
Abstract
To reveal the distribution patterns of pollutants in the elementary landscape-geochemical systems (ELGS) (top–slope–closing depression), the 137Cs distribution was studied in detail in the soil–moss cover of the test site, which characterizes undisturbed forest landscapes subjected to the fallout of technogenic radionuclides during the Chernobyl accident. Over the thirty years after the primary contamination, no unidirectional 137Cs migration from top to the closing depression was found in the ELGS. In contrast, all studied ELGS show the well expressed 137Cs cyclical variation, which is traced both in the soil and in the vegetation (moss) cover. The pattern of 137Cs variability in the moss cover is generally similar to the surface soil contamination, but in some cases demonstrates specific features. At present, the top 8-cm soil layer contains from 70 to 96% of total 137Cs content of soil cover, while 0–20-cm layer hosts 89–99% 137Cs. Such vertical distribution makes it possible to apply field spectrometry technique for study of the spatial structure of 137Cs contamination field. The 137Cs content in the photosynthesizing part of the moss best correlates with its reserve in the 2–4 cm soil layer (r0.01 = 0.747, n = 15). The revealed structural heterogeneity results from secondary redistribution of the radionuclide in ELGS and, in our opinion, reflects the general regularities of element migration in such systems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
The elementary landscape-geochemical system (top–slope–closing depression) is a primary chain in the spatial structure of any landscape. Therefore, the detailed study of distribution of elements and/or their compounds is of obvious interest. Due to significant practical importance, the most detailed studies of element migration in relief were aimed at determining the activity of soil erosion, frequently using 137Cs as a radioactive marker for obtaining experimental data (Walling D.E., He, 1999; Olson et al., 2008). In Russia, the Bryansk region experienced the strongest radioactive impact during the 1986 Chernobyl accident. Its western part with the 137Cs contamination density more than 40 Ci/km2 (1480 kBq/m2) was ascribed to the abandoned zone and was excluded from exploitation (Izrael’ et al., 1998; De Cort et al., 1998). This made it possible to apply field gamma-spectrometry for studying the structure of forming secondary 137Cs contamination fields, in particular, within ELGS. Among components of the ground cover, mosses were selected for monitoring the vegetation contamination due to their ubiquitous abundance and long lifetime (Nifontova, 2003; Aleksiayenak et al., 2013; Cevik and Celik, 2009; Florek et al., 2001; Steinnes and Njåstad, 1993; Gulan et al., 2020а; Malikova et al., 2019; Sawidis et al., 1997; Sawidis, 1984).
Our work is aimed at studying the spatial 137Cs distribution as a tracer, which allows identifying the regularities of element migration in soil and plant cover of ELGS 30 year after the primary contamination and over two-fold change of moss cover and needles of pine representing the predominant species of local woody vegetation.
MATERIALS AND METHODS
In 1990, the Russian Scientific-Practical and Expert-Analytical Centre under the State Chernobyl Committee of the Russian Federation set up 19 test sites in the abandoned zone of the Bryansk region to study and control the migration of the Chernobyl radionuclides under different landscape conditions within distinguished radioecological test site (Linnik et al., 1993). Starting in 2005, one of these test sites conditionally termed “Vyshkov-2” was annually used by collaborators of the Laboratory of Environmental Biogeochemistry of GEOKHI RAS to study the secondary migration of 137Cs as a technogenic tracer of pollutant redistribution in the soil–plant cover of the elementary landscape–geochemical systems (ELGS) (Korobova, 2006).
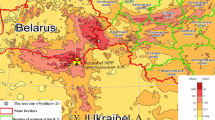
This test site is located 15 km SE of the town of Novozybkov on the high terrace of the Iput river, which is occupied by the pine forest with Hylocomium splendens, Dicranum, etc. (Bryidae subclass) grown on the sod–podzolic illuvial–ferruginous sandy loamy soils formed on the rewashed sandy deposits. Relative elevation range in the site is about 4.5 m (Fig. 1a). The soil cover has well expressed litter layer and humic horizon, with no traces of replowing.

