendoconularia remains were located in the sandstone together with flat pebbles of mudstone. There were no traces of secondary mineralization of organic matter. Each of the best preserved specimens in the lens occurs in two parts, consisting of a mold of the apical part of the cone in the sandstone and, therefore, three-dimensional, and as well as an imprint of the compressed pre-apertural part, which is two-dimensional and complex-folded. These parts are preserved differently in the rock: the molds oriented for the most part horizontally (Pl. 2, figs. 2, 6, Pl. 3, figs. 4b, 5), and the imprints shifted away from them and preserved upright (vertically) (Pl. 2, fig. 5, Pl. 3, figs. 1–3, 4a). The bend at the place of transition from the horizontal to the vertical does not show any signs of brittle deformation, which means that the initial body of the fossil was soft and flexible. The imprint of the pre-apertural part of the cone is usually located perpendicular to the longitudinal axis of the apical part (Pl. 2, fig. 4); but specimens in which this plane is slightly turned or even located along the axis were also found (Pl. 2, fig. 3). The planes of the vertically oriented imprints of the pre-apertural segment are oriented approximately perpendicular to the longitudinal axis of the lens in all specimens, whereas the orientation of the horizontally oriented sandy molds of the apical part is parallel to the axis of the lens in two approximately opposite directions (Fig. 2b). Based on these observations, we suggest that just prior to final burial, the soft cones of Vendoconularia were transported for a distance in a sandy channel already partially filled. At the same time, the apical parts of the cones, three-dimensional, heavy and filled with the sediment, rolled in the lower levels of the stream, and pre-apertural parts, compressed and light, rose to the upper levels as a flag or sail.

Fragment of a sandy lens containing the remains of Vendoconularia triradiata Ivantsov and Fedonkin, 2002 (I, II, III—separate slabs): (a) top view, (b) a scheme of the location of the identified Vendoconularia specimens (marked with dots and numbers 1–37) and flat argillaceous pebbles (dashed contours on the plan and dashed lines on the side view); (c) slab II, side view; arrows on the scheme and the sectors on the rose diagram indicate the orientation of the apertures of the fossil remains.
All of the elements of the Vendoconularia morphology present in the holotype are also observed on the new material (Pl. 2, figs. 1, 5, Pl. 3, figs. 1–4a). The only exceptions are ribbon-like, longitudinal internal structures, pronounced in the holotype and not on any other specimen. Previously unknown elements, well represented in the new material, represented by wide plates that are attached to the cone along longitudinal unpaired ridges and such directed outward (Pl. 3, figs. 1–5) (Ivantsov, 2017). The presence of plates confirms the above-mentioned assumption by Van Iten et al. (2005) on the separating character of the unpaired ridges in Vendoconularia. The pre-apertural part of the cone is either absent or so strongly crumpled that its structure cannot be established even in broad outlines.

Explanation of Plate 3
Vendoconularia triradiata Ivantsov and Fedonkin, 2002. Imprints of longitudinal plates (marked by arrows) and pre-apertural part of theca (Figs. 2, 4a). All specimens coated with ammonium chloride. Scale bars, 1 cm increments.
Fig. 1. Specimen PIN no. 4564/1047.
Fig. 2. Specimen PIN, no. 4564/1045.
Fig. 3. Specimen PIN no. 4564/1070.
Fig. 4. Specimen PIN no. 4564/1040: a—imprint of the pre-apertural part, b—fragment of the cast of the apical part.
Fig. 5. Specimen PIN no. 4564/1044.
DISCUSSION
A large number of new fossil specimens collected in Namibia and the White Sea region over the last couple of decades now allows us to understand the morphological variations and exclude random features that are not related to the structure of the organism in life. Protechiurus and Vendoconularia now seem to have many similarities, such as the conical shape of an initially soft structure, the closure of the acute end, hexahedral symmetry, the presence of a sculpture consisting of several longitudinal ridges (single and doubled), and an abundance of transverse ridges. The profound similarity between these two genera makes it possible to combine them into one family, Protechiuridae fam. nov. The difference lies in the presence of external longitudinal plates and internal ribbon-like elements in Vendoconularia (Fig. 3). But this difference does not seem significant; along with the particularly fine details of the sculpture unknown in Protechiurus, it may be a consequence of the incomplete preservation of the Namibian fossils. Nevertheless, it creates a formal obstacle for a merging of the Namibian and White Sea genera into one at present until more material is at hand. The presence of wide longitudinal plates directed outwards in Vendoconularia strengthens the hypothesis of the external position of the conical structure relative to the body. Most likely it was an external supporting and protective structure, similar to the theca of cnidarians (Ivantsov and Fedonkin, 2002).

