
Abstract
Strophostyles helvola is a close relative to common bean (Phaseolus vulgaris) and inhabits both coastal and non-coastal regions in North America. However, the mechanism of saline adaptation in S. helvola remains unclear. A transcriptome profiling would facilitate dissecting the underlying molecular mechanisms in salinity-adapted S. helvola. In this study, we reported the RNA-seq analyses of two genotypes (a salt-tolerant beach genotype and a salt-sensitive inland genotype) of S. helvola stressed with salt. S. helvola plants were grown in pots and treated with half lethal-guided dose of NaCl solution for 3 h, 24 h, and 7d. The plants supplied with the same amount of water were used as controls. The whole roots sampled from the three time points were equally pooled as one biological replicate, and three replicates were used for library construction and transcriptome sequencing on Illumina Hiseq 2500. The comparative analyses of root transcriptomes presented here provides a valuable resource for discovery of genes and networks involved in salt tolerance in S. helvola.
Design Type(s) | stimulus or stress design • strain comparison design • transcription profiling design |
Measurement Type(s) | transcription profiling assay |
Technology Type(s) | RNA sequencing |
Factor Type(s) | genotype • experimental condition |
Sample Characteristic(s) | Strophostyles helvola • root |
Machine-accessible metadata file describing the reported data (ISA-Tab format)
Similar content being viewed by others


Background & Summary
Soil salinity is becoming a critical environmental factor constraining plant growth and crop productivity worldwide because a majority of crops cultivated in irrigated arable land are salt sensitive. In addition, an increase of crop production is in high demand to sustain the growing human population, thus imposing a need of crop cultivation in marginal or coastal land. A sustainable and environmental-friendly alternative strategy is to develop salt-tolerant crops that can thrive in saline soils.
Next-generation sequencing of transcriptomes has been widely used to characterize the global expression patterns in various organisms under diverse conditions to facilitate gene discovery and address major questions associated with plant environmental stress responses1. In plants, global transcriptome profiling has been performed to elucidate molecular mechanisms of salt tolerance or response, especially in salinity-adapted genotypes mainly in model systems. Recently, more and more reports on non-model species with limited genomic sequence knowledge have been published. For example, the transcriptome analyses on Ipomoea imperati, a wild relative of sweet potato tolerant to high salinity, revealed the ABA signalling pathway and two membrane transporter genes2. Sonneratia alba represents one of the most salt tolerant mangrove species, and RNA-seq on it has identified salt responsive genes with signatures of natural selection3. These transcriptomic studies have allowed us to uncover gene expression mechanisms and novel genes beyond what we currently know from model species.
Strophostyles helvola (L.) Elliott, belonging to Fabaceae family, is herbaceous annual vine native to North America. S. helvola can colonize in wild places, either moist or dry conditions with preference in sandy soils, thus it is also called wild bean or sand bean. In addition, sand bean is a close relative of common bean (Phaseolus vulgaris L.), one of the most important grain legumes worldwide providing protein for human consumption and having roles in fixing atmospheric nitrogen4. Like other leguminous crops, cultivated common bean is a glycophyte, sensitive to salt, leading to reduced production if grown under salt conditions5. Thus far, molecular mechanisms of common bean’s tolerance to salinity was rarely studied6,7, thus the underlying mechanisms remain largely unclear. As a close halophyte relative of common bean, a global investigation of molecular response of salinity-adaptive sand bean can be very helpful with revealing the underlying mechanisms toward the goal to develop salt tolerant common bean.
In this study, we presented the transcriptome analyses of two genotypes of S. helvola (halophyte vs glycophyte) stressed with over half-lethal doses of NaCl which was not previously reported. We described the detailed procedure of transcriptome profiling for NaCl-treated and non-treated root tissues of a salt tolerant Beach genotype and a salt sensitive Inland genotype, respectively, during the time-course treatment of 3 h, 24 h, and 7d. In total, 30.5 Gb of transcriptome data (fastq.gz) from twelve root tissues were generated. We also presented in detail the analytic methods of how to obtain the raw sequence, quality control, sequence alignment, and differential expression analyses. We conducted data pre-processing to indicate the high quality of our data as visualized with FastQC and robustness of our results using analyses of multidimensional scales (MDS), dendrogram clustering and expression pattern analyses. The comparative expression profiling of the two S. helvola genotypes will provide valuable resources of genomic data for studying salinity tolerance in S. helvola and others leguminous relatives.