Location of the Vyshkov-2 test site (a) and profiles D-21 and D-25 with points of 137Cs activity measurement (b).
The spatial distribution of 137Cs was studied along two parallel 60-m long landscape–geochemical profiles cutting across ravine-like depression in the northeastern part of the test site (Fig. 1b), which is ubiquitously covered by moss cover dominated by Pleurozium schreberi, with the less contribution of two other species (Dicranum polysetum and Polytrichum commune) (<10%).
Each profile characterizes the specifics of radionuclide distribution in two adjacent ELGS of the northern and southern expositions. The analysis of four conjugated ELGS pairs allowed us to make justified conclusions on the migration tendencies, which are typical of the systems of the studied type.
The 137Cs distribution pattern in the soil cover of ELGS was studied by the measurement of 137Cs activity using a Violinist-III modified portable gamma spectrometer equipped with a collimated scintillation detector allowing the determination of integral activity of radionuclide in a hemisphere 20 cm across (Romanov et al., 2011).
An absolute error in field determinations of the activity was no more than 5–10 imp/s. Before measuring the 137Cs activity in ELGS with a step of 1 m, the relative height of each point was determined using the Boif DJD-20 theodolite to ensure accuracy up to 1 cm in changing the relative height. The measurement points in different years were numbered in opposite directions. Therefore, point 60 of profile D-21 corresponded to point 0 of profile D-25 (Fig. 1b).
To study the 137Cs distribution, plant samples were collected using a metallic ring 14 cm in diameter in each point of field measurements. In laboratory conditions, moss samples were separated into photosynthesizing (green) and nonphotosynthesizing (stem + rhizoid, hereafter rhizoid) parts with preliminary extraction of litter fall consisting mainly of pine needles.
To verify field measurements of 137Cs activity and to estimate its vertical migration, soil cores were collected in separate points of ELGS. Sampling was carried out using marked soil corer down to a depth of 40 cm, with subsequent subdivision of the core into 2‑cm layers down to a depth of 20 cm and into 5-cm layers below 20 cm. All samples were placed into plastic bags and weighed for subsequent determination of their natural moisture. The samples were preserved under dark cold conditions up to the measurement of 137Cs content in the laboratory using a stationary Canberra gamma spectrometer (USA). Error of 137Cs determination was no more than 1–5%, although sometimes reached 30–40% in the lower soil horizons with low RN content.
Data were treated using MS Excel and STATISTICA softwares. The period of variation of parameters in soil and moss cover was determined using Fourier Transform spectral analysis. The activity of 137Cs accumulation in the moss cover was estimated by relation of its accumulation in biomass to a weight unit of soil (accumulation coefficient, Ka) and to a soil contamination density unit (transfer coefficient, Kt, m2/kg).
To compare the 137Cs measurements performed in different years, all data were adjusted to a single date of August 15, 2020.
RESULTS AND DISCUSSON
The studied profiles (Fig. 2) characterize four ELGS: two ELGS of the southern exposition 26 m long with relative elevation range of 2.71 m (profile D-21) and 2.43 m (D-25), and two ELGS of the northern exposition 17 m long with relative heights range of 1.52 and 1.47 m, respectively (Fig. 2).