Schematic reconstruction of the theca of protechiurids: (a) Protechiurus edmondsi, (b) Vendoconularia triradiata (b'—cross-section).
The taxonomic position of protechiurids is controversial. The similarity of the elements of the surface sculpture of Protechiurus with those of some modern echiurids (transverse rows of papillae, longitudinal furrows and ridges) is complicated by the absence of a satisfactory hypothesis explaining the mechanism of formation of a sandy cast of a soft structure closed on all sides, such as in the body of the echiurids. Penetration of the sediment into the internal cavity, which is demonstrated by all known specimens of the protechiurids, from both the Namibian and White Sea localities, indicates that the walls of the buried structure were for some time sufficiently elastic to maintain the cavity volume with an open aperture. Then, as can be seen in the White Sea material, the pre-apertural part shrank, and further penetration of the sediment became impossible. Both an exuvia of ecdysozoans, lost as a result of molting, and an organic “shell” left by an animal from some other group, could have behaved in a similar way during burial. Closure of one end and the presence of radial longitudinal plates supports the second hypothesis. Based on the main morphological features, the details of the surface sculpture and the sizes (the first centimeters), the protechiurids are strikingly similar to conulariids (Cnidaria, Scyphozoa). Thecae of the conulariids are conical, radially-symmetrical, and faceted. A sculpture of their surface is represented by two rows of transverse ridges, often scalloped along one edge, and sometimes changed into chains of tubercles. The symmetry of the sliding reflection is often observed in the mutual arrangement of the ridges. The difference lies in the absence of any trace of primary mineralization and the 6-fold radial symmetry of the protechiurid remains, whereas conulariid thecae consist of calcium phosphate with normally four-fold symmetry. However, a “soft-bodied character” is common for most Vendian Metazoa, and can be explained in several ways—from an ability for biomineralization, not yet obtained, or an impossibility to preserve in this manner in the Precambrian biosphere, to a non-preservation of a poor mineralization in terrigenous rocks, in which these remains usually occur. It is obvious that the “soft-bodied character” of fossil remains cannot be used as strong proof for the lack of relationship between the Vendian and Paleozoic organisms, in particular, the protechiurids and cnidarians. The radial symmetry represents the greatest difficulty in establishing of the affinity of the protechiurids and conulariids. But 6‑fold symmetry of Hexaconularia He et Yang, 1986, the Early Cambrian conulariid, gives some support to the alliance of our Precambrian fossils (Van Iten et al., 2010).
The new material from the Arkhangel’sk region reflects the development of wide longitudinal plates separating the sides of the thecae from each other and facing outward in Vendoconularia. Such elements are unknown in the conulariids. But this sort of structure is found in some anabaritids, a group of small fossils of an unspecified systematic position, common in the carbonate sediments of Vendian-Cambrian boundary sediments worldwide (Kouchinsky et al., 2009). In Anabarites tricarinatus Missarzhevsky, 1969 from the Lower Cambrian of the Siberian platform, the plates reach maximum development and are comparable in width to those of Vendoconularia (Fig. 4) (Abaimova, 1978). The 3-fold radial symmetry, sometimes transforming into 6-fold symmetry, is characteristic for usually narrow-conical remains of the anabaritids, and their outer surface can have a corrugated sculpture. But the anabaritids are much smaller in size than the protechiurids, and their thecae had a primary carbonate, probably aragonitic, mineralization (Kouchinsky, Bengtson, 2002). In anabaritids, the sculpture of the thecal surface is much simpler than that of conulariids and protechiurids (Kouchinsky et al., 2009). However, a number of anabaritids are known only from internal molds composed of calcium phosphate. The initial shell material is not preserved or it was lost as a result of the acid treatment of samples, the traditional method of extraction of SSF. Therefore, finding anabaritids with a diverse and more complex surface sculpture is quite possible in the future.

Various forms of the theca of the anabaritid, Anabarites tricarinatus Missarzhevskii, 1969: (a) straight, (b) spirally twisted (according to Abaimova, 1978).
Thus, the protechiurids show features of both conulariids and anabaritids and may represent a link between these unique, possibly related groups of ancient organisms (Abaimova, 1978; Valkov, 1982).
