
Abstract
The greater amberjack Seriola dumerili is a promising candidate for aquaculture production. This study compares the ovary transcriptome of greater amberjack sampled in the wild (WILD) with hatchery-produced breeders reared in aquaculture sea cages in the Mediterranean Sea. Among the seven sampled cultured fish, three were classified as reproductively dysfunctional (DysF group), while four showed no signs of reproductive alteration (NormalF group). The DysF fish showed 1,166 differentially expressed genes (DEGs) compared to WILD females, and 755 DEGs compared to the NormalF. According to gene ontology (GO) analysis, DysF females exhibited enrichment of genes belonging to the biological categories classified as Secreted, ECM-receptor interaction, and Focal adhesion. Protein–protein interaction analysis revealed proteins involved in the biological categories of ECM-receptor interaction, Enzyme-linked receptor protein signaling, Wnt signal transduction pathways, and Ovulation cycle. KEGG pathway analysis showed DEGs involved in 111 pathways, including Neuroactive ligand-receptor interaction, Steroid hormone biosynthesis, Cell cycle, Oocyte meiosis, Necroptosis, Ferroptosis, Apoptosis, Autophagy, Progesterone-mediated oocyte maturation, Endocytosis and Phagosome, as well as Hedgehog, Apelin, PPAR, Notch, and GnRH signalling pathways. Additionally, DysF females exhibited factors encoded by upregulated genes associated with hypogonadism and polycystic ovary syndrome in mammals. This study -which is part of a broader research effort examining the transcriptome of the entire reproductive axis in greater amberjack of both sexes-, enhances our comprehension of the mechanisms underlying the appearance of reproductive dysfunctions when fish are reared under aquaculture conditions.
Similar content being viewed by others


Introduction
Rearing in captivity is often associated with reproductive dysfunctions of variable degrees in fish1,2. In males, reproductive dysfunctions may impact spermatogenesis3,4, ultimately leading to the production of sperm of low volume and quality3,4,5,6. In females, commonly observed reproductive dysfunctions include (a) the incapacity of oocytes to enter vitellogenesis (process of oocyte uptake of vitellogenin and accumulation of its derived yolk proteins in the form of yolk granules), (b) the arrest of vitellogenesis before completion, resulting in the loss of vitellogenic follicles via atresia, and (c) the failure of fully vitellogenic oocytes to undergo maturation, consequently leading to the lack of ovulation and spawning1,2,7.
Confinement-induced stress3,8,9,10, lack of suitable environmental conditions for reproduction11,12,13 and nutritional deficiencies14,15 are considered as the main causes of reproductive dysfunctions in aquaculture. The functional alterations of the brain-pituitary-gonad axis associated with these reproductive dysfunctions have not yet been fully elucidated. Low plasma levels of luteinizing hormone (Lh) concomitantly with high Lh pituitary content have been observed in fish failing to undergo oocyte maturation, ovulation and spawning, suggesting a reduced pituitary secretion of this hormone at the completion of vitellogenesis2,7,16,17,18. In fact, the stimulation of Lh release through the administration of gonadotropin releasing hormone agonists (GnRHa) is widely used as a therapy to alleviate this reproductive dysfunction in several fish species1,2,5,7,18,19,20,21,22,23,24.
The greater amberjack Seriola dumerili has a circumglobal distribution in the temperate zones of the three oceans and adjacent seas, including the Mediterranean Sea25,26,27. A worldwide consumers’ appreciation, rapid growth and high market prices28,29 make the greater amberjack an excellent candidate for domestication and aquaculture production throughout the world, along with other congeneric species30.
It has been well documented that greater amberjack caught from the wild in the Mediterranean Sea and reared under routine farming conditions (thereafter referred to as “wild-caught” fish) undergo severe reproductive dysfunctions. These same dysfunctions have been documented also in hatchery-produced (F1 generation) greater amberjack in aquaculture facilities in the Mediterranean Sea. In wild-caught fish reared in outdoor tanks, the occurrence of atretic degeneration of vitellogenic follicles was the first documented dysfunction31,32. Recently, a few studies on the oogenesis of wild-caught individuals confirmed the limited capacity of this species to spawn spontaneously15,33. In fact, wild-caught females had low ovary relative mass (gonado-somatic index, GSI), more than 50% of atresia of vitellogenic follicles during the phase of active gametogenesis, and up to 100% atretic vitellogenic follicles during the spawning season15,33. Wild-caught males, on the other hand, had low GSI, small seminiferous tubules, early cessation of the active spermatogenesis phase, and a high rate of germ cell apoptosis4. The gametogenesis impairment in both sexes was associated with low plasma concentrations of sex steroids in males4 and females15,33,34. Spermatogenesis impairment was also documented in hatchery-produced cultured greater amberjack. It was characterized by a precocious arrest of spermatogenesis, associated with the dysregulation of a large number of genes involved in various interconnected biological processes, such as steroidogenesis, cell cycle, meiosis, cell assembly, and apoptosis35,36.
