
Abstract
Previously, in male Greenfinch, we have described ultradian rhythms of body temperature with periods of 3–6, 8–10, 12–20, and 40–60 min. There are individual variations in this rhythmicity. Rhythms with a period of 3–7 min are more characteristic of some males but not for the others. These males had small values of the ratio of the harmonic spectral density in the range of 12–20 min to the harmonic spectral density in the range of 3–7 min. In this study, we hypothesized that ultradian rhythmicity of body temperature fluctuations is related to the nervous system activity in general and to temperament in particular. We have conducted two sets of experiments for estimating the aggression level (n = 12 males) and reaction to novelty (n = 17). Individuals with predominant rhythms with a period of 3–7 min are less aggressive and more slowly adapt to new conditions. Thus, the ultradian body temperature rhythmicity in Greenfinch is related to their behavioral features and temperament.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Cyclicity is typical of many natural processes. It often coincides with the cyclicity of abiotic factors: e.g., seasonality or alternation of the dark and light times of the day. However, ultradian rhythms with a period less than 24 h are also known. They are less studied; their functional significance is less clear [1]. The manifestations of ultradian rhythmics can affect, e.g., the level of hormones in the blood, physical activity, heart rate [2–4], as well as body temperature.
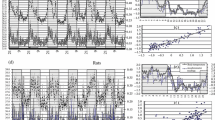
We have previously studied body temperature in greenfinch (Chloris chloris) [5]. This species has a temperature 4–5°C lower at night than during the day (Fig. 1a). During the day, ultradian rhythms with an amplitude of less than 0.3–0.5°C were revealed (Fig. 1b). In most males, these rhythms are grouped in several frequency ranges; their periods are about 40–60, 16–20, 11–13, 6–8, and 3–5 min. The amplitude of the rhythms has individual variability. In particular, some males were distinguished by pronounced 3- to 7-min rhythms (red line in Fig. 1b).

Dynamics of the body temperature of greenfinches (a) during one day and (b) for two daytime hours of the same day. The data are given for two males, with a short (red line) and an extended (blue line) yellow area on the rectrix.
The body temperature dynamics in the range of about 3–20 min in both animals and humans is probably determined by the tone of the autonomic nervous system: both sympathetic and parasympathetic inhibition leads to the disappearance of oscillations [6]. One of the effective physiological methods for assessing the regulatory effects of the autonomic nervous system, in particular on the heart, is heart rate variability [7]. This method is used not only to study the activity of the cardiovascular system, but also to assess the adaptive capabilities of the whole organism, which is reflected in the balance of sympathetic/parasympathetic responses. An important indicator in the method of assessing heart rate variability is the ratio of high and low frequency rhythmic changes in the time intervals between heartbeats. We assumed that a similar technique for analyzing rhythmic changes in body temperature in a lower frequency range of periods can be informative for assessing the balance of the autonomic nervous system, whose lability is associated with the concept of the strength of the processes of excitation and inhibition of the central nervous system. Consequently, the individual characteristics of body temperature rhythm can be associated with the temperament of animals and/or their behavior. In this study, we correlated the behavioral features of male greenfinches with the distinctness of ultradian rhythms of body temperature changes in the range of 3–20 min.
Greenfinches were caught in September–October 2019 and 2020 in the village of Yarustovo (Ryazan oblast). The birds were kept until next spring in individual cages indoors. In the winters of 2019–2020 and 2020–2021, we kept 12 and 17 males, respectively. For standardization, the study in the winter 2020–2021 was carried out on birds under the age of one year. The body temperature dynamics was recorded in December: 9 and 14 males were studied in 2019 and 2020, respectively. Body temperature was recorded using DTN4-28 sensors (Embi Reserch), which were implanted into the abdominal cavity for 20–40 days. Zoletil (Virbac Sante Animale) was used as an anesthetic during the surgery; it was injected intramuscularly at a rate of 3–7 mg/kg. The sensor allows recording the body temperature every minute and accumulating data for the entire period of the experiment. The power spectral density of the rhythms was determined by the fast Fourier transform method using the Statistica 7.0 software. Then, we calculated the average spectral density in the ranges of periods of 12–20 and 3–7 min, and found their ratio (hereinafter: the ratio 12–20/3–7 min). This indicator reflects the severity of the amplitude of rhythms with periods of 12–20 min in relation to rhythms with periods of 3–7 min.