Relative heights, points of 137Cs activity measurement along two profiles D-25 (a) and D-21 (b) crossing the hollow with indication of ELGS of different exposition (1, 3) and bottom (2), as well as soil core sampling points.
Profile D-21 is located 2 m to the north relative to the profile D-25, the correlation coefficient between heights of the corresponding measurement points accounted for r0.01 = 0.996; n = 26 along ELGS of the southern exposition and r0.01 = 0.995; n = 17 along ELGS of the northern exposition.
Collection of soil cores in separate points with different surface 137Cs activity allowed us to estimate the comparability of results of field and laboratory measurements.
Comparison of Field and Laboratory Measurements of 137Cs Activity
Results of statistical analysis of the laboratory measurements of 137Cs contents in samples of soil and moss cover and field measurements of activity in points of simultaneous sampling of soil and moss are presented in Table 1. According to core measurement data, the total radiocesium inventory in 0–40 cm soil layer varied from 3.2 to 9.11 kBq/dm2 (by 2.8 times), whereas the activity measured by portable gamma spectrometer in core sampling points varies from 115 to 218 imp/s (by 1.9 times). A different scatter in 137Cs variations (wider in core measurements compared to surface measurements) is obviously related to the averaging of activity over a greater area during field measurement compared to the core.
The contamination of moss cover changed by 1.5 times more than that of soil cover, which is caused by specifics of radiocesium transfer in biomass. Thereby, the contamination of moss cover accounted for, on average, 1% of soil contamination density and was no more than 2.4%, i.e., a contribution of plant to the total contamination is insignificant, while radionuclide is mainly restricted to soil (Table 1).
Analysis of vertical 137Cs distribution in the cores showed that 137Cs during 35-year period after primary contamination in undisturbed ELGS weakly migrated in the lower soil layers: 70–96% of its total reserve was fixed in 2020 in the top 8-cm layer of soil, while 0–20 cm layer retained 89–99% of the radionuclide. This fact is confirmed by statistically significant relationship between radionuclide reserve in 0–20 cm soil layer and field measurements of its activity in core sampling points, which provide most complete characteristics of ELGS (along ELGS D21_C, Fig. 6): correlation coefficient accounted for r0.01 = 0.738; n = 9.
Assessment of Variation Pattern of 137Cs activity in the Soil–Plant Cover of ELGS using Fourier Analysis and Harmonics Modeling
Based on the analysis of field measurement data, the 137Cs activity in ELGS showed practically two-fold variations from 113 to 219 imp/s, n = 84. Thereby, the studied ELGS slightly differed in the average values, range, and variation pattern, although ELGS of the southern exposition of profile D-21 showed a range of activity of 1.9 against 1.5 in other ELGS (Table 2), which is obviously related to the more active redistribution of radionuclide with water (this ELGS is located practically along runoff line, Fig. 9).
Analysis of variations of 137Cs activity in each ELGS regardless of slope exposition revealed its systematic cyclic variability within ranges indicated in Table 2, with different amplitude (Figs. 3a, 3b).

Distribution of 137Cs activity in the ELGS of the southern (a) and northern (b) expositions (shown is the topographically lower position of ELGS points of profile D-21 relative to ELGS points of profile D-25).
Spectral Fourier-analysis of the spatial distribution of 137Cs activity in the soil–plant cover made it possible to determine variation periods (harmonnics), which increased with an amplitude growth (Fig. 4).

Fourier analysis of spatial distribution of 137Cs activity in soil-plant cover: ELGS of the (a) southern exposition and (b) northern exposition.
For ELGS of the southern and northern expositions, the variation step of surface 137Cs activity was comparable, with insignificant differences. Periodicity along the entire D-21 profile corresponded to that of its ELGS. In profile D-25, such relationship was observed only within ~ 4-m interval. Periodicity of change of surface 137Cs activity over all points of profile D-25 seemed to be different from that of profile D-21 (Table 3). These differences were explained by solving inverse problem: using harmonics modeling.
The inverse periodicity problem was solved by the using the data of Fourier analysis of surface 137Cs activity. Periodicity was modeled based on three Fourier obtained main harmonics (2.85; 5.45, and 6 m). The initial and modeled variations are illustrated by two plots in Fig. 5. Correlation coefficient between factual variation of surface activity and its model was r0.01 = 0.528, n = 60.

137Cs surface activity distribution (factual data) and its model constructed based on three main harmonics of Fourier-analysis.
Obtained result demonstrates that inclusion of data on ravine bottom in profile in addition to measured surface activity 137Cs along ELGS leads to a significant deviation of model values from true ones along ELGS (Fig. 5). This fact suggests that at study of 137Cs migration and distribution in soil–moss cover, the ELGS should be considered separately from other landforms (Dolgushin and Korobova, 2021).
Analysis of Vertical 137Cs Distribution in the Soil Cover of ELGS
The 137Cs inventory in soils estimated from laboratory 137Cs measurement in cores collected from ELGS D-21_N varied in compliance with the 137Cs contamination density obtained during field measurements (Fig. 6).