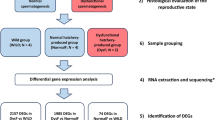
This study reports a comparative analysis of the ovary transcriptomes of wild greater amberjack reproducing in their natural reproductive grounds around the Pelagie Islands (Sicily, Italy) and hatchery-produced females reared in aquaculture sea cages, both sampled during the natural reproductive season in the Mediterranean Sea. This work is part of a broader research effort aimed at describing the effects of rearing in captivity on gene expression along the reproductive axis of greater amberjack, in order to elucidate the causes of reproductive dysfunctions in aquaculture.
Methods
Ethics
For this study, wild fish sampled in nature and hatchery-produced, farmed greater amberjack females were used. Wild fish beyond the size of first maturity37 were caught by an authorized commercial purse-seine fishing vessel during routine fishing operations. Farmed fish were produced at Argosaronikos Fishfarming S.A. (Salamina Island, Greece) under routine farming conditions. The use of farmed fish in this study was approved by the Greek National Veterinary Services (authorization number: AP 31337). All procedures involving animals were conducted in accordance with the “Guidelines for the treatment of animals in behavioural research and teaching”38, the “Ethical justification for the use and treatment of fishes in research: an update”39, and the “Directive 2010/63/EU of the European parliament and the council of 22 September 2010 on the protection of animals used for scientific purposes”40. The authors complied with the ARRIVE guidelines.
Sampling
Three wild and seven farmed greater amberjack females were sampled between 18th May and 1st June 2021 during the active gametogenesis period of the wild population in the Mediterranean Sea15. Wild individuals were caught around the Pelagie Islands (Sicily, Italy) from a purse-seine fishing vessel and sampled immediately after death. Farmed, hatchery-produced (first generation, F1) fish were produced from eggs obtained in Argosaronikos Fishfarming S.A. in 2017, following spawning induction of wild-caught breeders23,24 and were reared in sea cages under common aquaculture practices. When the fish reached reproductive maturity (2021), they were fed a commercial broodstock diet (Skretting, Vitalis Prima) 3 to 5 times a week until apparent satiation.
Before sampling, hatchery-produced fish were confined in a small PVC cage connected to the rearing cage. They were tranquilized with about 0.01 ml l−1 of clove oil (Roumpoulakis E.P.E., Greece) dissolved in ethanol at a 1:10 ratio. Subsequently, they were gently directed into a PVC stretcher and brought aboard a service vessel, where they were deeply anesthetized with 0.03 ml l−1 of clove oil in a 1-m3 tank. The fish were then euthanized by decapitation, placed in crushed ice and transferred to the farm facility for biometric data collection and tissue sampling. The time interval between fish death and sampling ranged between 30 min and 2 h.
For each fish, biometric data (fork length, FL, nearest cm; body mass, BM, nearest hg; gonad mass, GM, nearest g) were recorded, and the gonadosomatic index was calculated as GSI = 100 GM BM−1 (Table 1). Ovaries were excised and preserved as below specified.
Histological analysis of greater amberjack ovaries
For the histological analysis of ovaries, 1-cm thick gonad slices were cut and fixed in Bouin's solution, dehydrated in ethanol, clarified in xylene and embedded in paraffin wax. Five-μm thick sections were then stained with hematoxylin–eosin. The assessment of reproductive state was performed on the basis of the most advanced oocyte stage and the occurrence of post-ovulatory (POFs) and atretic vitellogenic follicles15,19. According to41, atretic vitellogenic follicles were identified on the basis of the fragmentation of the zona radiata, invasion of hypertrophic granulosa cells into the oocyte and progressive yolk reabsorption.