Methods
Overview of experimental design
The whole root tissues that were treated with NaCl and mock (water) were dissected from the plants during the time course investigation at 3 h, 24 h, and 7d. The frozen tissues collected from the three time points were pooled to generate a biological replicate. Total RNA was extracted from the pooled tissues using Qiagen RNeasy mini kit (Qiagen, Valencia, CA, USA) according to the manufacture’s instruction followed by library construction and transcriptome sequencing. The quality-controlled reads were aligned to P. vulgaris reference genome, P. vulgaris_218_v1.0 (https://phytozome.jgi.doe.gov). The resulting .bam files were used for differential expression analyses using edgeR8. The experiment design and analysis pipeline were shown in Fig. 1.

A time-course NaCl treatment was used to capture the maximum change in transcriptome of S. helvola plants. The transcriptome changes were obtained by a comparison of responses between treated plants with their counterpart controls. Briefly, 10-d old plants were supplied with NaCl solution for treatment, in parallel, the plants supplied with water were used as controls. Three biological replicates per condition were used for transcriptome sequencing. All raw reads were quality controlled prior to aligning to P. vulgaris reference genome (v1.0). The uniquely aligned reads were used for expression profile analyses.
Materials and treatment
Two S. helvola genotypes were used here for comparative study. One genotype was originally from the sandy soil along the beach in east coast of North Carolina, we named it “Beach genotype” in our study; The other genotype was originally from central Missouri, and we named it “Inland genotype”.
To determine the dose of NaCl that may trigger the tolerance response to salinity, we first tested the median lethal dose (LD50, the salt concentration required to kill 50% of the plants). The beach and inland genotypes were watered every 2 days with increasing increments of 50 mM NaCl solution9. We found that two genotypes required different doses of NaCl to cause 50% plants die. For Inland genotype, a final dose 350-mM NaCl may cause lethal toxicity in half, while a higher dose, 600 mM of NaCl, was needed for Beach genotype. These lethal limit data were used to guide the salt treatment for the following experiment.
The seeds were germinated in a growth chamber (Percival Scientific Inc., Perry, IA, USA) and the healthy seedlings were transplanted into flats filled with soil. Briefly, seed coat was sliced to facilitate germination and placed on moist filter paper on a petri dish. 3-day post germination, the seedlings were transplanted in 3 × 6 flat (3.10′ × 3.10′ × 2.33′) (Greenhouse MegaStore, US) filled with potting mix soil. For salt treatment for RNA-seq assay, we use the dose that was slightly lower than the dose identified for the lethal limit mentioned above, thus the plants could be stressed enough but would not die. Thus, we used 200 Mm and 400 Mm of NaCl solution as a final concentration to stress Inland and Beach genotypes, respectively. For treatment, the plants were supplied with 50 mL NaCl solution per day. In parallel, plants supplied with the same amount of water were used as controls. Whole roots were sliced off from the treatment and control plants at 3 h, 24 h and 7d, respectively, after the concentration of NaCl was reached as designed and flash frozen in liquid nitrogen. All frozen tissues were stored at −80 °C for RNA extraction. Three biological replicates per collection were collected.
RNA extraction, Library construction, and RNA sequencing
For each biological replicate, we used pooled root tissue from three plants for the library construction and sequencing, with each collected at 3 h, 24 h, and 7d, respectively. Briefly, the individual roots were ground and equal amounts of ground roots from the three time points were pooled to generate one biological replicate. Thus, three biological replicates per condition were generated. In total, twelve samples for both genotypes were used for total RNA extraction. Total RNA was isolated with RNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA). Purified RNA was quantified using a Quant-iT™ RiboGreen™ RNA Assay Kit (Invitrogen Carlsbad, CA, USA) and its integrity was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). One microgram of RNA samples with RNA integrity number (RIN) ≥ 7.0 (Table 1) from three independent biological replicates of each condition was used to generate cDNA libraries with insert sizes ranging from 300 to 350 bp using a TruSeq RNA Library Prep Kit from Illumina. Libraries were combined into a single pool and a 125 bp single-read sequencing run was conducted using a HiSeq 2500 instrument (Illumina, San Diego, CA, USA). Primary processing was performed on the raw reads to generate FASTQ files. RNA extraction, library construction and sequencing were performed in the Genomics Laboratory in the David H. Murdock Research Institute (Kannapolis, NC, USA).
