
Abstract—
The dominant rocks in the area between the village of Kovda and township of Zelenoborskii in northern Karelia, Russia, are amphibole–biotite and biotite plagiomigmatites, which develop after amphibolites. These plagiomigmatites contain epidote that replaces the amphibole and biotite and is found in reaction relations with these minerals. The epidote occurs as thin rims around mineral grains it replaced and in large euhedral crystals. The rocks contain 5–15% epidote. The epidote contains relatively little FeO (no more than 10.57 wt %) but elevated concentrations of CaО (up to 27.53 wt %) and Al2O3 (up to 31.28 wt %), i.e., the mole fraction of the pistacite end member is 0.14–0.22. The mineral crystallized at 400–600°C and 5 kbar. It was formed by metasomatic processes, when components of the replaced minerals interacted with components of the fluid. The amphibole–biotite and biotite rocks in which the epidote crystallized resemble amphibole–biotite and biotite gneisses. Geological, petrographic, and geochemical characteristics of the rocks indicate that they are plagiomigmatites that developed after amphibolites. The paper reports features that enable distinguishing between the plagiomigmatites and similar gneisses. The transformations of the amphibolite at its migmatization starts with the replacement of the hornblende by biotite, up to the complete replacement of the former at a certain progress of this process. Chemically, the original rocks in their migmatization sequence are gradually depleted in Fe, Ti, Mg, and Ca and enriched in Si, Na, and K. The normalized REE patterns of the plagiomigmatites are strongly fractionated: (La/Yb)n = 18.3 for the biotite plagiomigmatite and 30.9 for the biotite–hornblende plagiomigmatite. The rocks of the area occur in a tectonized zone, which is the boundary between two domains in the Belomorian Belt exhumed at 1.8 Ga according to the mechanism of metamorphic cores. This was favorable for the intense inflow of fluids, created a certain pressure (both total and that of H2O + CO2), temperature, and elevated oxidation potential, i.e., conditions required for epidote crystallization.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
INTRODUCTION
The Belomorian Belt is one of the most thoroughly studied structures of the Baltic Shield (much of the territory of this belt was covered by 1 : 10 000 state geological survey and mapped; Kalafati, 1967; Chuikina, 1975), but many aspects of the evolution of this structure remain disputable as of yet. This is, first and foremost, the problem of the exhumation of the deep-sitting rocks and understanding the structural–material transformations associated with this process. Various migmatites and pegmatites in the Belomorian Belt are long though to have been produced by ultrametamorphism, whereas other rocks, which contain such typomorphic index minerals as garnet, cordierite, and kyanite, are believed to have been produced by high-pressure metamorphism. It was not until recently that their genesis was related to the exhumation of the Belomorian complex (Terekhov, 2007). This paper discusses the origin of epidote in rocks of the Belomorian Belt in the context of transformations of the amphibolites during their exhumation. The importance of studying this mineral in the geological structure is dictated not so much by the fact that this mineral was found in the rocks as by that it occurs in the extension structures: the complexes of the metamorphic cores in which epidote may be an indicator of tectonic exhumation (Davy et al., 1989; Chamov, 2016).
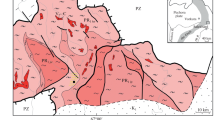
The rocks discussed herein are found in the central, the narrowest, part of the Belomorian Belt east of Kovdozero Lake, which is the boundary between tectonotypic areas (Fig. 1). South of this area, a synclinorium was distinguished in which the Belomorian Group is subdivided into three units (Chuikina, 1975), and the northern part of the belt is thought to be a northeast-dipping monocline (Kalafati, 1967). Our study area has long been considered to be a key point in discussions between proponents and opponents of either of these two structural interpretations.

Principal geological features of the Belomorian Belt and its surroundings. (1) (a) Paleozoic and (b) Proterozoic alkaline intrusions; (2) (a) Late and (b) Early Karelian granites; (3) Early Proterozoic (a) layered intrusions and (b) drusites and anorthosites; (4) (a) Riphean and (b) Karelian rocks; (5) granulites and migmatites developing in them (a) Proterozoic, (b) Archean; (6) kyanite-bearing rocks in (a) the peripheries and (b) central part of the Belomorian Belt; (7) (a) Central Belomorian (CBMZ) and other metamorphic zones and (b) plagiomigmatites and blastomylonites (Khetolambina and Keret complexes); (8) Archean (a) greenstone belts and (b) granite-gneiss basement; (9) Main Belomorian Detachment, which brought the deep rocks to the surface; (10) study area. Roman numerals I and II mark the northern and central domains of the Belomorian Belt (metamorphic cores).
The Belomorian Mobile Belt contains much amphibolites and migmatites developing after them, along with kyanite–garnet–biotite and garnet–biotite plagiogneisses. The greatest number of amphibole units, whose thicknesses vary from a few dozen to a few hundred meters, was found in the Yona and Loukhi areas. In addition to them, various parts of the Belomorian Belt host domains with much amphibolites. One of them was found near the village of Kovda (Figs. 1, 2). The amphibolites commonly occur in association with plagiomigmatites, with the latter replacing the former. The only mafic minerals of the plagiomigmatites are hornblende and biotite, which occur in variable amounts and variable proportions. The plagiomigmatites most often contain biotite alone, whose content amounts to approximately 15%. Sometime rocks in various parts of the belt contain epidote, which is most often viewed (without describing it) as a secondary mineral.

