
Abstract—
While lanthanum is known to regulate the metabolism of microorganisms using single-carbon compounds, there is no information about its effect on soil communities. This is the first report on response of methanotrophic communities to the introduction of lanthanum, determined using high-throughput sequencing of the 16S rRNA genes in experiments with soil microcosms. It was found that after one and two months after the introduction of lanthanum salts the proportion of Methylobacter in the total pool of sequences increased (up to 9 and 15%, respectively). At the same time, the content of methylotrophic Methylotenera increased up to 10 and 19%, respectively. Thus, lanthanum was found to stimulate the formation of Methylobacter −Methylotenera associations under elevated methane content in the soil, which may affect the contribution of agrosoils to the regulation of methane content in the atmosphere.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Methane is an important greenhouse gas, and although its content in the atmosphere is less than 0.02%, its contribution to current global warming is estimated at 15% (Saunois et al., 2020). The only known biological way of absorbing methane from the Earth’s atmosphere is its oxidation by microbial communities in aerobic soils; therefore, any changes in the intensity of this process can have global consequences. According to modern calculations, the intensity of methane absorption by soil microorganisms is 22.4 Tg yr–1, of which half is in the soils of the temperate zone (Dutaur and Verchot, 2007).
The soils of agrocenoses are under the constant influence of various compounds supplied with atmospheric precipitation, fertilizers, emissions from enterprises and vehicles (Smith et al., 2016). Soil microbial communities are responsible for the implementation of the most important ecosystem processes, which causes an ever-increasing interest in studying their resilience and recovery after stressful impacts (Griffiths and Philippot, 2013). Data on changes in the composition of methanotrophic communities during the application of fertilizers are very limited, and nitrogen compounds are most often considered. Very little is known about the effect of other elements, such as lanthanides, on soil microbial communities.
Lanthanides are included in the group of rare earth elements (REE), which are widely used in modern technologies (Vodyanitskii and Rogova, 2016). The main part of the lanthanides that enter the soil during the decomposition of lithogenic minerals is contained in organometallic complexes and is inaccessible to plants and microorganisms. (Kotel’nikova et al., 2021). At the same time, lanthanides that enter the soil as a result of anthropogenic activity (mineral and organic fertilizers, microfertilizers) are in a biologically available form and can disrupt the REE cycle in the environment (Ramos et al., 2016). In the last decade, much attention has been paid to lanthanides, especially light ones, due to their stimulating effect on plant growth (Kotel’nikova et al., 2021), while information on the effect on soil microorganisms is very limited. As an example, systematic studies of the effect of lanthanides on nitrifying and ammonifying soil microorganisms in Buryatia have been carried out for a number of years (Chimitdorzhieva and Abasheeva, 2014).
The discovery of lanthanide-dependent methanol dehydrogenase (XoxF-MDH) (Chistoserdova, 2016) demonstrated the important role of rare earth elements in the metabolism of methylotrophs. Light lanthanides, especially lanthanum (La), can effectively replace calcium in methanol dehydrogenase. As a result, the presence of lanthanides, even in nanomolar amounts, regulates the expression of genes for the synthesis of alternative methanol dehydrogenase and affects the formation of extracellular methanol by methanotrophs, which can lead to changes in the composition of satellite microorganisms (Krause et al., 2017). This opens up prospects for developing new approaches to regulating the composition and activity of soil methanotrophs and their associated microbiota. At the same time, the authors are not aware of works on the assessment of the effect of lanthanum or other lanthanides on methanotrophic soil communities.
The aim of this study was to evaluate the effect of lanthanum salts on the composition of methanotrophic communities in experiments with soil microcosms.
In November 2020, a series of experiments was laid with sod-podzolic medium loamy soil of the surface horizon A (0–20 cm) of a fallow forb-grass meadow. The object is located near the Poshekhonskaya poultry farm, Yaroslavl oblast (58°30′36′′ N and 39°08′32′′ E), and has been used in crop rotation with the introduction of high doses of organic fertilizer for more than 20 years. The soil is moderately acidic (pH 5.4), with high content of organic matter (Corg 21.7 g kg–1 according to Tyurin), high content of nitrates (261 mg kg–1), and low content of ammonium nitrogen (<5 mg kg–1).
The original soil (option K) was used to create microcosms, each of which was 10 g of soil of natural moisture placed in a 100-mL glass vial. The vials were placed in a desiccator with a gas mixture consisting of 10% of methane and 90% of air for incubation for two months at 25°C. Once a week, the gas phase was replaced and the soil moisture was controlled gravimetrically. The following options were investigated: M, no additives; ML1, addition of lanthanum chloride solution (5 μg La3+), incubation for one month; ML2, introduction of a lanthanum chloride solution (5 μg La3+), incubation for two months. Two separate samples per variant were used for DNA extraction and molecular analysis, a total of 8 samples.