RNA extraction and sequencing
For RNA-seq, small ovary samples were stored in RNA later® (Thermo Fisher Scientific, Waltham, Massachusetts, U.S.), transported to the laboratory within one week, and frozen at -80 °C. Total RNA extraction was performed on 2.5 mg ovary samples, lysed and homogenised with TissueLyser II (Qiagen, Germany) setting 2’ and 20 Hz frequency, by RNeasy® Plus Micro kit (Qiagen, Germany) following the manufacturer's protocol. The quantity and quality of extracted total RNA were checked by Nanodrop 1000 spectrophotometer (Thermo Scientific, Waltham, Massachusetts, U.S.) and Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, California, U.S.), respectively. High-quality RNA samples (RIN range 7–8) were then used to prepare the mRNA libraries by SureSelect Strand Specific RNA Library Preparation kit (Agilent Technologies, Santa Clara, California, U.S.). In particular, poly-A selection and directional mRNA libraries were carried out using 1 µg of total RNA. Finally, paired-end sequencing (2 × 75 bases) was performed on the Illumina NextSeq platform (Illumina Inc., San Diego, California, U.S.).
RNAseq data analysis
Raw sequencing data in FASTQ format were quality-checked using the FastQC program (http://www.bioinformatics.babraham.ac.uk/projects/fastqc) and adaptor sequences as well as low quality regions (phred cutoff < 25) were trimmed using fastp (version 0.20.0) (with parameters: -detect_adapter_for_pe -x -q 25 -n 1 -l 50 -y -w 8)42. Cleaned reads were aligned onto the Seriola dumerili reference genome (Sdu_1.0, assembly accession GCF_002260705, https://www.ncbi.nlm.nih.gov/assembly/GCF_002260705.1) using STAR (version 020,201)43 with default parameters. Read counts per gene were performed by featureCounts (version 1.6.0)44 and differential gene expression analysis was carried out using DESeq245. Only genes with an adjusted P value ≤ 0.05, |log2(FC)|> 1.5 and |log2(FC)|< −1.5 were used for downstream analyses.
DAVID (Database for Annotation, Visualization, and Integrated Discovery database https://david.ncifcrf.gov/tools.jsp)46 and ShinyGO (http://bioinformatics.sdstate.edu/go)47 were used to perform the functional annotation of differently expressed genes (DEGs), and the gene ontology (GO) enrichment analysis. By applying a false discovery rate (FDR) < 0.05, these analyses were able to identify specific categories (biological processes, molecular functions, cellular components and pathways), potentially involved in reproductive dysfunctions. A protein–protein interaction (PPI) network based on DEGs associated with each comparison was built using STRING (https://string-db.org/). DEGs were mapped to Kyoto encyclopedia of genes and genomes (KEGG) pathways using KEGG Mapper—Search (https://www.genome.jp/kegg/mapper/search.html)48.
All queries launched on DAVID, ShinyGO and STRING were restricted to taxon ID 41447 (Seriola dumerili).
Results
Evaluation of reproductive state
The three wild fish had mid-to-late vitellogenesis as the most developed oocyte stage (maximum oocyte diameter ≈ 450 μm) (Table 1; Fig. 1a). This stage of oogenesis was the expected stage at the time of sampling and served as the normal reference group for the RNAseq comparative study (WILD group). Among the seven hatchery-produced greater amberjack females, four had mid-to-late vitellogenic oocytes (maximum oocyte diameter ≈ 410 μm) as the most advanced oocytes stage and displayed mild (< 50%) atresia of vitellogenic follicles (Table 1; Fig. 1b). These fish were considered as fish undergoing normal oogenesis and were designated as “non-dysfunctional farmed” fish (NormalF) for ovary RNAseq analysis. The other three individuals were affected by severe (> 50%) atresia of the most advanced vitellogenic follicles and were designated as “dysfunctional farmed fish” (DysF) for ovarian RNAseq analysis. The atretic vitellogenic follicles were in the alpha or beta stage of atresia. Alpha atretic follicles were characterized by large vitellogenic follicles showing a breakdown of the nuclear envelope, zona radiata disappearance, coalescence of yolk granules (Fig. 1c). Atretic follicles in the beta stage were smaller than alpha atretic follicles and were devoid of yolk granules (Fig. 1a, c).

Micrographs of ovary sections from wild and hatchery-produced greater amberjack. (a,b) Ovaries from wild and hatchery-produced fish, respectively, showing mid-late vitellogenesis as the most advanced oocyte stage and minor atresia of vitellogenic follicles. (c) Ovary from a hatchery-produced fish showing major atresia of the most advanced vitellogenic follicles (100% atresia). H-E staining. Magnification bars = 150 μm in (a,b), 200 μm in (c). pg primary growth ovarian follicles, α alpha atretic follicle, β beta atretic follicle, vf vitellogenic follicle.