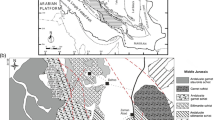
(a) Schematic geological map of the detailed-study area between the 1083 and 1102 kilometer posts at the St.-Petersburg–Murmansk Highway and (b, c) sketches of outcrop fragments. (1) Amphibolites, (2) garnet amphibolites; (3) biotite–hornblende and hornblende–biotite plagiomigmatites; (4) biotite plagiomigmatites; (5) quartz–plagioclase veins; (6) shattered rocks; (7) biotite basificates; (8) strike and dip symbols; (8) (a) faults and (b) structural lines.
The plagiomigmatites developing after amphibolites look like similarly to the metasedimentary gneisses and have an analogous composition. Some geologists believe that the migmatites are gneisses, others classify them with tonalites of the TTG (tonalite–trondhjemite–gneiss) association (Slabunov et al., 2008). According to many other geologists (Beus et al., 1993; Khodorevskaya et al., 2002; Glebovitskii and Sedova, 2005; Levitskii, 2005; Kozlovskii, 2016) and our own data, the amphibole–biotite and biotite rocks are migmatization products of amphibolites.
The first geologists who studied the Belomorian territory interpreted the systematic rocks sequence in the stratigraphic section as a stratigraphic sequence and distinguished a number of formations, with one of them consisting of amphibolites and plagiomigmatites developing after them and referred to as the Khetolambina Formation. Later studies resulted in that the formations were revised and reinterpreted as tectonic nappes (Miller and Mil’kevich, 1995).
As follows from recently acquired data, migmatization of rocks of the Belomorian Belt was a long-lasting process, which began when these rocks were formed at 3.0–2.8 Ga and continued until the drusite (referred to as coronite in the literature in English) complex was formed at 2.45 Ga. The rocks of this complex are only weakly migmatized (Glebovitskii, 2005). The structural setting of intrusions of the drusite complex and the metamorphism of their rocks indicate that they were formed at depths of 20–25 km. The mica-bearing pegmatites were formed at roughly the same depths (at a pressure of P = 7 kbar), mostly at 1.8 Ga (Terekhov, 2007). Starting at that time, the rapid exhumation of rocks of the Belomorian Belt began and was associated with the widespread origin of metasomatic rocks, including those with epidote. It has been demonstrated in many publications that the Belomorian Belt was exhumed to the zone of brittle deformations from depths of 20–25 km at 1800 Ma (Bibikova et al., 1999; Terekhov, 2007). The exhumation of the rocks was associated with metasomatic processes and the origin of such metasomatites as corundum–amphibole–garnet rocks (basificates) and felsic rocks, such as pegmatites, migmatites, and potassic granites.
One of the minerals typical of this evolutionary episode of the deep rocks is epidote, which is believed (Davy et al., 1989; Chamov, 2016) to be a typomorphic mineral of structures formed at regional extension, i.e., metamorphic cores, with which the Belomorian Belt may be classified. Below we present petrochemical characteristics of migmatization of the amphibolites at the exhumation of the rocks and describe in much detail epidote that grew in the plagiomigmatites.
GEOLOGICAL OVERVIEW
In the course of targeted geochemical studies in northern Karelia, rocks were documented in an area approximately 20 km in length in natural outcrops and in roadway excavation along the St. Petersburg–Moscow Highway between the village of Kovda and township of Zelenoborskii (Figs. 1 and 2). Most of the outcrops expose amphibole–biotite and biotite leucocratic rocks, which look like closely similar to the amphibole–biotite and biotite gneisses. Variable amounts of amphibolites typically occur among these rocks. The three rocks show different mutual relations in discrete outcrops. However, if an outcrop is made up of two or three rock varieties, they exhibit mutual transitions both along and across the strikes. The transitions are usually gradual, and the contacts are diffuse. The amphibolites occur among the amphibole–biotite and biotite rocks as relatively narrow stripes or small relics of variable size and shape (Figs. 2b, 2c). Some of the outcrops entirely consist of amphibolites, which are variably migmatized. Some outcrops show that the rocks are deformed in small folds (with amplitudes of the limbs of 20–30 cm). The limbs of these folds consist of clearly distinguishable migmatite, whereas their cores are made up of fine-grained patchy plagiogranite. The patches are composed of aggregates of large biotite flakes, sometimes with amphibole crystals. The melanocratic accumulations were likely formed by basifications. In one of the outcrops, a small ultramafic intrusive body is hosted by pale-colored biotite rocks. The body contains single gedrite crystals.
Pale gray fine-grained fine-flaky homogeneous rocks cropping out 100 m south of kilometer post 1088 contain 8–10% biotite. Chains of small biotite laths define the foliated structure of this rock. The homogeneity of this rock is in places disturbed by the occurrence of thin rare amphibolite beds with diffuse contacts. The rock may have been mylonitized. In the northern part of the area, a number of outcrops consist of fine-grained garnet amphibolite. These rocks are crenulated, with the small folds accentuated by thin stripes of plagiogranite that plagiomigmatized the amphibolite. The rocks are also cut by small pegmatoid veinlets. Near kilometer posts 1092–1094, the amphibolite occur in contact with pale amphibole–biotite and biotite rocks, which are disintegrated, transformed to rubble, and limonitized. The destruction of these rocks was induced by deformations related to the development of relatively long (approximately 3 km long) fault.
Rocks in this area locally host scarce potassic mineralization. The grains of potassic feldspar occur as small porphyroblasts, single large (up to 15 × 10 cm) microcline crystals, and thin veinlets obliquely cutting across the foliation of the rocks.
The amphibole–biotite and biotite rocks contain epidote. Before describing the epidote itself, below we describe how its host rocks were formed.