Total DNA was isolated from 0.25 g of soil using the Power Soil DNA Isolation Kit (Qiagen, Carlsbad, CA, United States). The composition of prokaryotic soil communities was assessed by high-throughput sequencing of the variable V3–V4 region of 16S rRNA gene on a MiSeq sequencer (Illumina, United States), and the initial software processing of the obtained sequences was carried out at Biospark LLC. Two libraries were created for each DNA sample. Library analysis was performed by end-paired reads generating at least 10 000 paired reads per sample using the following reagents: MiSeq Reagent Kit v2 nano and MiSeq v2 Reagent Kit. The sequencing data obtained were processed in a software written using the QIIME 1.9.1 algorithm, including the combination of forward and reverse reads, deletion of technical sequences, filtering of sequences with low reading reliability of individual nucleotides (quality less than Q30), filtering of chimeric sequences, alignment of reads to the reference 16S rRNA sequence. When calculating the diversity indexes, the data were normalized according to the sample with a minimum number of reads. The distribution of sequences by OTUs using the Silva database v. 132 and the calculation of diversity indexes were performed using the QIIME 2 software (Caporaso et al., 2010). The OTU classification algorithm with an open reference was used; the classification threshold was 97%.
The analysis performed showed that the share of methanotrophic 16S rRNA gene sequences in the original sod-podzolic soil was less than 0.1% of all identified sequences. Bacteria of the genera Methylosinus (0.05 ± 0.01%), Methylocystis (0.01 ± 0.006%), and Methylomonas (0.004 ± 0.002%) were detected.
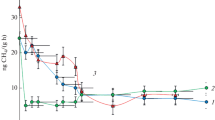
The incubation of microcosms with methane, as well as methane and lanthanum salts, reduced the diversity of the microbial community (Table 1), but increased the share of methanotrophs and changed the structure of the methanotrophic community. After 30 days of incubation with methane, the amount of Methylosinus increased to 0.38 ± 0.08%. The community also contained Methylobacter (2 OTUs, 0.43 ± 0.11%) and single sequences of Methylocella (0.03 ± 0.01%) (Fig. 1), whose number in the initial soil was below the detection level. Thus, the number of methanotrophs after 1 month of incubation increased by an order of magnitude and amounted to about 1% of the total number of sequences.

Indicators of the relative abundance of the dominant methanotrophic bacteria Methylobacter (white columns) and methylotrophic bacteria Methylotenera (black columns) in the soil samples of the microcosm experiment. Designations of experiment options: 1, M; 2, ML1; 3, ML2.
The introduction of lanthanum fundamentally changed the number and composition of methanotrophs. One month later, the share of 16S rRNA gene sequences in methanotrophs reached 9.2%, of which Methylobacter constituted 8.97 ± 0.9% (3 OTUs) and Methylosinus 0.10 ± 0.08% (Fig. 1). The effect of lanthanum was prolonged, and after two months the share of Methylobacter gene sequences increased to 15.28 ± 3.2% (6 OTUs) and that of Methylosinus to 0.19 ± 0.1% (Fig. 1). All Methylobacter OTUs belonged to M. tundripaludum (98.12–99.06% identity of the 16S rRNA gene with the type strain), while Methylosinus OTUs belonged to M. sporium (99.24% identity) (Table 2).
The methylotrophs of the genus Methylotenera assigned to the class Methylophilaceae (Fig. 1) formed consortiums with methanotrophic bacteria in shallow methane seeps (Danilova et al., 2021) and lake sediments (van Grinsven et al., 2021). Two months later, the share of Methylotenera 16S rRNA gene sequences exceeded 19 ± 1.30% (Fig. 1). All of them belonged to M. versatilis (Table 2) showing 98.12–98.46% identity of 16S rRNA with the isolate from the bottom sediments of Lake Washington, United States (Lapidus et al., 2011). The methylotrophic bacteria Hyphomicrobium facile (2 OTUs, 0.4 ± 0.2%) were also found in the variants without the introduction of lanthanum.
Thus, it has been established for the first time that the introduction of lanthanum in the soil of the temperate climatic zone against the background of an increase in the methane content, which is typical of agricultural soils with the application of organic and mineral fertilizers, leads to the absolute dominance of methanotrophs of the genus Methylobacter and obligate methylotrophs of the genus Methylotenera. The coordinated response of Methylobacter and Methylotenera to the addition of lanthanum salt suggests their joint activity in the regulation of methane exchange between the soil and the atmosphere, which requires further study.