RNAseq and differential gene expression analysis
The ovary comparative transcriptome analysis among the three groups of fish in different reproductive conditions (WILD, NormalF and DysF) produced an average of 28 million paired-end reads per sample. After an appropriate cleaning procedure, high-quality reads were aligned to the Seriola dumerili reference genome. About 90% of the cleaned reads were uniquely mapped to the reference genome. Reads generated in this study are freely available through the SRA (Short Read Archive) database under the BioProject accession number PRJNA1056146.
The transcriptome analysis identified 21,678 genes, of which 19,621 were shared among the three groups, and 351, 359, and 176 genes were specifically expressed in the WILD, NormalF, and DysF groups, respectively (Fig. 2a, Supplementary Table S1). The principal component analysis (PCA) of the 1000 most variable genes showed a clear separation of samples belonging to the DysF group, from the WILD and NormalF groups (Fig. 2b).

Venn diagram (a) and PCA (b) of shared and unique genes related to ovary samples of wild (WILD), dysfunctional (DysF) and normal (NormalF) hatchery-produced greater amberjack.
Differential gene expression analysis showed 1166 dysregulated genes in the comparison DysF vs. WILD, of which 1072 were downregulated and 94 upregulated; 755 dysregulated genes in the comparison DysF vs. NormalF, of which 687 were downregulated and 68 upregulated; and only 20 dysregulated genes in the comparison NormalF vs. WILD, of which 11 were downregulated and nine upregulated (Supplementary Table S2). Among the observed DEGs, five were shared between the DysF vs. WILD and NormalF vs. WILD comparisons, seven were shared between the DysF vs. NormalF and NormalF vs. WILD comparisons. Additionally, 621 genes were shared between the DysF vs. NormalF and DysF vs. WILD comparisons (Fig. 3; Supplementary Table S3).

Venn diagram of shared and unique DEGs in the DysF vs. NormalF, DysF vs. WILD and NormalF vs. WILD comparisons. DysF dysfunctional farmed group, NormalF non-dysfunctional farmed group, WILD wild group.
GO enrichment analysis
Biological categories related to gene ontology (GO) enrichment analysis performed on DEGs of each comparison were showed in Table 2. DysF fish exhibited statistically significant enrichment of genes, compared with non-dysfunctional groups (NormalF and/or WILD) in various biological categories (Supplementary Table S4). Specifically, the DysF group showed enriched genes associated with cellular components, including the GO terms Secreted (i.e., vascular endothelial growth factors; insulin like growth factor binding protein 4; inhibin subunit alpha; neuron-derived neurotrophic factor) and Extracellular matrix (i.e., nidogen-1-like; tenascin-1-like; collagen type VI alpha 3 chain; adamts genes). Additionally, DysF females showed significantly enriched genes with molecular functions related to the GO terms Receptors (i.e., AXL receptor tyrosine kinase; eph receptor A3; androgen receptor-like; low density lipoprotein receptor-related protein 5; platelet-derived growth factor receptor; sphingosine-1-phosphate receptor 2; smoothened, frizzled class receptor). Furthermore, KEGG pathways significantly enriched by DEGs in the DysF group included ECM-receptor interaction (i.e., integrin, alpha 1; collagen type VI alpha 3 chain; dystroglycan 1; laminin, alpha 4), Focal adhesion (i.e., integrin, alpha 1; collagen, type IV, alpha 5; platelet derived growth factor d) and Regulation of actin cytoskeleton (i.e., integrin alpha 1; fibroblast growth factor 5; fibroblast growth factor receptor 2, moesin-like).
Although no enriched biological category emerged, a few dysregulated genes involved in inflammation (ptx3), angiogenesis (xdh, ptx3), neurogenesis/dendritic spine morphogenesis (ephb3, ngf), and neurological/behavioural disorders (ngf, citron Rho-interacting kinase-like-LOC111231034) were found in the NormalF vs. WILD comparison.
Relationship between enriched pathways and PPi network analysis
The relationship among the top 20 enriched pathways obtained by ShinyGO are shown in Fig. 4. All enriched KEGG pathways were interconnected except for “cell surface receptor signalling” in DysF vs. NormalF and “ECM-receptor interaction” in both comparisons.

Relationship between enriched pathways in ovary samples from the three analysed groups. Pathways (nodes) are connected if they share 20% or more genes. Darker notes are more significantly enriched gene sets. Larger nodes represent larger gene sets.
