
Abstract
Influence of fish personality on infection rate is poorly studied. In the experiments on young-of-the-year Oncorhynchus mykiss and cercariae of the trematode Diplostomum pseudospathaceum, we tested the hypothesis that infection rate differs between more and less active (“bold” and “shy”) fish. Will individual differences in infection persist upon re-infection? Fish serve as a second intermediate host for this trematode. Positive correlation was found between the results of consecutive infections. Accumulation of parasites with successive infections leads to an aggregated distribution of D. pseudospathaceum among the hosts, affecting individual fitness and polymorphism in fish populations. Persistent individual differences in parasite burden among fish and, as a result, vulnerability for predators confirms the role of parasites as an important factor of natural selection.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
The role of parasites as a factor of formation of stable personality behavioral syndrome (PBS) receives more and more attention [1, 4]. This role may be no less than that of predators. The individual differences in infestation expressed in aggregated distribution of parasites by hosts are well known [10]. Considering that the allocation of dispersal stages in the environment is inhomogeneous, the individual differences in infestation among hosts may result from occasional encounters with aggregations of parasites. However, this may also lead to random distribution of parasites in the host population. For formation of aggregated distribution, when a small part of the population would bear the main parasite burden, some specimens should more frequently encounter parasites, they may be permanently more attractive or less resistant to them. In addition to potential differences in immune protection [5], the aggregated distribution of parasites may be related to PBS of hosts. However, there is almost no information on PBS influence on parasite infestation, except a study on tadpoles of Lithobates sylvaticus. This study revealed that the most active specimens were less infested with larvae of trematodes Echinoparyphium spp. [6].
The stable relationship between traits of individual behavior such as motor activity, exploratory activity, boldness, and aggression are well known in fish [2, 4, 12]. Would more active and “bold” fish (B-fish) sooner start exploration of new situations more infested or less infested with trematode larvae than less active, shy fish (S-fish)? Would individual differences in infestation be retained upon repeated infestation with the same parasites under the conditions of individual infestation in homogeneous environment and infestation in groups in inhomogeneous environment, i.e., under the conditions similar to natural ones? Our experiments were performed on young-of-the-year mykiss Oncorhynchus mykiss infested with cercariae Diplostomum pseudospathaceum liberated by gastropods Lymnaea stagnalis. For these trematodes, the fish were a second intermediate host. The more parasites settle in chrystalline lenses of fish eyes, the easier they can manipulate the host behavior [3], thus increasing the chances to get into the final host, a piscivorous bird. Thus, fitness of the parasite and the host differently depend on the number of metacercariae in fish eyes.
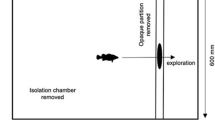
The experiment was made in three stages. At the first stage, the fish were separated into bold and shy according to how rapidly they left the acclimation zone (the start chamber with open exit) and began exploration of the experimental space. B-fish left this zone with 5 min, S-fish used no less than 10–30 min. After that, 40 B-fish and 40 S-fish were used. At the second stage, the fish were infested individually in 2-L tanks at a concentration of cercariae 200 specimens/L. At the third stage, 12 days after the first infestation, the fish were infested repeatedly. In this case, half of the fish (20 B-fish and 20 S-fish) were infested as in the first infestation. The other half was infested in groups composed of two B-fish and two S-fish in a 120-L tank with two sections. The sections were separated by a barrier with a passage for fish at the bottom. The parasites were placed into one of sections to obtain a mean concentration equal to that used in the case of individual infestation. The exposure time was 15 min in the case of individual infestation, 30 min in group infestation. The fish groups were composed in such a way that in each replication the particular specimens of B-fish and S-fish could be discerned by size. On an average in the whole sample the size of B-fish and S-fish did not differ significantly. During the experiment, the fish were fed with pelleted feed and kept in flow-through tanks at a temperature of 15–16оС. Two days after the repeated infestation, the fish were euthanized by means of MS-222, eye lenses were extracted, and the number of parasites in them was counted. Metacercariae from the first and second infestation clearly differed in size and mobility.
Among the B-fish, the infestation rate in the case if individual infestation differed five- to sixfold, and among S-fish, it differed by two- to three-fold. Upon repeated individual infestation, the abundance of parasites both in B-fish and S-fish differed approximately fivefold (Fig. 1). The mean infestation rate of S-fish was by 20% higher than that of B-fish (59 ± 2.8 and 48 ± 3.3, respectively). We assume that the S-fish suffer higher stress and slower get accustomed to a new situation than B-fish do. A higher stress is accompanied by a higher ventilation favoring penetration of parasites through gills (the main way of penetration of these parasites into fish) [9].

Personal infestation (number of metacercaria in fish eyes) with a trematode Diplostomum pseudospathaceum of “bold” and “shy” young-of-the-year“ mykiss Onchorhynchus mykiss in the case of primary and secondary infestation. The primary infestation was made in all fish by separate specimens in homogeneous environment; the secondary infestation was made in half of the fish in the same way as in the primary infestation, another half was infested in groups of four fish (two bold ones, two shy ones) in the heterogeneous environment (parasites were present in one partly isolated section of the tank, not present in another section). (a–c) Solitary fish were infested repeatedly; (d–f) fish in groups were infested repeatedly. Rs—Spearman correlation coefficient.