REFERENCES
Caporaso, J.G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F.D., Costello, E.K., Fierer, N., Peña, A.G., Goodrich, J.K., Gordon, J.I., Huttley, G.A., Kelley, S.T., Knights, D., Koenig, J.E., Ley, R.E., et al., QIIME allows analysis of high throughput community sequencing data, Nature Meth., 2010, vol. 7, pp. 335–336.
Chimitdorzhieva, I.B. and Abasheeva, N.E., Effect lantana na mikrobiologicheckuyu aktivnost’ dinamiku azotnogo fonda pochv (Effect of Lanthanum on Soil Microbiological Activity and Dynamics of the Nitrogen Fund), Ulan-Ude, BG-SKhA, 2014. Chistoserdova, L., Lanthanides: new life metals?, World J. Microbiol. Biotechnol., 2016, vol. 32, article 138. https://doi.org/10.1007/s11274-016-2088-2
Danilova, O.V., Ivanova, A.A., Terent’eva, I.E., Glagolev, M.V., and Sabrekov, A.F., Microbial community composition of floodplains shallow-water seeps in the Bolshaya Rechka floodplain, Western Siberia, Microbiology (Moscow), 2021, vol. 90, pp. 632‒642.
Dutaur, L. and Verchot, L.V., A global inventory of the soil CH4 sink, Glob. Biogeochem. Cycles, 2007, vol. 21, pp. 1–9.
Griffiths, B.S. and Philippot, L., Insights into the resistance and resilience of the soil microbial community, FEMS Microbiol. Rev., 2013, vol. 37, pp. 112–129.
Kotelnikova, A.D., Rogova, O.B., and Stolbova, V.V., Lanthanides in the soil: routes of entry, content, effect on plants, and genotoxicity (a review), Euras. Soil Sci., 2021, vol. 54, pp. 117‒134.
Krause, S.M.B., Johnson, T., Karunaratne, Y.S., Fu, Y., Beck, D.A.C., Chistoserdova, L., and Lidstrom, M.E., Lanthanide-dependent cross-feeding of methane-derived carbon is linked by microbial community interactions, Proc. Natl. Acad. Sci. U. S. A., 2017, vol. 114, pp. 358–363.
Lapidus, A., Clum, A., LaButti, K., Kaluzhnaya, M.G., Lim, S., Beck D.A.C., Glavina del Rio T., Nolan, M., Mavromatis, K., Huntemann, M., Lucas, S., Lidstrom, M.E., Ivanova, N., and Chistoserdova, L., Genomes of three methylotrophs from a single niche reveal the genetic and metabolic divergence of the Methylophilaceae, J. Bacteriol., 2011, vol. 193, pp. 3757‒3764.
Ramos, S.J., Dinali, G.S., Oliveira, C., Martins, G.C., Moreira, C.G., Siqueira, J.O., and Guilherme, L.R., Rare earth elements in the soil environment, Curr. Pollut. Rep., 2016, vol. 2, pp. 28–50.
Saunois, M., Bousquet, P., Poulter, B., Peregon, A., Ciais, P., Canadell, J.G., Dlugokencky, E.J., Etiope, G., Bastviken, D., Houweling, S., Janssens-Maenhout, G., Tubiello, F.N., Castaldi, S., Jackson, R.B., Alexe, M., et al., The global methane budget 2000–2012, Earth Syst. Sci. Data, 2016, vol. 8, pp. 697–751.
Smith, P., House, J.I., Bustamante, M., Sobocká, J., Harper, R., Pan, G., West, P.C., Clark, J.M., Adhya, T., Rumpel, C., Paustian, K., Kuikman, P.M., Cotrufo, F., Elliott, J.A., McDowell, R., et al., Global change pressures on soils from land use and management, Global Change B-iol., 2016, vol. 22, pp. 1008‒1028.
van Grinsven, S., Damsté, J.S.S., Harrison, J., Polerecky, L., and Villanueva, L., Nitrate promotes the transfer of methane-derived carbon from the methanotroph Methylobacter sp. to the methylotroph Methylotenera sp. in eutrophic lake water, Limnol. Oceanogr., 2021, vol. 66, pp. 878–891.
Vodyanitskii, Yu.N. and Rogova, O.B., The biogeochemistry of lantanides in soils, Dokuchaev Soil Bull., 2016, vol. 84, pp. 101‒118.
Funding
This study was supported by the Russian Science Foundation, project no. 22-24-00418.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement of the welfare of animals. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
Translated by E. Babchenko
Rights and permissions
About this article

Cite this article
Kravchenko, I.K., Sizov, L.R., Tikhonova, E.N. et al. The Effect of Lanthanum on the Composition of Methanotrophic Community of Sod-Podzolic Soil. Microbiology 91, 599–603 (2022). https://doi.org/10.1134/S002626172260135X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S002626172260135X