CONCLUSIONS
Our research supports the hypothesis proposed by G.P. Abaimova and A.K. Valkov concerning the affinity of anabaritids and conulariids. Both groups have in the past been classified as scyphozoans due to the fact that all three of these groups: 1) possess a radially symmetrical thecae, 2) have 3 / 6-fold taxa; and 3) are present in the Late Precambrian. The difference in the theca mineralization can be explained by the long-established divergence of these groups: it may have occurred at a time when the mechanism of biomineralization in the cnidarians had not been developed, and the order of radial symmetry has not been firmly established. The protechiurids may belong to the basal group of ancient scyphozoans, ancestral to the conulariids on the one hand, and to the anabaritids on the other.
-
SYSTEMATIC DESCRIPTION
-
Family Protechiuridae fam. nov.
Distinctive features. Originally non-mineralized conical theca, round-hexagonal in cross-section, open at the wide end (aperture) and closed at the pointed (apical) end. The surface of the theca hosts 12 alternating longitudinal ridges, 6 single and 6 double, bounding rows of short transverse ridges scalloped along the upper edge.
Composition. Two genera: Protechiurus Glaessner, 1979 (=Vendoglossa tuberculata Seilacher, 2007) and Vendoconularia Ivantsov et Fedonkin, 2002.
-
Genus Vendoconularia Ivantsov et Fedonkin, 2002 emend. Ivantsov, nov.
Vendoconularia: Ivantsov, Fedonkin, 2002, p. 1227.
Type species—Vendoconularia triradiata Ivantsov et Fedonkin, 2002.
Initial diagnosis (Ivantsov, Fedonkin, 2002). Conical test with 6 faces and specific pattern of the arrangement of ridges, exhibiting two series of main and secondary ridges both having short spines directed towards the aperture.
Changed diagnosis. Theca with 6 wide, outwardly directed longitudinal plates.
Comparison. It differs from Protechiurus by the presence of longitudinal plates.
Composition. The genus is monospecific.
Distribution. Late Vendian of the Arkhangelsk Region.
-
Vendoconularia triradiata Ivantsov et Fedonkin, 2002 emend. Ivantsov, nov.
-
Pl. 2, fig. 1a–1d
Vendoconularia triradiata: Ivantsov, Fedonkin, 2002, p. 1227; Fedonkin et al., 2007, p. 144; Ivantsov, Leonov, 2009, p. 63; Van Iten et al., 2005, p. 619; Ivantsov, 2017, p. 129.
Holotype—PIN, No. 4564/1025, shown in: Ivantsov, Fedonkin, 2002, text Fig. 2, pl. 1, Figs. 1–4; Arkhangelsk region, right bank of the Onega river, 1 km below the mouth of the Somba river, Yarnema locality; Upper Vendian, Ust-Pinega (?) Formation, local accumulation Y2 (IV).
Initial description (Ivantsov, Fedonkin, 2002). Conical, elongated exoskeleton with a thin, flexible, non-mineralised wall comprising 6 identical faces. External surface of a face bears two rows of fine, transverse ridges (rods?) situated in an alternating position, and lateral longitudinal ridges extending along corner groove. There are two kinds of regularly alternating rods in every row of the face: main rods connect the midline and lateral ridges of the face; secondary rods do not reach the lateral ridge and their distal ends are turned towards the aperture joining the next main rod. Rods of the opposite row on the same face are situated in an alternating position (symmetry of gliding reflection). Apertural side of the rods bears miniature spines (adapertural spines). Internal anatomy is represented by three linear structures extending along every other corner groove closer to the narrow apical part of the test. Apertural lobes are not preserved or absent.
New description. Elongated conical, originally unmineralized theca, round-hexagonal in cross-section, closed at the pointed apical end, bearing 6 thin, outwardly directed longitudinal plates. On the surface of the theca there are 12 alternating longitudinal ridges, 6 unpaired and 6 double, bounded on the sides by rows of short transverse ridges scalloped along the upper edge. In each row, there are two types of regularly alternating transverse ridges: the main ones are joined by both double and single longitudinal ridges, while the secondary transverse ridges do not reach the double ridge; their distal ends are turned towards the aperture joining the next main transverse ridge. The ridges of the opposite rows are arranged in an alternating order. Unpaired longitudinal ridges are the bases for the longitudinal plates. An internal anatomy is represented by three linear structures extending along each second double longitudinal ridge in proximity to the apical part of the theca.
Distribution. Arkhangelsk region, southeastern White Sea area, Yarnema locality; Late Vendian, Ust-Pinega (?) Formation.
Material. About 80 specimens from a single locality.
REFERENCES
Abaimova, G.P., Anabaritides—the oldest fossils with a carbonate skeleton, Novye materialy po stratigrafii i paleontologii Sibiri (New Data on the Stratigraphy and Paleontology of Siberia), Tr. Sib. Nauchno-Issled. Inst. Geol. Geofiz. Miner. Syr. (SNIIGGIMS), vol. 260, Novosibirsk: Nauka, 1978, pp. 77–83.