A network based on Protein–Protein interaction (PPi) was generated to evaluate functional relations between DEGs for each comparison (Figs. 5 and 6; Supplementary Table S5). Three main protein-interaction groups emerged both in DysF vs. NormalF and DysF vs. WILD comparisons. Many of these proteins are involved in biological categories associated with extracellular matrix (ECM-receptor interaction), signaling (Enzyme-linked receptor protein signaling), and transduction (Wnt signal transduction) pathways. Moreover, several proteins associated with the Ovulation cycle category (AMH, AXL, Inhibin subunit beta Aa, Lutropin-choriogonadotropic hormone receptor, and ADAMSTs) were identified. No protein interaction groups emerged from the comparison of NormalF vs. WILD.

Protein–protein interaction (PPi) networks in DysF vs. NormalF. Networks were built using a confidence protein interaction (score = 0.5). Node background indicates gene upregulation (red, log2FC > 1.5) or downregulation (blue, log2FC < −1.5). Node contour indicates biological categories: ECM-receptor interaction (orange), Enzyme-linked receptor protein signaling pathway (blue), Wnt signal transduction pathway (green).

Protein–protein interaction (PPi) networks in DysF vs. WILD. Networks were built using a confidence protein interaction (score = 0.5). Node background indicates gene upregulation (red, log2FC > 1.5) or downregulation (blue, log2FC < 1.5). Node contour indicates biological categories: ECM-receptor interaction (orange); Enzyme-linked receptor protein signaling pathway (blue); Wnt signal transduction pathway (green).
KEGG analysis
The KEGG analysis showed DEGs involved in 111 pathways (Supplementary Table S6). Among these pathways, those likely associated with the reproductive function such as oogenesis or follicular atresia include: neuroactive ligand-receptor interaction, Steroid hormone biosynthesis, Cell cycle, Oocyte meiosis, Necroptosis, Ferroptosis, Apoptosis, Autophagy, Progesterone-mediated oocyte maturation, Endocytosis, and Phagosome, as well as Hedgehog, Apelin, PPAR, Notch, GnRH, p53, and mTOR signalling pathways. Moreover, in the DysF vs. NormalF comparison, several dysregulated gene-encoding factors associated with mammal hypogonadism, such as neuronal guanine nucleotide exchange factor and semaphorin-3F-like or polycystic ovary syndrome (PCOS), such as thymocyte selection-associated high mobility group box, chemokine (C–X–C motif) ligand 14, protein-lysine 6-oxidase-like and small glutamine rich tetratricopeptide repeat co-chaperone beta, were identified. In the same comparison, several dysregulated genes encoding factors involved in immune system functioning and inflammation, such meprin A subunit beta-like, galectin-2-like; cysteinyl leukotriene receptor 2-like and C-X-C motif chemokine 10-like, were found.
Discussion
The integration of histological and RNA-seq data from wild and hatchery-produced female greater amberjack provided novel insights into the molecular mechanisms underlying the observed oogenesis impairment in hatchery-produced fish. Histological evaluation of the ovaries revealed that three out of seven hatchery-produced greater amberjack females exhibited a reproductive dysfunction similar to that observed in individuals caught from the wild as juveniles and subsequently reared in captivity until reaching reproductive maturity15,33. Although the number of fish used in this study was limited due to the scarcity of hatchery-produced greater amberjack breeders and the challenges of collecting samples from wild breeders on commercial fishing vessels, transcriptome analyses revealed statistically significant differences in gene expression between fish showing reproductive dysfunction and those with no histological evidence of such dysfunction. Among the 19,621 genes expressed in all the three groups, approximately 4 to 6% of them were differentially expressed between the two groups, with the majority of dysregulated genes showing a downregulation in dysfunctional fish.
GO enrichment analysis
GO enrichment analysis in reproductively dysfunctional fish revealed enrichment of genes associated with secretion, extracellular matrix protein synthesis, extracellular matrix-receptor interaction, and focal adhesion. Additionally, genes encoding for growth factors such as vascular endothelial growth factors and several receptors, including the low-density lipoprotein receptor, were identified. Dysregulation of a few genes involved in morphogenesis, inflammation, and neurological/behavioral disorders suggests that fish in the NormalF group experienced limited captivity-related stress, which did not affect reproductive function. Indeed, no enriched biological categories, KEGG pathways or PPi networks were identified when comparing NormalF to WILD fish. These findings contrast with those observed in males in a previous study, where molecular evidence of reproductive dysfunction was detected even in individuals with no histological evidence of gametogenesis alterations35. However, it cannot be ruled out that hatchery-produced, farmed fish initially classified as normal, may later develop a reproductive dysfunction. Indeed, all fish from the same origin used as breeders, showed no response to spawning-induction treatments, suggesting potential dysfunctionality across all hatchery-produced broodstocks49.