To assess the behavioral characteristics of different males, we used two approaches, direct and indirect. As an indirect method, we used the plumage coloration, which in birds can reflect the characteristics of behavior and temperament, e.g., the level of aggressiveness or courage ↔ cowardice [8, 9]. We used the extent (in percent of the total length) of the yellow field of the second rudder from the edge. This feather is two-colored, its distal part is black, and the base is yellow. Previously it was shown that this trait is individual (it stably differs between individuals) and is probably associated with the physiological and/or genetic characteristics of birds [10].
For the direct assessment of behavior, two series of experiments were carried out. In the winter 2019–2020, we studied the aggressive behavior of birds, and in the winter of 2020–2021, reaction to novelty.
To study aggressive behavior, in each of the six cages two males were placed. The “pairs” selection was based on the contrasting length of the yellow part of the rectrix; both birds also belonged to the same age cohort.
After 2–3 weeks, a more aggressive dominant and a subordinate was revealed in each cage. All subordinates of the greenfinch lost most of their plumage of the occipital region (it was plucked out by the dominant bird). The dominant male in all cases was the one with a greater length of the yellow part of the feather. The median ratio of 12–20/3–7 min was higher in dominants: 2.6 (2.3–2.9, n = 4) in dominants versus 2.2 (1.0–2.4, n = 5) in subordinates (the differences were significant according to the Mann–Whitney test: W = 18.5, p = 0.049).
The reaction to novelty was studied in 17 males. The experiments were carried out according to the method described in [8]. At 10 a.m., the birds were deprived of food. After 3.5 h, a new brightly colored feeder was placed in the cage. The time after which the bird began to peck seeds from it (approach latency) was noted. It served as an indicator of the bird’s reaction to novelty. If the male did not approach the feeder for 30 min, the experiment was terminated. The experiment was repeated twice, with an interval of about 10 days for each of the males. In the first series of experiments, the feeder was orange, and in the second, it was bright blue. In experiments with an orange feeder, 12 out of 17 males approached it within 30 min. All 17 birds approached the blue feeder. The approach time to one or another feeder were correlated: Spearman’s correlation coefficient R = 0.65, p = 0.023, n = 12. This means that this experiment reveals individual characteristics of behavior.
The Generalized Linear Mixed Model (GLMM) was used to identify the relationship between the 12–20/3–7 min ratio and the approach latency. In this analysis, a random factor was used, namely, the body weight of the male, measured in the early morning on the day of the first experiment. This was done because the body state of birds can affect food motivation and, subsequently, the experiment. The approach time to the orange feeder did not significantly depend on the ratio 12–20/3–7 min (GLMM: d.f. = 7, t = –1.69, p = 0.135, n = 9); however, the approach latency in case of the blue feeder, depended (GLMM: d.f. = 12, t = –1.81, p = 0.005, n = 14). The nature of the dependence was similar in both experiments (Figs. 2a, 2b). The approach time to the orange (GLMM: d.f.= 10, t = –0.99, p = 0.027, n = 12) and blue (GLMM: d.f.= 15, t = –1.13, p = 0.004, n = 17) feeders significantly depended on the length of the yellow field of the feather (Figs. 2c, 2d).

The approach time for (a, c) the orange feederin the first series of experiments, and (b, d) the blue feeder in the second series of experiments, depending on the ratio 12–20/3–7 min (top) and the proportion of yellow area from the total length of the 4th rectrix (%) (bottom).
Our experiments have shown that birds with a shorter yellow color on the rectrix (1) were subordinate in paired sittings and (2) were less risk-averse (their approach latency to the unfamiliar feeder was longer). According to the data for both years of the study, the length of the yellow area on the tail correlated with the ratio 12–20/3–7 min: Spearman’s correlation coefficient R = 0.69, p < 0.001, n = 23 (Fig. 3). This means that in subordinate and fearful birds (little yellow area) ultradian rhythms of body temperature with a periodicity of 3–7 min were more pronounced (example: Fig. 1b).

Relationship between the ratio 12–20/3–7 min and the proportion of yellow area in the total length of the fourth rectrix (in percent).
Interestingly, the dose of anesthetic (zoletil) required to immobilize birds with a high proportion of yellow area in the tail (70–80%) was 1.5–2 times higher than for birds with a small proportion of yellow area. This additionally confirms the difference in the reactivity of the nervous system in greenfinches with varying degrees of yellow color development on the tail feathers.