Inventory of 137Cs in 0–20-cm soil layer of ELGS of D-21_N (according to soil core data).
This was due to the fact that the main fraction of radionuclide is confined to the upper layers within the depth 4–12 cm (Fig. 7). It should be noted that the total radionuclide vertical distribution pattern in different points of core sampling along ELGS showed no significant differences from that in the closing depressions.

137Cs vertical distribution in soil cores (а–h) of the northern ELGS of profile D-21 (position of cores in relief is shown in Fig. 2).
It is pertinent to mention that some increase of radiocesium content was observed below 8–10 cm, in layers containing organic interbed.
The ELGS closing depression of the southern and northern expositions along profile D-21 sharply differed in the total radionuclide inventory in 40-cm soil layer: 49 Bq/cm2 in core D21-28 and 78 Bq/cm2 in core D21-45. In addition, in the vertical soil profile, practically all radionuclide inventory in the lower part of the ELGS of the steep southern slope was confined to 0–2 cm layer, whereas the main 137Cs inventory in the lower part of ELGS of the more gentle and wetter northern slope is restricted to a layer 4–12 cm, with maximum activity at a depth of 8–10 cm (Figs. 8a, 8b).

137Cs vertical distribution in cores located in the closing depressions of ELGS of the southern (a) and northern (b) expositions.
Such difference can be explained by: (1) essentially higher moisture of ELGS of the northern exposition compared to that of the southern expositions, which is proved by the higher humus content in profile D21-45, (2) position of D-21-28 core on the runoff line (Fig. 9).

Position of runoff line on the test site according to relief (modeling results in ArcGIS).
The total inventory and penetration depth of 137Cs in closing depressions of two other ELGS of different expositions (profile D-25) also appeared to be dissimilar. In the ELGS of the northern exposition, 137Cs in a 40-m-thick layer is lower and confined to the thinner top layer of vertical profile (0–2 cm against 2–6 cm), which is obviously caused by its localization on the purely thalweg runoff line (Figs. 9, 10).

137Cs vertical distribution in cores located in the closing depressions of ELGS of the northern (a) and southern (b) expositions along profile D-25.
Thus, all studied cores show no unambiguous increase of 137Cs in closing depressions of ELGS. The vertical 137Cs distribution in the depressions is affected by surface runoff. For instance, core D21-28 compared to the core D35-36 located higher on the runoff line is characterized by the greater 137Cs activity of the top 0–2 cm layer (30 against 25 Bq/cm2) and its sharper decrease in the lower 2–4 cm layer (5 against 15 Bq/cm2, i.e., six-fold against two-fold decrease, Figs. 8, 10).
Distribution Pattern of 137Cs Activity in the Plant Cover of ELGS its Assessment using Fourier Analysis and Modeling based on Harmonics
The 137Cs distribution in the plant cover of separate ELGS is illustrated in Figs. 11 and 12.

Distribution of 137Cs in the green part of the moss cover of ELGS of the southern (a, D-21_S, D25_S) and northern (b, D-21_N and D-25_N) expositions (the relief for different profiles is reduced to the same height).