ORIGIN OF THE AMPHIBOLE–BIOTITE AND BIOTITE ROCKS. PETROGRAPHIC DESCRIPTION
We have studied the transformations of the amphibolites at their plagiomigmatization, starting with the protolithic rocks to granite. The protolithic rock is amphibolite, which contains 50–60% amphibole, 30–40% plagioclase, and about 10% quartz. The amphibole is common hornblende close to the hastingsite isomorphic series (Boyd, 1961). The plagioclase is andesine (An 34–36). The amphibole is medium-grained, homogeneous, nematogranoblastic rock. The transformations of the amphibolite start with rock recrystallization and the crystallization of single thin biotite platelets. The amphibolite is weakly biotitized. Later, during the first stage of migmatization, hornblende is more actively replaced by biotite and shows nonlinear contacts with it. During this stage, small euhedral biotite platelets are formed, which developed as aggregates with hornblende along even lines, without evidence of corrosion. This biotite belongs to the neosome. The index of refraction of this biotite is higher than that of biotite in the biotitized amphibolite (nm = 1.647 and nm = 1.633, respectively). The hornblende also occurs in two populations. One of them comprises prismatic crystals up to 1.2–3.3 mm long or equant grains with uneven corrosion outlines because of their replacement not only by biotite but also by plagioclase and quartz. This is the hornblende of the protolithic rocks. The other variety is found as small (0.4–0.6 mm) euhedral bluish green grains in the neosome. This hornblende is richer in iron than the previous variety (ng = 1.691, np = 1.673, 2V = 65° and ng = 1.674, np = 1.656, 2V = 76°, respectively). The plagioclase occurs as grains of variable size and shape. Roundish or polygonal grains up to 1.5–2.0 mm occur together with anhedral grains or small tablets with polysynthetic twins. The tablets consist of andesine (An 27). The quartz forms small (0.3–0.5 mm) anhedral grains. The first migmatization stage of the amphibolite is thus marked by the origin of biotite–hornblende migmatite containing 20–25% hornblende and 10–15% biotite. The neosome of this migmatite contains the stable mineral assemblage Hbl + Bt + Pl + Qz. The microstructure of this rock is lepidonematogranoblastic.
The progress in the migmatization processes results in hornblende–biotite plagiomigmatite, in which biotite dominates over hornblende. The further evolution of the processes gives rise to plagiomigmatite. This rock is widespread in the study area. The stable mineral assemblages in the neosome is Bt + Pl + Qz. The rocks either do not contain any protolithic material at all or contain only its relics. The rocks usually consist of minerals of the previous stages that differ from the neosome minerals in morphology and in possessing a lower Fe mole fraction, and the plagioclase is more calcic. The biotite plagiomigmatites are fine-grained lepidogranoblastic rocks. Biotite most commonly occurs in the plagiomigmatites as small euhedral platelets 0.4–0.6 mm long. Along with them, the rocks contain large biotite platelets or their fragments, which sometimes host vermicular quartz inclusions. This is biotite that replaced the amphibole. It can be seen in several thin sections that the two biotite populations occur in various mutual relationships, although the plagiomigmatites of this stage typically contain small biotite crystals evenly distributed in the rock. The average content of this mineral is 12–15% but may locally increase to 20%. The plagiomigmatites contain about 60% plagioclase on average. The plagioclase comprises a number of its genetic types. One of them is only occasionally found in thin sections: these are small plagioclase tablets and prisms with rounded outlines, 0.8 × 0.4 mm, intensely saussuritized. Their composition is andesine (An 34–36). This plagioclase is scarce in the rocks and is relics inherited from the protolithic rocks. The plagiomigmatites of this stage mostly commonly contain subequant or irregularly shaped plagioclase grains 0.5 × 0.3 to 2.0 × 1.0 mm, corresponding to oligoclase (An 27–25). This plagioclase is unevenly and insignificantly saussuritized. The plagiomigmatites contain variable quartz concentrations, which never exceed 20%. The quartz grains are anhedral, 0.2–0.6 to 3 × 2 mm. The single large quartz grains host small plagioclase inclusions and small apatite crystals. The large quartz grains were likely formed in relation to the later silification of the rocks.
The newly formed amphibole–biotite and biotite plagiomigmatites resemble the amphibole–biotite and biotite gneisses. The rocks can be reasonably reliably distinguished and identified based on their geological relations with the amphibolites and on petrographic features. The plagiomigmatites exhibit features that are atypical of the gneisses: (a) they contain relatively little quartz, less than 20%; (b) the biotite plagiomigmatites contain relict amphibole grains; (c) the rocks host quartz symplectites in biotite; (d) the rocks contain biotite of two types (protolithic and neosomic), whose Fe mole fractions are different; and (e) in the amphibole–biotite plagiomigmatites, amphibole was replaced by biotite and its contents vary.
Migmatization of the amphibolite sometimes ends with the origin of biotite plagiomigmatite but sometimes continues and is associated with more profound transformations of the protolith. This marks the incipient recrystallization of the biotite plagiomigmatite in the form of thin bands and patches of irregular shape. Biotite occurs between them as thin discontinuous bands. The rock is heterogeneous nebular plagiomigmatite. Its plagioclase grains vary in size, shape, and composition, with the dominance of oligoclase (An 17–20). The transformations of the amphibolite during its migmatization end with the origin of biotite granite. The granite is not widespread in the area, and its origin seems to have been separated in time from the origin of the plagiomigmatites.