Babcock, L.E. and Feldmann, R.M., Devonian and Mississippian Conulariids of North America. Part A. General description and Conularia, Ann. Carnegie Mus., vol. 55, Pittsburgh, Pennsylvania, 1986, pp. 349–410.
Dzik, J., Anatomical information content in the Ediacaran fossils and their possible zoological affinities, Integr. Comp. Biol., 2003, vol. 43, no. 1, pp. 114–126.
Glaessner, M.F., An echiurid worm from the Late Precambrian, Lethaia, 1979, vol. 12, no. 2, pp. 121–124.
Grazhdankin, D., Patterns of distribution in the Ediacaran biotas: Facies versus biogeography and evolution, Paleobiology, 2004, vol. 30, no. 2, pp. 203–221.
Grazhdankin, D.V., Patterns of evolution of the Ediacaran soft-bodied biota, J. Paleontol., 2014, vol. 88, no. 2, pp. 269–283.
Grazhdankin, D., Forbidden fruits in the Garden of Ediacara, Paläontol. Z., 2016, vol. 90, no. 4, pp. 649–657.
Hall, M., Kaufman, A.J., Vickers-Rich, H., Ivantsov, A., Trusler, P., Linnemann, U., Hofmann, M., Elliott, D., Cui, H., Fedonkin, M., Hoffmann, K.-H., Wilson, S.A., Schneider, G., and Smith, J., Stratigraphy, palaeontology and geochemistry of the Late Neoproterozoic Aar Member, southwest Namibia: Reflecting environmental controls on Ediacara fossil preservation during the terminal Proterozoic in African Gondwana, Precambrian Res., 2013, vol. 238, pp. 214–232.
van Iten, H., Leme, J.M., Rodrigues, S.C., and Simões, M.G., Reinterpretation of a conulariid-like fossil from the Vendian of Russia, Palaeontology, 2005, vol. 48, no. 3, pp. 619–622.
van Iten, H., Zhu Mao-yan, and Li Guo-xiang, Redescription of Hexaconularia He and Yang, 1986 (Lower Cambrian, South China): implications for the affinities of conulariid-like small shelly fossils, Palaeontology, 2010, vol. 53, no. 1, pp. 191–199.
Ivantsov, A.Yu., Yuzhno-afrikanskii element v soobshchestve vendskikh organizmov Arkhangel’skoi oblasti (The South African Element in the Community of Vendian Organisms in the Arkhangelsk Region), Tr. Arkhangel. Tsentra Ross. Geogr. Ob-va, no. 3, Arkhangelsk: Arkhangel. Tsentr Ross. Geogr. Ob-va, 2015, pp. 194–201.
Ivantsov, A.Yu., The most probable Eumetazoa among late Precambrian macrofossils, Invertebr. Zool., 2017, vol. 14, no. 2, pp. 127–133.
Ivantsov, A.Yu. and Fedonkin, M.A., Conulariid-like fossil from the Vendian of Russia: a metazoan clade across the Proterozoic/Palaeozoic boundary, Palaeontology, 2002, vol. 45, no. 6, pp. 1219–1229.
Ivantsov, A.Yu. and Zakrevskaya, M.A., Fenomen isklyuchitel’noi sokhrannosti pozdnedokembriiskikh makrofossilii (The Phenomenon of Exceptional Preservation of Late Precambrian Macrofossils), Tr. Paleontol. Ob-va, vol. 1, Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2018, pp. 46–53.
Ivantsov, A.Yu., Narbonne, G.M., Trusler, P.W., Greentree, C., and Vickers-Rich, P., Elucidating Ernietta: New insights from exceptional specimens in the Ediacaran of Namibia, Lethaia, 2016, vol. 49, no. 4, pp. 540–554.
Kouchinsky, A. and Bengtson, S., The tube wall of Cambrian anabaritids, Acta Palaeontol. Pol., 2002, vol. 47, no. 3, pp. 431–444.
Kouchinsky, A., Bengtson, S., Feng Wei-min, Kutygin, R., and Valkov, A.K., The Lower Cambrian fossil anabaritids: affinities, occurrences and systematics, J. Syst. Pal., 2009, vol. 7, no. 3, pp. 241–298.