Recently, a dysregulation of genes involved in extracellular matrix has been found in the testis of reproductively dysfunctional greater amberjack males35. Changes in the expression of genes encoding extracellular matrix proteins have also been observed during the testicular maturation in rainbow trout Onchorhynchus mykiss, suggesting their involvement in the reorganization of seminiferous tubules during the testicular cycle48. In humans, factors within the extracellular matrix have been reported to create the complex environment necessary for the interaction between ovarian follicles and their surroundings, ensuring follicle growth and development50. Additionally, downregulation of extracellular matrix and cell adhesion molecules has been identified in the RNA expression profile of ovaries from patients affected by PCOS, an endocrinopathic condition characterized by oligo/anovulation, hyperandrogenism and polycystic ovaries51. In the present study, the ovarian follicle degeneration characterizing the reproductive dysfunction was associated with the downregulation of several genes involved in cell-to-cell and cell-to-matrix interactions, such as genes encoding for collagens, integrins and laminins.
The observed downregulation of genes encoding for growth factors, including vascular endothelial growth factors that promote angiogenesis52, is coherent with the observed extensive atresia of vitellogenic follicles in the DysF group. Angiogenesis is a complex process regulated by a balance of pro- and anti-angiogenetic factors, and disruption of this process may result in ovarian dysfunction53. In mammals, active angiogenesis is associated with gonad development and folliculogenesis, and follicles that fail to acquire satisfactory vascular support usually undergo atresia54. In prepubertal zebrafish Danio rerio, during the transition from primary to secondary oocyte growth, biological processes related to angiogenesis, including blood vessel morphogenesis and lymph vessel formation are enriched in up-regulated genes. This suggests that formation of blood vessels is necessary to ensure sufficient supply of nutrients, hormones and oxygen to support the rapid growth of follicles in fish55.
Different lipoprotein receptors have been found to bind vitellogenin34,56,57. The downregulation of lpr5 observed in the present study may reduce the ability to uptake vitellogenin and complete vitellogenesis. In greater amberjack, significantly lower expression of lipoprotein receptor (vtgr and lrp13) gene has been observed in wild-caught, captive-reared fish compared to fish sampled in nature34. This reduced expression was associated with a reduced number of vitellogenic oocytes and, consequently, lower fecundity. Despite good egg quality observed in wild-caught greater amberjack treated with GnRHa22,23,24, fecundity was significantly lower compared to fish in the wild41.
Relationship between enriched pathways and PPi network analysis
The outputs of the ShinyGO analysis, alongside the GO enrichment analysis, clearly indicated that the observed gene dysregulation in dysfunctional greater amberjack involves many interconnected pathways related to ovarian morphogenesis and development. In the DysF group, many interconnected proteins associated with the biological categories Enzyme-linked receptor protein signalling pathway and Wnt signal transduction pathway (both involved in cell proliferation, differentiation, development, inflammatory response and apoptosis), ECM-receptor interaction and Ovulation cycle, were encoded by downregulated genes. Among the interconnected proteins involved in Ovulation cycle, AMH, AXL, Inhibin Subunit Beta A, Lutropin-choriogonadotropic hormone receptor and ADAMSTs play recognized roles in folliculogenesis and follicular atresia. Anti-Müllerian hormone (AMH) is known as a critical testicular signal driving the development of the male reproductive tract in vertebrates58. In humans, it is synthesized and secreted by granulosa cells of small ovarian follicles and is actively involved in folliculogenesis59. Recent evidence suggests that AMH may also indirectly affect folliculogenesis by modulating GnRH neuron activity, which controls pituitary gonadotropin secretion59. Moreover, during mouse embryonic development, AMH and its receptors are expressed by migratory GnRH neurons, suggesting AMH signaling involvement in congenital hypogonadotropic hypogonadism pathogenesis60. AXL, a member of the Tyro3-Axl-Mer (TAM) receptor tyrosine kinase subfamily, transduces signals from the extracellular matrix into the cytoplasm, influencing cellular functions such as growth, cell survival, cell proliferation, migration and differentiation, and has been implicated in human reproductive diseases including some forms of hypogonadotropic hypogonadism61,62. The inhibin subunit beta A (INHBA) gene encodes a member of the transforming growth factor-beta (TGF-β) superfamily and is proteolytically processed to generate a subunit of the dimeric activin and inhibin protein complexes. These complexes activate and inhibit, respectively, follicle stimulating hormone (Fsh) secretion from the pituitary gland63. Increased expression of the β-subunit of Fsh (Fshβ) was found in both young and adult inhba-deficient zebrafish; however, the adult females exhibited ovarian follicles arrested at the full-grown stage without oocyte maturation and ovulation taking place64.