The process of amphibolite migmatization is associated with an increase in the Fe mole fraction of amphibole and biotite in the neosomes of the migmatites and a systematic increase in the total Fe mole fraction of the rocks in the sequence from the protolith to granite.
EPIDOTE IN THE ROCKS DEVELOPING AFTER AMPHIBOLITES: CHARACTERISTICS OF EVOLUTION
In the plagiomigmatites, epidote occurs near amphibole and biotite, and hence, its distribution depends on the distributions of these minerals. As seen in thin sections, the epidote replaces both amphibole and biotite, with biotite replaced most intensely. The distribution of epidote in the rocks is uneven, and its content ranges from 2–7 to 10–15%. The mineral occurs as small grains along the margins of biotite and amphibole grains, with corrosion outlines (Figs. 3a, 3b). Some biotite and amphibole grains are almost completely replaced by epidote, with only small relics of these minerals remaining and showing embayed corrosion outlines (Fig. 3b). Often, the development of the epidote starts with thin rims along the margins of amphibole and biotite grains (Fig. 3c). Note the large skeletal epidote grain in this photo and the euhedral epidote crystal (well faceted hexagon) that hosts an unreplaced part of an amphibole grain. A nearby prismatic epidote crystal hosts a biotite relic, which has (similar to the aforementioned amphibole relic) torn outlines. The biotite relics are sometimes hosted in large euhedral epidote crystals (Fig. 3d). Some faces of these crystals exhibit small sawtoothed outlines because of biotite replacement (Fig. 3e). The epidote grains often contain quartz symplectites. Some of the grains partly consist of epidote and partly of biotite with symplectites in both minerals. Symplectites in the epidote are quartz that was released when the epidote replaced biotite, whereas symplectites in the biotite were formed when amphibole was replaced by the biotite. An epidote grain hosted a biotite relic, which hosted an amphibole relic. Such relations between minerals confirm not only that the epidote replaced biotite but also that the biotite replaced amphibole, pointing to a “migmatization relic”.

Microphotographs (plane-polarized light, one polarizer) of thin sections showing the replacement of amphibole and biotite by epidote in the plagiomigmatites: (a) epidote replaces biotite and occurs in corrosive contact with it (thin section 3/11a); (b) epidote hosts amphibole relics with embayed outlines (thin section 13/11-2a); (c) thin epidote rims around amphibole, a skeletal; epidote grain with a biotite relic, and a six-rayed aggregate of epidote crystals with an amphibole relic (thin section 13/11-2); (d) biotite relics in a large epidote crystal and (bottom left) a nonlinear contact of this epidote crystal with biotite (thin section 9/11a); (e) sawtooth-shaped contact (right) of an epidote crystal and biotite relic (thin section 51).

Chondrite-normalized REE patterns of the amphibolite and rocks replacing it (Shcherbakova, 1988). See Table 1 for rock names.
The epidote occurs as grains of variable shape and size. Most commonly its grains are 1.0–1.2 × 0.4–0.6 mm, although the rocks also contain both smaller epidote grains and larger ones, up to 2.5 mm long. The epidote is colorless in thin sections, but some of its grains possess a pale greenish yellow color. Many sections of the epidote grains display clearly seen cleavage. The interference colors vary from yellow and pale blue to bright blue with bright pink patches. The birefringence of epidote grains in a single thin section varies from 0.010 and 0.015 to 0.049. According to (Lodochnikov, 1974), the composition of epidote and, hence, its optic properties can vary even in a single thin section. The cores of epidote grains usually host oval orthite inclusions. These inclusions are orange-brown and have low birefringence colors. The epidote and biotite are often found in association with sphene and relatively much apatite.
The epidote-group minerals belong to the orthorhombic and monoclinic systems. Until recently, this group was thought to comprise the following minerals (Deer et al., 1978): zoisite, clinozoisite, epidote, piedmontite, and allanite. A nomenclature of the minerals of the epidote group was proposed in 2006 (Armbruster et al., 2006) by an international group of researchers based on a great number of analyses of these minerals. This nomenclature suggests not to classify zoisite with the epidote group as an orthorhombic polymorph of clinozoisite, and the following three subgroups are suggested to distinguish: clinozoisite, allanite, and dollaseite. Depending on its composition, the mineral is classed with one of these groups.
PRINCIPAL GEOCHEMICAL FEATURES OF THE ROCKS AND MINERALS
Rocks
Major components. Below we present analyses of rocks corresponding to their sequence formed at amphibolite migmatization, from the protolith to migmatite (Table 1). As seen in the table, even the initial amphibolite transformations into biotite–hornblende plagiomigmatite are associated with the depletion of the original rock in FeO, Fe2O3, MgО, CaО, and TiO2 by 1.2–1.5 times. For example, the MgO concentration therewith decreases from 8.4 to 5.3 wt %, and that of CaO decreases from 9.9 to 6.1 wt %. This behavior of the components is caused by the partial replacement of hornblende by biotite, plagioclase, and quartz. The further stages of amphibolite transformations are associated with a further decrease in the concentrations of the aforementioned components. Simultaneously, at the earliest migmatization stages, the biotite plagiomigmatites become enriched compared to the protolithic amphibolite in K2O (2.74 wt % versus 1.81, 1.35 wt %). In the absence of K-feldspar in the rocks, the increase in the K2O concentration is explained by the crystallization of biotite. The biotite plagiomigmatites contain no more than 5% biotite and 0.5–1.0 wt % K2O. The K2O concentration increases to 4.38 wt % only in the final rocks in the sequence of amphibolite transformation products: in the microcline granite. The Na2O concentration at the first stage, in the biotite–hornblende plagiomigmatite, increases insignificantly but continues to systematically increase at each of the successive stages and reaches 3.86–5.20 in the plagiogranite, which is two to three times higher than in the protolithic amphibolite. This behavior of Na2O is explained not only by the increase in the plagioclase content but also by the decrease in its CaO concentration. The biotite plagiomigmatites and granites, i.e., the final migmatization products, are thus significantly depleted in FeO, MgO, CaO, and TiO2, for example, the Fe2O3 concentration is 0.74–0.01 wt %. Simultaneously the rocks become notably enriched in SiO2, Na2O, and K2O. This behavior of components in the rocks corresponds to silicic–alkaline metasomatism.