The discovered positive relationship between the results of consecutive infestations (the Spearmen correlation coefficient for B-fish is Rs = 0.58, p = 0.0006; for S-fish, Rs = 0.84, p < 0.0001) (Fig. 1) indicates stability of personal behavioral/physiological difference making the basis of differential susceptibility to parasites.
In the case of group infestation, the individual differences increase almost twofold (variation coefficient, percent in infestation rate: 77 for B-fish (45% in the case of individual repeated infestation) and 78% for S-fish (35%). On the whole, the positive correlation between the first and second infestation is retained (Fig. 1). This relationship is mainly due to S-fish having the widest range in infestation rate. The highest infestation found in some S-fish hardly may be explained by a higher stress and ventilation as other S-fish fully escaped the repeated infestation. This difference may be caused by a slow reaction to a new situation. Occasionally getting to sites with different concentration of parasites the S-fish probably did not exhibit a rapid reaction to a local situation. The specimens getting to sites with a high concentration of parasites have a higher infestation rate due to both a longer stay in aggregation of cercariae and a higher ventilation rate characteristic of S-fish.
The fish in a group may exchange information on a higher risk of infestation with parasites [8]. This mechanism may probably be more commonly used by B-fish, with their higher motor and exploratory activity. This enables them to discover and avoid sites with a high concentration of parasites thus decreasing the variation of individual infestation. In S-fish, the decreased level of activity may provide a lower infestation risk, at least in a part of the population.
Certain combinations of behavioral characteristics turn out to be most advantageous, i.e., provide the highest adaptability at various conditions [11]. For example, the combination of a high exploratory and motor activity with boldness supplies undisputable advantages in resource search in the heterogeneous environment [14]. The same qualities may make an animal more vulnerable for predators and parasites when these natural enemies are not evenly distributed in space.
Bold and shy Lepomis gibbosus captured in nature differed in the composition of infracommunities of parasites which were related to their different exploratory activity [13]. It is difficult to reveal the role of behavioral or physiological mechanisms influencing the infestation rate of fish and supporting stable individual differences on such material. Our experimental data demonstrate that differences in infestation rate are retained at consecutive “waves” of infestation not only in maximally simple (localized) situation in homogeneous environment in the absence of interactions between fish. The individual differences in infestation rate are reproducible also in a more realistic environment where the fish may interact and move between biotopes. Being accumulated in consecutive infestations these effects may be an important factor of formation of the aggregated distribution of parasite among hosts, influence the individual adaptability and support polymorphism in populations.
In the parasite–host systems where the definite host is a predator (in the present case, a piscivorous bird) the parasite may play a role of occasional “noise” or “catalyzer” intensifying the influence of a predator. The parasites themselves may play a role of a selection factor ensuring stable personal differences in the fish with different “personality.” A higher infestation rate decrease survival of hosts, first of all due to greater vulnerability for predators.
REFERENCES
Barber, I. and Dingemanse, N.J., Philos. Tr. Roy. Soc. B, 2010, vol. 365, pp. 4077–4088.
Budaev, S.V., Mikheev, V.N., and Pavlov, D.S., Biol. Bull. Rev., 2015, vol. 76, no. 1, pp. 26–47.
Gopko, M.V., Mikheev, V.N., and Taskinen, J., Behav. Ecol. Sociobiol., 2015, vol. 69, 1723–1730.
Kekäläinen, J., Lai, Y.T., Vainikka, A., Sirkka, I., and Kortet, R., Behav. Ecol. Sociobiol., 2014, vol. 68, pp. 197–204.
Koolhaas, J.M., Brain Behav. Immun., 2008, vol. 22, pp. 662–667.
Koprivnikar, J., Gibson, C.H., and Redfern, J.C., Proc. R. Soc. Lond. B, 2012, vol. 279, pp. 1544–1550.
Kortet, R., Hedrick, A.V., and Vainikka, A., Ecol. Lett., 2010, vol. 13, pp. 1449–1458.
Mikheev, V.N., Pasternak, A.F., Taskinen, J., and Valtonen, E.T., Paras. Vect., 2013, vol. 6, p. 301.
Mikheev, V.N., Pasternak, A.F., Taskinen, J., and Valtonen, E.T., Paras. Vect., 2014, vol. 7, p. 281.
Shaw, D.J., Grenfell, B.T., and Dobson, A.P., Parasitology, 1998, vol. 117, pp. 597–610.
Sih, A., Bell, A.M., and Johnson, J.C., Trends Ecol. Evol., 2004, vol. 19, pp. 372–378.
Sneddon, L.U., J. Fish Biol., 2003, vol. 62, pp. 250–260.
Wilson, D.S., Coleman, K., Clark, A.B., and Biederman, L.J., Comp.Psychol., 1993, vol. 107.
Wolf, M., van Doorn, G.S., Leimar, O., and Weissing, F.J., Nature, 2007, vol. 447, pp. 581–585.
Funding
The experimental investigations were supported by the Russian Foundation of Basic Research (project no. 17-04-00247a), processing of the material and composition of MS were supported by the Russian Science Foundation (project no. 19-14-00015) and were supported with scope of the State Contract no. 0149-2019-0008.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. All applicable international national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by N. Smirnov
Rights and permissions
About this article

Cite this article
Mikheev, V.N., Pasternak, A.F. & Taskinen, J. Personality Influences Risk of Parasitism in Fish. Dokl Biol Sci 488, 141–144 (2019). https://doi.org/10.1134/S0012496619050053
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0012496619050053