Distribution of 137Cs in the rhizoid part of moss cover of the ELGS of the southern (a, D-21_S, D25_S) and northern (b, D-21_N and D-25_N) expositions (the relief for different profiles is reduced to the same height).
The radionuclide content in plant cover of all ELGS showed more significant variations than in soil cover: variation coefficient accounted for 32–54% against 16–30% (Tables 4 and 5). The 137Cs content in the studied components of ground plant cover decreased in the series: rhizoid part of moss > green part of moss > needle–leaf fall (Tables 4–6). Within separate ELGS, it varied twice to five times in the photosynthesizing part of moss, 3–6 times in rhizoid part, and up to 13 times in litter fall.
The 137Cs accumulation in the photosynthesizing part of moss in general corresponded to its content in rhizoid part, which is confirmed by sufficiently high correlation coefficient between radionuclide contents in the indicated parts for both profiles (along D-21—r0.01 = 0.704; n = 60, along D-25—r0.01 = 0.519; n = 57). The highest coefficient was obtained for moss cover from two ELGS: D-25_N (r0.01 = 0.839, n = 16) and D-21_S (r0.01 = 0.790, n = 26). Note that the former ELGS is located on the more wettened slope of the northern exposition, while the latter ELGS is relatively wettened owing to the location practically on the runoff line, which gives grounds to believe that the correlation of 137Cs reserve in green and rhizoid parts of the moss is caused by the water exchange activity between them.
Thus, the 137Cs distribution in the moss cover of ELGS was similar to that of soil cover, although the step and amplitude of variations in some cases were different. No radiocesium accumulation was found both in separate parts of moss cover and in the whole moss cover in closing depressions of ELGS.
To quantify the periodicity of 137Cs variability in moss cover, data on ELGS and on two profiles in general were processed by Fourier analysis. Intervals obtained for the plant cover of ELGS insignificantly differed from those of soil cover (Tables 3, 7).
In general, the periodicity of 137Cs change in moss cover seemed to be comparable both on ELGS (from 2 to 6.50 m) and in all points of profiles crossing the hollow (from 2 to 15 m), which is rather related to the greater significance and similar transfer of radionuclide to plant in each point.
The 137Cs variation in whole moss sample along D-21 profile was modeled based on three main harmonics (2.3, 4.6, and 15.3 m). To compile more reliable model, we used data set of 45 successive measurements due to the sharply elevated 137Cs contents in four samples located in different positions (out of order), which prevented constructing the model of uninterrupted variability on all 60 points. Initial and model data are illustrated as two plots in Fig. 13. The correlation coefficient between factual and model values of surface activity is r0.01 = 0.739 (n = 45).

Distribution of 137Cs content in total moss sample (factual data) and its model constructed using three main harmonics of the Fourier analysis according to data on profile D-21.
Thus, it was shown that the plant cover is characterized by the cyclic 137Cs variations, which makes it possible to estimate its periodicity using input data (harmonics) of Fourier analysis.
Relationship of 137Cs Distribution in the Moss and Soil Cover of ELGS
In absence of current aerial contamination the 137Cs presence in the photosynthesizing part of moss cover indicates radionuclide transfer to plant from contaminated substrate. The coefficients of 137Cs transfer to the moss cover from its inventory in 0–20 cm soil layer based on core sampling varied significantly from 0.009 to 0.048 m2/kg. On average, the transfer value seemed to be comparable with previously obtained value for moss cover of eluvial forested landscape studied in 1994 on the Barki-2 test site, which is also located in the abandoned zone of ChNPP (Korobova, 2006). This indicates a continuous radionuclide circulation in the soil–plant system on the contaminated territories over no less than 25-year period. With a growth of 137Cs inventory in soil layer, its accumulation in the moss cover of ELGS in general decreased. (Fig. 14).

Relations of 137Cs inventory in soil cover 2–4 cm deep (kBq/m2) and its transfer into moss cover (kBq/kg) in core sampling points on ELGS of D-21_S and D-21_N.
The comparison of 137Cs content in the air-dry plant mass and in soil at different depths (according to core data) revealed a significant positive correlation between 137Cs in photosynthesizing part of moss and in the soil layers 8–10 and 10–12 cm deep, i.e., in layers where the most mobile radionuclide fraction penetrates (r0.05 = 0.479 and 0.592, respectively, n = 23), which confirms the probability of pollutant influx in the plant in its most mobile species, since lower layers gain dissolved species.
The transfer of 137Cs from soil to the moss cover was also estimated along ELGS D25_S (Table 8). Kt was estimated from ratio of radiocesium content in plant in mass unit to the radionuclide contamination density in two variants. Variations of Kt and Ka in ELGS D25_S are presented in Fig. 15 with statistical parameters in Table 8.