Pre-processing of sequencing data
The quality of the raw sequence generated from transcriptome sequencing was assessed with FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Low quality (< 20) bases and adapter sequences were trimmed with Trimmomatic v 0.3610 with following parameters: ILLUMINACLIP: path/to/adaptor.fa:2:30:10 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:36. After filtering, the remaining reads were called “clean reads” and were re-assessed with FastQC. All the results of FastQC were merged and visualized using MultiQC (http://multiqc.info). Clean reads were aligned to common bean (Phaseolus vulgaris) reference genome Pvulgaris_218_v1.0 (https://phytozome.jgi.doe.gov) using RNA-seq aligner STAR software11. The general feature format (gff3) file (Pvulgaris_218_v1.0.gene_exons.gff3) corresponding to Pvulgaris_218_v1.0 downloaded at Phytozome was used as an input for STAR. The options used for running STAR are: --runThreadN 16 --genomeDir/path/to/directory --sjdbGTFtagExonParentTranscript Pvulgaris_218_v1.0.gene_exons.gff3 --readFilesIn read.fastq.gz --readFilesCommand zcat --outFileNamePrefix Name --outSAMtype BAM Unsorted SortedByCoordinate. The STAR-resultant .bam files were used to estimate the abundance of uniquely-mapped reads using FeatureCounts12. Difference expression analyses was conducted using EdgeR13. Heat maps were made using heatmap.2 function of the gplots package14.
Code availability
Codes that were used for data processing are included in the Methods and available as supplementary material (Supplementary File 1).
Data Records
The project was deposited into the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) accession (Table 1 and Data Citation 1). The abundance count for all the samples was deposited at Gene Expression Omnibus (GEO) database (Data Citation 2).
Technical Validation
Quality control
A total of 12 RNA libraries were prepared and sequenced with the sequencing depth ranging from 41.0–55.7 million single-end reads (Table 1). We applied FastQC to determine the data quality and measured several important parameters. The assessment for the filtered data was shown in Fig. 2, and the distribution of mean quality score and per sequence quality scores indicated the high quality of filtered sequences, with scores of most sequences over 35. Over 99% of the raw reads were kept after quality control and a 75.89–79.36% of the clean reads were mapped to unique location in the common bean reference genome Pvulgaris_218_v1.0 (Table 1).

(a) Mean quality scores per position. (b) Per sequence quality scores. (c) GC content distribution. (d) Read length distribution.
Analysis of RNA-seq data
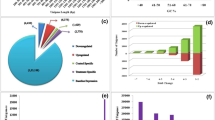
The clean reads with single alignment on P. vulgaris reference exosome were counted (Fig. 3a) and normalized by counts per million (Fig. 3b) for differential expression analysis. Gene clustering analyses were used to examine the difference between the biological replicates. Figure 3c shows that three replicates from the same group cluster together while samples from different groups are well separated. This result was further supported by multi-dimensional scale (Fig. 3d) showing that gene expression profiles of all biological replicates can clearly separate the four groups and cluster biological replicates together with small variability per group. We further explored the expression profiles in two comparisons and visualized with mean-difference (MD) plots. As shown in Fig. 3e and f, a majority of the genes are cantered around the line of zero log(fold change), and the differentially expressed genes (DEGs) were highlighted in red with a threshold of fold change ≥ 2 and fdr ≤ 0.05. We identified 2910 DEGs in Beach genotype and have showed their expression pattern across all the samples in Fig. 3g. Consistent with Fig. 3c, replicates from each group are clustered together. DEGs showing distinct expression pattern in treated Beach genotype compared with the other three groups merit further exploration.

(a) Library size of each replicate. (b) Distribution of log2 transformed count per million. (c) Clustering analyses of gene expression in all 12 samples. (d) Multidimentional scale analyses of gene expression in all 12 samples. All replicates per condition cluster together. (e) And (f) showed the MD plots of log2-expression and average abundance of each gene. Each dot represents a gene. Significantly up and down regulated genes are highlighted in red above log2(fold change)= 1 and below log2(fold change) = −1, respectively. (g) A heat map showing expression patterns of 2910 differential expressed genes across 12 samples.