The lutropin-choriogonadotropic hormone receptor, a member of the G protein coupled receptor family, mediates Lh activity in the ovary. In mammals, it is upregulated by Fsh and chorionic gonadotropin and it is downregulated by decreased receptor gene transcription and/or increased mRNA degradation following Lh stimulation (post-transcriptional downregulation)65,66. In the present study, downregulation of this gene in the dysfunctional fish suggests insufficient pituitary gonadotropin secretion, typical of captivity-induced reproductive dysfunction2.
Disintegrin and metalloproteinase enzymes belonging to the ADAMTS family are involved in tissue morphogenesis, tissue remodeling, inflammation and vascular biology, and participate in follicle rupture and ovulation processes in mammals67. Dysregulation or functional changes in ADAMTS proteases have been linked to reproductive disorders, such as PCOS and premature ovarian failure68.
KEGG analysis
Consistent with the widespread gene dysregulation, reproductive dysfunction involved many KEGG pathways. Several genes encoding cell receptors involved in the Neuroactive ligand-receptor interaction (a signaling pathway directly related to neuro function69), such as lutropin-choriogonadotropic hormone receptor, thyroid hormone receptor beta isoform X1, prostaglandin E2 receptor EP3 subtype, and d prodynorphin, were downregulated in dysfunctional fish. In rodents, prodynorphin, a preproprotein proteolytically processed to form opioid peptides that bind the kappa-type of opioid receptor70, is expressed in granulosa and luteal cells under gonadotropin regulation71. Neurons in the central nucleus of the Amygdala (CeA)72 and hypothalamic neurons73 expressing prodynorphin, kisspeptin, and neurokink B (Kdyn neurons) have been shown to regulate GnRH pulsatility. Inactivation of prodynorphin gene from multiple sources, both intrinsic and extrinsic to the CeA, induces anxiety-like behavior in mice72. It has been hypothesized that the altered GnRH pulsatility in women affected by PCOS may result from a reduced hypothalamic opioid inhibitory activity73.
In dysfunctional fish in the present study, downregulation of several genes involved in GnRH and steroid hormone biosynthesis pathways was observed. In the latter pathway, the following genes were found to be downregulated: steroid 17-alpha-hydroxylase (cyp17A1), cholesterol side-chain cleavage enzyme (cyp11a1), and cytochrome P450 family 19 subfamily A member 1 (cyp19a1; aromatase). The cyp17A1 gene encodes for the steroidogenic enzymes that produce progestins, mineralocorticoids, glucocorticoids, androgens and estrogens74. The cyp11a1 encodes for a cytochrome P450 monooxygenase that catalyzes the side-chain hydroxylation and cleavage of cholesterol to pregnenolone, the precursor of most steroid hormones75. The cyp11a1 gene produces a cytochrome P450 monooxygenase that catalyzes the aromatization of androstenedione, testosterone, and 16a-hydroxyandrostenedione76.
The failure of oocytes to undergo maturation after completion of vitellogenesis observed in cultured fish is often due to a reduced pituitary release of gonadotropins, particularly Lh, which triggers oocyte maturation via 17,20β-dihydroxy-4-pregnen-3-one (17,20β-P) synthesis1,2,7. Low plasma Lh levels have also been found in wild-caught, captive-reared fish belonging to the same stock as the parents of the fish analysed in the present study77.
Therefore, the observed dysregulation in cultured greater amberjack of genes involved in steroid synthesis, cell cycle, meiosis and progesterone-mediated oocyte maturation pathways may have originated from insufficient Lh plasma levels, in the present study. Furthermore, the widespread depression of steroid hormone synthesis, associated with the downregulation of androgen receptor-like genes in DysF fish, may have been exacerbated by the atretic loss of vitellogenic oocytes and the associated death of follicular and thecal cells. These cells are involved in testosterone synthesis and its conversion to 17-β estradiol.78.
The alteration of PPAR and Apelin signaling pathways may also have played a role in the oogenesis impairment of DysF greater amberjack. In mammals, adipokines, including apelins, are present in different ovarian cells and can affect steroidogenesis and ovulation through the activation of the nuclear transcription factors PPARS, which are involved in ovarian development and follicular atresia. Their alteration may induce infertility79,80. Although the gene expression data and histological evidence of extensive atresia of vitellogenic follicles, indicate that the observed reproductive dysfunction affected the ability of vitellogenic oocytes to proceed towards maturation, the alteration of Wnt, Notch, and Hedgehog signalling pathways, strongly suggest that the oogenesis impairment may also affect early oogenesis stages. These three pathways represent highly conserved mechanisms that contribute to stem cell maintenance and proliferation capacity81,82,83,84. Apoptosis play an important role if the regulation of fish spermatogenesis85 and a high density of apoptotic spermatogonia was observed in wild-caught Atlantic bluefin tuna Thynnus thynnus3 and greater amberjack males confined in captivity4, suggesting that the anti-apoptotic pathways protecting stem germ cells from apoptosis might not work properly under prolonged captivity-induced stress.