Rare-earth elements (REE). The distribution of REE was analyzed in the rock sequence corresponding to migmatization of the amphibolite, from the protolithic rock to biotite granite (Table 1). In the amphibolite, REE are weakly fractionated, (La/Yb)n = 0.75, and depleted in LREE. The successive members of the migmatization rock sequence are enriched in LREE and depleted in HREE compared to the protolithic amphibolite. The LREE concentrations particularly significantly increase at incipient migmatization, in the biotite–hornblende plagiomigmatite, in which La concentration is six times higher than in the amphibolite. Data on REE concentrations in the rock sequence corresponding to migmatization indicate that these elements are strongly fractionated. The (La/Yb)n ratio changes from 18.3 in the plagiogranite to 30.9 in the biotite–hornblende plagiomigmatite.
Minerals
Biotite is the main mafic mineral in rocks of the amphibolite migmatization sequence. The composition of the biotite was analyzed in two biotite plagiomigmatite samples, and the stoichiometric coefficients of cations were calculated (Table 2). The biotite is dark bright brown, likely because of the high TiO2 concentrations (2.42–2.82 wt %). The highest temperature biotite is the richest in TiO2. However, Ti concentration in biotite depends on its Fe mole fraction and is at a minimum in medium-temperature biotite (Korikovsky, 1967). Biotite in the rocks has XFe = 61–69%. As seen in Table 2, biotite of higher Fe mole fraction bears higher TiO2 concentrations.
Epidote. The epidote was analyzed in fifteen grains in two samples of the biotite plagiomigmatite.
As seen in this table, the epidote contains relatively little Fe (7.27–10.57 wt %, and this mineral in three samples contains less than 5% Fe recalculated to FeO). According to the nomenclature recommended for the minerals of the epidote group (Armbruster et al., 2002), some of the analyzed grains, which are richer in Fe, can be classified with epidote, and the others are clinozoisite. Epidote and clinozoisite are not divided by a clearly defined boundary, all of their properties vary gradually and in direct correlation with the Fe mole fraction of the minerals. Some researchers (Kepezhinskas and Khlestov, 1971) employed the Fe/(Fe + Al) ratio as the Fe mole fraction of epidote. This ratio is referred to as the pistacite component in the foreign literature (Brunsmann et al., 2002). This ratio in most of the analyzed epidote grains is 0.14–0.22 and decreases to 0.07–0.09 in grains poor in Fe (Table 2).
We have also analyzed the composition of the accessory apatite and sphene. Both minerals and epidote contain relatively much calcium: 55–58 wt % CaO in apatite and 27.76 wt % CaO in sphene.
DISCUSSION
The process of rock migmatization, including the migmatization of amphibolites in Belomorie, is discussed in analyzed in numerous publications (Beus et al., 1993; Glebovitsky and Sedova, 2005; Levitskii, 2005; Kozlovskii, 2016). The processes has also been studied experimentally (Khodorevskaya et al., 2002). The authors of these publications often invoked different mechanisms to form migmatites and granites. Migmatized rocks of particular interest are the amphibole–biotite and biotite plagiomigmatites developing after amphibolites because these migmatites resemble amphibole–biotite and biotite gneisses. Because plagiomigmatites developing after amphibolites are widespread among Belomorian rocks, it is important to correctly identify them. Many outcrops in the area expose plagiomigmatites developing after amphibolites and containing epidote. According to literature data, epidote is found in rocks of various genesis: metamorphic, metasomatic, and magmatic. This mineral is believed to be most typical of metamorphic rocks of low and intermediate grades of regional metamorphism of the greenschist (T = 250–550°C, Р = 1.5–3–4 kbar) and epidote-amphibolite (T = 500–650°C, Р = 3.5–6 kbar) facies. In greenschist-facies rocks, epidote occurs in association with actinolite, chlorite, and albite. The typical minerals of the epidote-amphibolite facies are epidote, biotite, oligoclase, and sometimes albite. They are often regarded as stress minerals but crystallized at low temperatures (Deer et al., 1978). Many researchers (particularly abroad) describe epidote crystallization in magmatic rocks and present data on modeling the magmatic crystallization of epidote. Magmatic epidote was described in granodiorite in the Mesozoic accretionary terrane between northern California and southeastern Alaska (Zen and Hammarstrom, 1984). These authors have modeled epidote crystallization in synthetic granodiorite melt at P = 8 kbar and believe that plutonic rocks with epidote shall crystallize under moderately high pressures corresponding to the lower crust. V.N. Smirnov and E.A. Zin’kova (Smirnov and Zin’kova, 1993) summarized their own results on magmatic epidote crystallization and compared these and experimental data of other authors. This led them to the conclusion that magmatic epidote can crystallize only under a high total pressure and elevated water content in the melt. For example, the total pressure at the crystallization of the epidote-bearing Verkhneisetskii massif in the Urals was evaluated at 4–5 kbar at a water pressure of 8 kbar (Smirnov and Zin’kova, 1993).