Usage Notes
The RNA-seq fastq.gz files were deposited at NCBI SRA public repository and could be downloaded using fastq-dump tool of SRA Toolkit (https://www.ncbi.nlm.nih.gov). Other than Trimmomatic, FASTX (http://hannonlab.cshl.edu/fastx_toolkit/) and cutadapt15 are also commonly used for trimming and adapter removal. The alternative aligners for RNA-seq sequence could also be used, such as TopHat216 and HISAT217. The reference genome of P. vulgaris, the annotation file, and gff3 file could be retrieved at Phytozome database (https://phytozome.jgi.doe.gov). For downstream differential expression analyses, Cufflinks package coupled with CummeRbund may generate transcriptome assembly, expression abundance, differential expression analyses, and visualization of analyses results. HTSeq18 could also be used as alternative of featureCounts for quantification and performed differential expression analyses with DESeq219.
Additional information
How to cite this article: Zhang, H. et al. Transcriptome profiling of a beach-adapted wild legume for dissecting novel mechanisms of salinity tolerance. Sci. Data. 5:180290 doi:10.1038/sdata.2018.290 (2018).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
References
Han, Y. X., Gao, S. G., Muegge, K., Zhang, W. & Zhou, B. Advanced applications of RNA sequencing and challenges. Bioinform Biol Insig 9, 29–46 (2015).
Luo, Y. et al. Salt tolerance response revealed by RNA-Seq in a diploid halophytic wild relative of sweet potato. Sci Rep 7, 9624 (2017).
Chen, S. F. et al. Transcriptome sequencing of a highly salt tolerant mangrove species Sonneratia alba using Illumina platform. Mar Genom 4, 129–136 (2011).
Schmutz, J. et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat Genet 46, 707–713 (2014).
Maas, E. V. & Hoffman, G. J. Crop salt tolerance-current assessment. J Irrig Drainage Div 103, 115–134 (1977).
Hiz, M. C., Canher, B., Niron, H. & Turet, M. Transcriptome analysis of salt tolerant common bean (Phaseolus vulgaris L.) under saline conditions. PLoS One 9, e92598 (2014).
Buyuk, I. et al. Genome-wide identification of salinity responsive HSP70s in common bean. Mol Biol Rep 43, 1251–1266 (2016).
McCarthy, D. J., Chen, Y. S. & Smyth, G. K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res 40, 4288–4297 (2012).
Orsini, F. et al. A comparative study of salt tolerance parameters in 11 wild relatives of Arabidopsis thaliana. J Exp Bot 61, 3787–3798 (2010).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Dobin, A . et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Warnes, G. R. et al. gplots: various R programming tools for plotting data. https://cran.r-project.org/web/packages/gplots/index.html (2016).
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnetjournal 17, 10–12 (2011).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol 14, R36 (2013).
Kim, D., Landmead, B. & Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat Methods 12, 357–360 (2015).
Anders, S., Pyl, P. T. & Huber, W. HTSeq-a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15, 550 (2014).
Data Citations
Acknowledgements
B.S. was supported by The National Institute of General Medical Sciences of the National Institutes of Health (Award Number: R15GM122029) and University of North Carolina (2018_FRG_SONG).
Author information
Authors and Affiliations
Contributions
B.S. conceived the project and designed the experiment. C.Z. conducted the experiment. H.Z., D.P., J.K., S.F. performed the data analyses. H.Z. and B.S. wrote the manuscript. All authors read the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
ISA-Tab metadata
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/ The Creative Commons Public Domain Dedication waiver http://creativecommons.org/publicdomain/zero/1.0/ applies to the metadata files made available in this article.
About this article

Cite this article
Zhang, H., Zuelsdorf, C., Penneys, D. et al. Transcriptome profiling of a beach-adapted wild legume for dissecting novel mechanisms of salinity tolerance. Sci Data 5, 180290 (2018). https://doi.org/10.1038/sdata.2018.290
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/sdata.2018.290
- Springer Nature Limited
This article is cited by
-
Integrated de novo transcriptome of Culex pipiens mosquito larvae as a resource for genetic control strategies
Scientific Data (2024)
-
Water requirements and crop coefficients of edible, spicy and medicinal herbs and vegetables; a review aimed at supporting plant and water management
Irrigation Science (2024)
-
Transcriptomic analysis reveals candidate genes associated with salinity stress tolerance during the early vegetative stage in fababean genotype, Hassawi-2
Scientific Reports (2023)