Linnemann, U., Ovtcharova, M., Schaltegger, U., Gärtner, A., Hautmann, M., Geyer, G., Vickers-Rich, P., Rich, T., Plessen, B., Hofmann, M., Zieger, J., Krause, R., Kriesfeld, L., and Smith, J., New high-resolution age data from the Ediacaran-Cambrian boundary indicate rapid, ecologically driven onset of the Cambrian explosion, Terra Nova, 2019, vol. 31, pp. 49–58.
Narbonne, G.M., The Ediacara biota: Neoproterozoic origin of animals and their ecosystems, Annu. Rev. Earth Pl. Sci., 2005, vol. 33, no. 1, pp. 421–442.
Pflug, H.D., Neue Fossilreste aus den Nama-Schichten in Südwest-Afrika, Paläontol. Z., 1966, vol. 40, nos. 1–2, pp. 14–25.
Seilacher, A., The nature of vendobionts, The Rise and Fall of the Ediacaran Biota, Vickers-Rich, P. and Komarower, P., Eds., Geol. Soc. Spec. Publ., vol. 286, London: Geol. Soc. Lond., 2007.
Stankovsky, A.F. and Fedonkin, M.A., The localities of the Vendian soft-bodied fauna in the southeastern White Sea region, Ocherki po geologii i poleznym iskopaemym Arkhangel’skoi oblasti (Essays on Geology and Mineral Resources of the Arkhangelsk Region), Galimzyanov, R.M., Ed., Arkhangelsk: Pomorsk. Gos. Univ. im. M.V. Lomonosova, 2000, pp. 142–153.
Stankovsky, A.F., Verichev, E.M., and Dobeiko, M.P., Vendian of the Southeastern White Sea Area, Vendskaya sistema. Istoriko-geologicheskoe i paleontologicheskoe obosnovanie (The Vendian System: Historical Geology and Paleontological Substantiation), vol. 2: Stratigrafiya i geologicheskie protsessy (Stratigraphy and Geological Processes), Sokolov, B.S. and Fedonkin, M.A., Eds., Moscow: Nauka, 1985, pp. 67–76.
Valkov, A.K., Biostratigrafiya nizhnego kembriya vostoka Sibirskoi platformy (Uchuro-maiskii raion) (Biostratigraphy of the Lower Cambrian of the East of the Siberian Platform, Uchur–Maya Region), Moscow: Nauka, 1982.
Vickers-Rich, P., Ivantsov, A.Yu., Trusler, P.W., Narbonne, G.M., Hall, M., Wilson, S.A., Greentree, C., Fedonkin, M.A., Elliot, D.A., Hoffmann, K.H., and Schneider, G.I., Reconstructing Rangea: new discoveries from the Ediacaran of southern Namibia, J. Paleontol., 2013, vol. 87, no. 1, pp. 1–15.
Vickers-Rich, P., Narbonne, G., Laflamme, M., Darroch, S., Kaufman, A.J., Kriesfeld, L., Hall, M., Trusler, P., Smith, J., Elliott, D., Sharp, A., Ivantsov, A., Fedonkin, M., Schneider, G., Hoffmann, C., et al., The Nama Group of Southern Namibia: The End Game of the First Large, Complex Organisms on Earth, the Ediacarans [IGCP493/587], 35th International Geological Congress, 2016 Pre-conference Field Guide, 21–25 August, Namibia, Dandenong North, Victoria: New Artworx P/L, 2016.
ACKNOWLEDGMENTS
The studied samples are stored in the Geological Museum of the Geological Survey of Namibia (Windhoek) and in the Paleontological Institute of Russian Academy of Sciences (PIN RAS). Photographs were made by S.V. Bagirov (PIN RAS). The field work and the preparation of the fossil material were carried out with the support of the Russian Foundation for Basic Research, Grant no. 17-05-02212-a.
Funding
The laboratory and theoretical studies are supported by the grant for the fundamental research on the topic of the state assignment of PIN RAS, “The emergence of life, the formation of the biosphere, and the development of ancient biotas”. The work in Namibia was carried out as part of the International Geosciences Programs IGCP 493, 587and 673 and in part supported by several grants from the National Geographic Society and with the assistance of the Geological Survey of Namibia, especially the then Director Dr Gabi Schneider and with the field support of Charlie Hoffmann and local landowners Barbara and Brunо Boehm-Ernie, Lothar Gessart and Bernd and Lela Roemer.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by M. Zakrevskaya
Rights and permissions
About this article

Cite this article
Ivantsov, A.Y., Vickers-Rich, P., Zakrevskaya, M.A. et al. Conical Thecae of Precambrian Macroorganisms. Paleontol. J. 53, 1134–1146 (2019). https://doi.org/10.1134/S0031030119110054
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030119110054