Epidote is known to be often formed by contact-metasomatism processes in skarns. The experimental modeling of the origin of epidote-bearing skarns indicates that these rocks crystallize at temperatures of 350–500°C, at elevated KCl/NaCl ratios in the solutions, and at elevated Eh and Fe activity in the solutions (Ezhov and Zaraiskii, 1993).
Epidote is often described in the literature as a metasomatic mineral, which often occurs in unusual forms. Epidote in the form of ovoids was locally found in the Magadan batholith in northeastern Russia. The ovoids are zoned and as long as 50 cm. According to (Firsov, 1959), the epidote was formed by metasomatic processes, under the effect of fluid that introduced Ca and other components.
Only a few publications describe in detail epidote in the Belomorian Belt, although even the very first researchers of this structure documented epidote in biotite gneisses and amphibolites in some parts of the belt: in the Kaita-Tundra, Vad-Ozero, and Kovdozero areas (Shurkin et al., 1962; Duk, 1967). These authors did not present a detailed descriptions of the epidote, and this mineral is referred to as an additional phase to the main mineral assemblage of the rocks. No rocks metamorphosed to the epidote-amphibolite facies were described by these researchers in the area. At geological survey and mapping in western Belomorie, zoisite blastolites were found, in which zoisite and clinozoisite occur in association with labradorite (Duk, 1967). Petrographic descriptions of rocks from the Engozero and Kovdozero domains of the belt (Kozlov et al., 2006) and from the North Keret and Tikshozero greenstone belts (Slabunov et al., 2007) mention that epidote was identified in the biotite and biotite–amphibole gneisses and amphibolites as a secondary mineral.
Secondary minerals are believed to replace primary ones under the effect of weathering, hydrothermal solutions, or fluids coming from deep levels. These minerals typically crystallize in fracturing and shattering zones and in the intergranular space. Their grains are usually small, and these minerals are rich in OH and H2O (Lodochnikov, 1974). Zoisitites were studied in much detail in eastern Belomorie (Slabunov et al., 2015). These rocks occur as lenses of various size among granite-gneisses. According to these authors, zopisitites developed after anorthosites at eclogite-facies metamorphism (T = 750°C, P = 20 kbar). Epidote was also documented in corundum-bearing metasomatites of the Belomorian Belt as a mineral of the metasomatic zoning (Terekhov, 2007).
In the course of fieldwork, we found epidote in plagiogranites of the Keret Formation. Its small pistachio-green grains compose, together with amphibole and biotite, relatively small roundish accumulations disseminated in the rocks. The epidote replaced biotite, which in turn replaced amphibole. The growth of epidote is facilitated by the abundance of calcium, which is released when plagioclase becomes less calcic in the plagiogranites. Epidote grows metosamatically. Its content in the thin sections does not exceed 0.6%. The epidote itself and the rocks replacing amphibolites in which this mineral occurs were formed by metasomatic processes, when mineral components are replaced under the effect of endogenic solutions (Korzhinskii, 1956). The diffusion-controlled transport of material at metasomatism proceeds through pores in the rocks and seems to be the most realistic in light of the fact that the rock temperature was not changed when the epidote was formed. At diffusion-controlled metasomatism, “old” components are dissolved practically simultaneously with the introduction of “new” ones, and the rock remain solid.
The stable mineral assemblage of the neosome of the biotite plagiomigmatites is Bt + Pl + Qz, and that of the amphibole–biotite plagiomigmatites is Hbl + Bt + Pl + Qz. In these rocks, epidote replaces amphibole and biotite, and its relationships with the replaced minerals are always corrosive. At a certain stage of epidote formation, its large single euhedral crystals are produced. Some of their crystal faces in contact with biotite relics also show corrosive relations with biotite. The amphibole and biotite are replaced at reaction interaction of their components with fluid, and metasomatic epidote is formed. This process is associated with silica release, and this silica forms symplectites in the epidote. The occurrence of epidote in the biotite plagiomigmatites may lead geologists who believe that these migmatites are gneisses to misinterpreting these rocks are those progradely metamorphosed to the epidote-amphibolite facies. However, the epidote would then have occurred as an individual rock-forming mineral and would have not showed corrosive relations with other minerals, which is not the case. Evidence that epidote can crystallize, without any indications of corrosive relations with other minerals, at prograde metamorphism is provided by materials of other geologists. For example, the rocks of the Stanovoi complex (mostly amphibolites and plagiogneisses), which were metamorphosed to the epidote-amphibolite facies at temperatures of 600–650°C, contain epidote as the main rock-forming mineral. These rocks also contain biotite, hornblende, almandine, muscovite, kyanite, and albite–oligoclase. None of these minerals was found in reaction relations with the epidote. The epidote occurs as individual euhedral crystals ranging from a few fractions of a millimeter to 1.5–3.0 mm (Kozyreva and Romanenko, 1978).
Another example of epidote in biotite plagiomigmatites as a mineral produced by prograde metamorphism is as follows. It is widely known that the great majority of rocks of the Belomorian complex are metamorphosed to the amphibolite facies. Had the metamorphic processes resulted in the exhumation of the rocks and their subsequent retrogression and the origin of epidote in the biotite rocks, then according to this scenario, solutions responsible for the retrogression would have formed not only epidote but also other OH-bearing minerals. The hornblende would have been replaced by tremolite and chlorite, the biotite would have been chloritized, and epidote would have been formed together with albite at plagioclase decomposition. However, the rocks show no evidence of these processes. The most likely scenario for the origin of this epidote is thus its metasomatic genesis in plagiomigmatite.