The similar data is also known from literature. In barn owl chicks (Tyto alba), the dependence of the size of black plumage spots on the duration and distinctness of ~4-h ultradian rhythms of alternation of activity/rest was revealed [1]. The authors associate this fact with various behaviour strategies inherent in many animals, proactive and reactive [11]. The results of our research show that male greenfinches with a small area of yellow in the tail and pronounced 3- to 7-min fluctuations in temperature can be attributed to reactive individuals, more “cowardly” and less aggressive.
The pattern identified, the predominance of high-frequency fluctuations in the dynamics of minute changes in body temperature in less emotionally stable individuals, is probably determined by endogenous factors associated with the peculiarities of the functioning of the autonomic nervous system. However, we have previously shown that the body temperature dynamics is influenced by an environmental exogenous factor [5]. In particular, a connection was established between fluctuations in body temperature in the range of 4–16 min with Pc5 pulsations of the geomagnetic field [12]. In our study, the birds that were less aggressive and fearful of new objects had more pronounced short-period fluctuations in body temperature and, probably, greater sensitivity to the geomagnetic field, in contrast to individuals showing a tendency to dominate and more confident behavior.
In this study, we have demonstrated the possibility of assessing the state of the autonomic nervous system by the severity of the harmonic spectral density of body temperature changes in different ranges of periods. This technique can also be used in the study of individual psychophysiological reactions, the level of stress load, and other biological parameters, which are reflected in the balance of the sympathetic/parasympathetic activity of the autonomic nervous system.
REFERENCES
Scriba, M.F., Henry, I., Vyssotski, A.L., Mueller, J.C., Rattenborg, N.C., and Roulin, A., J. Biol. Rhythms, 2017, vol. 3, no. 5, pp. 456–468.
Gerkema, M.P., Daan, S., Wilbrink, M., Hop, M.W., and van der Leest, F., J. Biol. Rhythms, 1993, vol. 8, no. 2, pp. 151–171.
Kimura, F. and Tsai, C.W., J. Physiol., 1984, vol. 353, no. 1, pp. 305–315.
Tornatzky, W., Cole, J.C., and Miczek, K.A., Physiol. Behav., 1998, vol. 63, no. 5, pp. 845–853.
Diatroptov, M.E., Panchelyuga, V.A., Panchelyuga, M.S., and Surov A.V., Dokl. Biol. Sci., 2020, vol. 494, pp. 228–231.
Braulke, L.J. and Heldmaier, G., Cryobiology, 2010, vol. 60, no. 2, pp. 198–203.
Baevskii, R.M., Usp. Fiziol. Nauk, 2006, vol. 37, no. 3, pp. 42–57.
Mateos-Gonzalez, F. and Senar, J.C., Animal Behav., 2012, vol. 83, no. 1, pp. 229–232.
Williams, L.J., King, A.J., and Mettke-Hofmann, C., Animal Behav., 2012, vol. 84, no. 1, pp. 159–165.
Karu, U., Saks, L., and Horak, P., Physiol. Biochem. Zool., 2007, vol. 80, no. 6, pp. 663–670.
Koolhaas, J.M., Korte, S.M., De Boer, S.F., Van Der Vegt, B.J., Van Reenen, C.G., Hopster, H., De Jong, I.C., Ruis, M.A.W., and Blokhuis, H.J., Neurosci. Biobehav. Rev., 1999, vol. 23, no. 7, pp. 925–935.
Diatroptov, M.E., Yagova, N.V., Petrovskii, D.V., and Surov, A.V., Byull. Eksp. Biol. Med., 2021, vol. 171, no. 5, pp. 639–644.
Funding
The study was carried out in the framework of the State Contract of the Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, no. 0089-2021-0004 “Ecological and evolutionary aspects of animal behavior and communication.”
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
COMPLIANCE WITH ETHICAL STANDARDS
All procedures performed in studies involving experimental animals When working with experimental were in accordance with the European Convention for the Protection of Vertebrate Animals Used for Experiments or Other Scientific Purposes (Strasbourg, 1986). For the experimental work, a positive conclusion was received from the Ethical Commission of the Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences no. 14 dated January 15, 2018.
CONFLICT OF INTEREST
The authors declare that they have no conflicts of interest.
Additional information
Translated by T. Kuznetsova
Rights and permissions
About this article

Cite this article
Diatroptov, M.E., Opaev, A.S. & Surov, A.V. Ultradian Rhythms of Body Temperature Are Associated with Temperament in Greenfinch (Chloris chloris, Fringillidae, Aves). Dokl Biol Sci 499, 93–96 (2021). https://doi.org/10.1134/S0012496621040025
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0012496621040025