Transfer and accumulation coefficients of 137Cs, down the slope of ELGS D25_S.
As seen in Fig. 15, these parameters also show cyclic variations in ELGAS, but have different amplitude and not always coincide in frequencies, which is also related to the variability of biomass. In the found published data, the 137Cs transfer coefficient in Pleurozium schreberi collected in Serbia varied from 0.001 to 0.058 m2/kg (Dołhańczuk-Śródka et al., 2011), which in general corresponds to our data in Table 8. The coefficient of 137Cs accumulation by moss Pleurozium schreberi according to the same authors is close to our estimates (from 0.11 to 3.28). According to other data, this coefficient for Hypnum cupressiforme collected from the same area (Gulan et al., 2020b) seemed to be higher and varied within 0.02–8.50, which is higher than our data but lower than data obtained for the same species by (Dragović et al., 2010)—1.01–13.1. The elevated accumulation of the radionuclide in moss Hypnum cupressiforme can be related to the fact that this species is hydrophilic and resistant to contamination.
Calculation of correlation coefficients confirmed the direct relationship between Kt1 and Ka2 r0.01 = 0.878; n = 21. Significant correlation coefficients were also found between Kt 1 and 2 and Ka (r0.01 = 0.756; n = 21 and r0.01 = 0.701; n = 20), which shows their principal comparability.
CONCLUSIONS
Results of our study revealed no unidirected downslope secondary migration of the initially uniform 137Cs fallout both in soil and in moss cover of ELGS. In contrast, the distribution of the radiomarker reveals systematically ordered (cyclic) pattern.
The analysis highlighted a need in more detailed ELGS studies. At that, the technogenic radioactive isotopes provide more detailed insight into spatial redistribution of pollutants in such systems.
The study of regularities of element migration in the ELGS is the key helping optimum solution of several practical tasks, in particular, geochemical monitoring, predicting ion consequences and determinating measures decreasing anthropogenic contamination of the territory and agricultural production, as well as the applying schemes of fertilization decreasing expenses. For this reason, the developed approach to the ELGS study and obtained regularities are of significant interest.
REFERENCES
Yu. V. Aleksiayenak, M. V. Frontasyeva, M. Florek, I. Sykora, K. Holy, J. Masarik, L. Brestakova, M. Jeskovsky, E. Steinnes, A. Faanhof, and K. I. Ramatlhape., “Distributions of 137Cs and 210Pb in moss collected from Belarus and Slovakia,” J. Environ Radioact. 117, 19–24 (2013).
U. Cevik and N. Celik, “Ecological half-life of 137Cs in mosses and lichens in the Ordu province, Turkey by Cevik and Celik,” J. Environ Radioact. 100, 23–28 (2009).
M. De Cort, G. Dubois, Sh. D. Fridman, M. G. Germenchuk, Yu. A. Izrael, et al., Atlas of Radioactive Contamination of Europe by Cesium after Chernobyl Accident (Luxemburg Office for Official Publications of the European Communities, Luxemburg, 1998).
A. Dołhańczuk-Śródka, Z. Ziembik, M. Wacławek, and L. Hyšplerová, “Transfer of cesium-137 from forest soil to moss Pleurozium schreberi,” J. Ecol. Chem. Eng. S. 18 (4), 509–516 (2011).
S. Dragović, N. Mihailović, and B. Gajić., “Quantification of transfer of 238U, 226Ra, 232Th, 40K and 137Cs in mosses of a semi-natural ecosystem,” J. Environ Radioact. 101, 159–164 (2010).
E. Steinnes and O. Njåstad, “Use of mosses and lichens for regional mapping of 137Cs fallout from the Chernobyl accident,” J. Environ Radioact. 21 (1), 65–73 (1993).