The crystallization temperature of epidote lies within the range of 400–600°C at a pressure of 5 kbar. This temperature is inferred from the diagram for zoisite and clinozoisite (Brunsmann et al., 2002) with pistacite isopleths in epidote (Fig. 5). According to data acquired by many scientists, factors important for the origin of epidote are, in addition to certain temperatures and total pressures, also H2O and CO2 pressure, Eh, Fe activity, etc.

T–Хps diagram for coexisting zoisite and clinozoisite: (1) pistacite end member of the epidote described herein, (2) pistacite composition fields according to (Brunsmann et al., 2002).
CONCLUSIONS
The plagiomigmatites replacing amphibolites in the study area contain 5–15% epidote, which replaces amphibole and biotite. The epidote occurs in a great diversity of forms, from small grains and thin rims around the replaced minerals to large euhedral crystals. The epidote shows corrosive contacts with the replaced minerals.
The epidote is relatively poor in Fe and rich in Al and Ca and contains 0.14–0.22 of the pistacite component. The epidote is of metasomatic genesis and was formed when amphibole and biotite components interacted with fluid components at 400–600°C and 5 kbar.
Most rock outcrops at the study area expose pale amphibole–biotite and biotite rocks, which often host amphibolites in the form of narrow bands and relict domains. Considered together, geological, petrographic, and geochemical features of these rocks indicate that their amphibole–biotite and biotite varieties are products of amphibolite migmatization, i.e., are plagiomigmatites.
Analysis of the chemical composition of the rocks and the sequence of transformation products of the amphibolites during their migmatization, from the protolithic amphibolite to biotite granite, suggests that this process was associated with the removal of Fe, Ti, Mg, and Ca from the rocks and introduction of Si, Na, and K into them. These relations between components in the rocks correspond to siliceous–alkaline metasomatism. The origin of metasomatic epidote in the plagiomigmatites can be paralleled with basification. Basification coupled with migmatization can be associated with epidote crystallization, along with the origin of monomineralic aggregates of amphibole and biotite, which are found in the plagiomigmatites. It is highly probable that components released at migmatization are utilized in this process.
The metasomatic growth of apatite in rocks in the study area may be explained by the widespread tectonic processes that accompanied rock exhumation, occurred in Belomorian Belt at 1.8–1.75 Ga, and was related to the development of downdip–strike-slip fault zones between the northern and central domains of the Belomorian Belt. These domains can be viewed as independent metamorphic cores. These processes could be favorable for the intense fluid inflow and result in P–T parameters suitable for epidote formation.
REFERENCES
T. Armbruster, P. Bonazzi, M. Akasaka, V. Bermanec, C. Chopin, R. Giere, S. Heuss-Assbichler, A. Liebscher, S. Menchetti, Y. Pan, and M. Pasero, “Recommended nomenclature of epidote-group minerals,” Europ. J. Mineral. 18 (5), 551–567.
A. A. Beus, T. F. Shcherbakova, and L. N. Kuklei, “Geochemical features of granitization and average composition of rocks of the Belomorian Complex,” Geochemical Evolution of Granitoids in the Lithosphere History, Ed. by A. A. Beus (Nauka, Moscow, 1993) [in Russian].
E. V. Bibikova, A. I. Slabunov, S. V. Bogdanova, and T. Skiold, “Early Precambrian tectono-thermal evolution of the Earth crust in the Karelian and Belomorian provinces of the Baltic Shield: U–Pb isotopic evidence from sphene and rutile,” Geochem. Int. 37 (8), 750–764 (1999).
I. D. Borneman-Starynkevich, Guidebook on Calculation of Mineral Formulas (Nauka, Moscow, 1964) [in Russian].
F. R. Boyd, “Hydrothermal investigations of amphibolites,” Researches in Geochemistry, Ed. by P. H. Abelson (Wiley, New York, 1959) pp. 377–396.
A. Brunsmann, G. Franz, and W. Heinrich, “Experimental investigation of zoisite–clinozoisite phase equilibria in the system CaO–Fe2O3–Al2O3–SiO2–H2O,” Contrib. Mineral. Petrol. 143, 115–130 (2002).
N. P. Chamov, Structure and Evolution of the Middle Russian–Belorussian Province in the Neoproterozoic (GEOS, Moscow, 2016) [in Russian].
E. P. Chuikina, “Structure and pegmatite abundance in North Karelia,” in Muscovite Pegmatites of the USSR (Nauka, Leningrad, 1975), pp. 153–159 [in Russian].
P. Davy, G. Guerin, and J. P. Brun, “Thermal constrains on the tectonic evolution of a metamorphic core complexes (Santa Catalina Mountains),” Earth Planet. Sci. Lett. 94. 425–440 (1989).
W. A. Deer, R. A. Howie, and J. Zusman, Rock-Forming Minerals (Longman, London, 1963), Vol. 1.
V. L. Duk, Folds in Ultrametamorphic Zones (Nauka, Leningrad, 1967) [in Russian].
S. V. Ezhov and G. P. Zaraisky, “Experimental modeling of epidote-bearing skarns,” Experiment in Mineralogy, Ed. by M. B. Epel’baum (Moscow-Chernogolovka, 1993), pp. 136–157 [in Russian].
L. V. Firsov, “Metasomatic epidite ovoids in porphyrites, Proc. All-Union Research Institute of Gold and Trace Metals, Ed. by N. A. Vanshedt (Magalan, 1959), Vol. 14, pp. 117–124 [in Russian].
V. A. Glebovitsky, Early Precambrian of the Baltic Shield (Nauka, St. Petersburg, 2005) [in Russian].
V. A. Glebovitsky and I. S. Sedova, “Specifics of Late Paleoproterozoic ultrametamorphism in the Belomorian allochthon,” Dokl. Earth Sci. 401 (1), 267–271 (2005).
L. V. Kalafati, “Method of geological mapping of Precambrian in the Ena and Strel’ninsky areas of the Murmansk oblast,” Problems of Study of Precambrian Geology (Nauka, Leningrad, 1967), pp. 99–103 [in Russian].
K. B. Kepezhinskas and V. V. Khlestov, Statistical Analysis of Epidote-Group Minerals and their Paragenetic Types (Nauka, Moscow, 1971) [in Russian].
L. I. Khodorevskaya, V. M. Shmonov, and V. A. Zharikov, “Granitization of amphibolite: experimental modeling at 750°C and 5 kbar pressure,” Dokl. Earth Sci. 383 (2), 218–221 (2002).
S. P. Korikovsky, Metamorphism, Granitization, and Postmagmatic Processes in the Precambrian of the Udokan–Stanovoy Zone (Nedra, Moscow, 1967) [in Russian].
D. S. Korzhinskii, “Essay of metasomatic processes,” Main Problems in the Concept of Magmatogenic Ore Deposits (AN SSSR, Moscow, 1955), pp. 332–452 [in Russian].
N. E. Kozlov, N. O. Sorokhtin, and V. N. Glaznev, Archean Geology of the Baltic Shield (Nauka, St. Petersburg, 2006) [in Russian].
V. M. Kozlovskii, “Physicochemical model for the genesis of Svecofennian plagiomigmatites of the Belomorian complex, northern Karelia,” Geochem. Int. 44 (9), 855–866 (2006).
V. M. Kozlovsky, and Y. Y. Bychkova, “Geochemical evolution of amphibolites and gneisses of the Belomorian mobile belt during Paleoproterozoic metamorphism,” Geochem. Int. 54 (6). 529–542 (2016).
I. V. Kozyreva and I. M. Romanenko, “Epidote-group minerals in high-grade rocks of the Stanovoy Complex,” Minerals and Parageneses of Minerals, Ed. by T. N. Tokareva (Nauka, Leningrad, 1978), pp. 131–140 [in Russian].
V. I. Levitsky, Petrology and Geochemistry of Metasomatism during Formation of Continental Crust (Akad. Nauk, Novosibirsk, 2005) [in Russian].
V. N. Lodozhnikov, Major Rock-Forming Minerals (Nedra, Moscow, 1974) [in Russian].
Yu. V. Miller and R. I. Mil’kevich, “Thrust-fold structure of the Belomorian zone and its correlation with the Karelian granite–greenstone terrane,” Geotektonika, No. 6, 80–92 (1995).
A. R. Prunier and D. A. Hewitt, “Experimental observations on coexisting zoisite–clinozoisite,” Am. Mineral. 70, 375–378 (1985).
T. F. Shcherbakova, Amphibolites of the Belomorian Complex and their Granitization (Nauka, Moscow, 1988) [in Russian].
K. A. Shurkin, N. V. Gorlov, and M. E. Sal’e, Belomorian Complex of North Karelia and Southwestern Kola Peninsula. Geology and Pegmatite Abundance (AN SSSR, Moscow–Leningrad, 1962) [in Russian].
A. I. Slabunov, Geology and Geodynamics of Archean Mobile Belts: Evidence from the Belomorian Province of the Fennoscandian Shield (Karel’sk. Nauchn. Ts. RAN, Petrozavodsk, 2008) [in Russian].
A. I. Slabunov, O. I. Volodichev, Syao Li, and O. A. Maksimov, “Archean zoisitites of the Gridino eclogite-bearing mélange (Belomorian province): geology, U-Pb age of zircons, and geodynamic implications,” Tr. Karel’sk. Nauch, Ts. RAN 14, 85–105 (2015).
V. I. Smirnova, And E. V. Zin’kova, “Magmatic epidote in the granitoids of the Verkhneset massif (Middle Urals),” Dokl. Ross. Akad. Nauk 329 (3), 332–335 (1993).
E. N. Terekhov, “Lapland–Belomorian mobile belt as an example of the root zone of the Paleoproterozoic rift system of the Baltic Shield,” Litosfera, No. 6, 15–39 (2007).
E. N. Terekhov, “REE distribution in corundum-bearing and other metasomatic rocks during the exhumation of metamorphic rocks of the Belomorian Belt of the Baltic Shield,” Geochem. Int. 45 (4), 364–380 (2007).
E-an Zen and J. M. Hammarstrom, “Magmatic epidote and its petrologic significance,” Geology 12 (19), 515–518 (1984).
Funding
This study was carried out under a government-financed research project for the Geological Institute, Russian Academy of Sciences.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Translated by E. Kurdyukov
Rights and permissions
About this article

Cite this article
Shcherbakova, T.F., Terekhov, E.N. & Kuklei, L.N. Metasomatic Epidote in Amphibolite-Replacing Rocks: An Indicator of Tectonic Processes Related to the Exhumation of Rocks of the Belomorian Belt. Petrography and Geochemistry. Geochem. Int. 59, 1157–1169 (2021). https://doi.org/10.1134/S0016702921120053
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0016702921120053