M. Florek, M. Frontasyeva, B. Mankovska, K. Oprea, S. Pavlov, E. Steinnes, and I. Sykora, “Air pollution with heavy metals and radionuclides in Slovakia studied be the moss biomonitoring technique,” Ekologia Bratislava. 22, 157–162 (2001).
L. Gulan., T. Jakšić., B. Milenkovic, and J. Stajić, “Elemental concentrations and soil-to-moss transfer factors of radionuclides in the environment of North Kosovo and Metohija,” Sci. Bull. 10 (2), 59–64 (2020).
L. Gulan., T. Jakšić., B. Milenkovic, J. M. Stajic, P. Vasić, Z. Simić, and N. Zlatić, “Mosses as bioindicators of radionuclide and metal pollution in northern Kosovo and Metohija mountain region,” J. Radioanal. Nucl. Chem. 326, 315–327 (2020).
Yu. A. Izrael, Atlas of Radioactive Pollution of European Russia, Belorussia, and Ukraine (Federal. Sluzhba Geodez. Kartograf. Rossii, Moscow, 1998) [in Russian].
E. M. Korobova, “Distribution of cesium and strontium radionulicdes in soil—vegetation cover of landscapes contaminated after the Chernobyl accident,” Geochemistry of Natural and Anthropogenically Transformed Biogeosystems (Nauchnyi Mir, Moscow, 2006), pp. 249–277 [in Russian].
V. G. Linnik, A. I. Kuvylin, V. N. Kuzmichev, and E. M. Korobova, “Organization of databases of radioecogical information on experimental test site in the Bryansk district,” Radiatsiya i Risk. 3, 121–128 (1993).
I. N. Malikova, V. D. Strakhovenko, and B. L. Shcherbov, “Distribution of radionuclides in moss-lichen cover and needles on the same grounds of landscape-climatic zones of Siberia,” J. Environ. Radioact. 198, 64–78 (2019).
M. G. Nifontova, Extended Abstract of Doctoral Dissertation in Biology (IERZh UrO RAN, Perm, 2003) [in Russian].
K. Olson, A. Gennadiyev, and V. Golosov, “Comparison of fly-ash and radio-cesium tracer methods to assess soil erosion and deposition in Illinois Landscapes,” Soil Sci. 173 (8), 575–586 (2008).
S. L. Romanov, E. M. Korobova, and V. L. Samsonov, “Experience in the Application of the upgraded VIOLINIST-III instrument in field radioecological studies,” Yadern. Izmerit.-Inform. Tekhnol. 3 (39), 56–61 (2011).
T. Sawidis, “Uptake of radionuclides by plants after the Chernobyl accident,” J. Environ. Pollut. 50 (4), 317–324 (1988).
Th. Sawidis, G. Heinrich, and M. Chettri, “Cesium-137 monitoring using mosses from Macedonia, N. Greece,” Water Air Soil Pollut. 110, 171–179 (1997).
T. J. Sumerling, “The use of mosses as indicators of airborne radionuclides near a major nuclear installation,” Sci. Total Environ. 35, 251–265 (1984).
D. E. Walling and Q. He, “Improved models for estimating soil erosion rates from cesium-137 measurements,” J. Environ. Qual. 28 (2), 611–622 (1999).
ACKNOWLEDGMENTS
We are grateful to V.Yu. Berezkin, L.I. Kolmykova, and V.S. Baranchukov, the collaborators of the Laboratory of Environmental Biogeochemistry, for the participation in field works. S.L. Romanov is thanked for cooperation in studying the structure of anthropogenic radionuclide-contaminated fields.
Funding
The studies were performed in the framework of the government-financed task of the Laboratory of Environmental Biogeochemistry. Field and laboratory works were partially supported by the Russian Foundation for Basic Research (project no. 19-05-00816).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Translated by M. Bogina
Rights and permissions
About this article

Cite this article
Dolgushin, D.I., Korobova, E.M. Peculiarities of 137Cs Distribution in the Soil–Moss Cover of the Elementary Landscape Geochemical Systems. Geochem. Int. 60, 779–791 (2022). https://doi.org/10.1134/S001670292208002X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S001670292208002X