The dysregulation of genes encoding immune system and inflammation factors observed in the DysF vs. NormalF comparison, along with the alterations in Endocytosis and Phagosome pathways, aligns with the reported involvement of the immune system in the inflammatory process associated with follicular atresia41. Moreover, the dysregulation of Endocytosis and Phagosome pathways may be linked to the reduced ability of the ovaries in dysfunctional fish to internalize egg yolk precursors by micropinocytosis, resulting from the loss of vitellogenic oocytes through atresia.
The presence of upregulated genes associated with Necroptosis and Apoptosis pathways in dysfunctional fish may also be associated with follicular atresia. Oocyte atresia in teleost fish involves at least three different processes: autophagy (self-digestion of oocyte and follicular cell components), heterophagy (phagocytosis of egg components by granulosa cells that act as macrophages) and follicular cell death by apoptosis41. No information is available regarding the involvement of necroptosis in teleost fish atresia; in mammals, however, recent studies demonstrated that oxidative stress triggers necroptosis in granulosa cells and oocytes86.
Conclusions
In the present study, we report that 50% of hatchery-produced greater amberjack females were affected by extensive atresia of ovarian follicles in vitellogenesis. This dysfunction involved dysregulation of genes associated with many biological categories and showed molecular traits common to the PCOS and congenital hypogonadism, both conditions associated with infertility in mammals. Ongoing analyses aim to characterize gene expression dysregulation in the pituitary and brain of reproductively dysfunctional greater amberjack, with the goal of enhancing our understanding of reproductive dysfunction along the entire reproductive axis, in fish reared in captivity. This knowledge may, hopefully, lead to the development of better protocols for the reproductive control in a species of emerging interest for the aquaculture industry.
Data availability
Reads generated in this study are freely available through the SRA (Short Read Archive) database under the BioProject accession number PRJNA1056146 https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1056146. All the other data produced and/or analyzed during the current study are included in this article in Supplementary Tables.
References
Zohar, Y. & Mylonas, C. C. Endocrine manipulations of spawning in cultured fish: From hormones to genes. Aquaculture 197, 99–136 (2001).
Mylonas, C. C., Fostier, A. & Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocr. 165, 516–534 (2010).
Zupa, R. et al. Comparative analysis of male germ cell proliferation and apoptosis in wild and captive Atlantic bluefin tuna (Thunnus thynnus L.). J. Appl. Ichthyol. 29, 71–81 (2013).
Zupa, R. et al. Rearing in captivity affects spermatogenesis and sperm quality in greater amberjack Seriola dumerili (Risso, 1810). J. Anim. Sci. 95, 4085–4100 (2017).
Corriero, A. et al. Proliferation and apoptosis of male germ cells in captive Atlantic bluefin tuna (Thunnus thynnus L.) treated with gonadotropin-releasing hormone agonist (GnRHa). Anim. Reprod. Sci. 116, 346–357 (2009).
Ventriglia, G. et al. Effects of different hormonal treatments on spermatogenesis advancement in hatchery-produced greater amberjack Seriola dumerili (Risso 1810). Gen. Comp. Endocr. 348, 114447 (2023).
Mylonas, C. C. & Zohar, Y. Endocrine regulation and artificial induction of oocyte maturation and spermiation in basses of the genus Morone. Aquaculture 202, 205–220 (2001).
Sumpter, J. P., Pottinger, T. G., Rand-Weaver, M. & Campbell, P. M. The wide-ranging effects of stress in fish. In Perspectives in Comparative Endocrinology (eds. Davey, K. G. et al.). 535–538 (National Research Council of Canada, 1994).
Pankhurst, N. W. & Van der Kraak, G. Effects of stress on reproduction and growth of fish. In Fish Stress and Health in Aquaculture (eds. Iwama, G. K. et al.). 73–93 (Cambridge University Press, 1997).
Corriero, A. et al. Evidence that severe acute stress and starvation induce rapid atresia of ovarian vitellogenic follicles in Atlantic bluefin tuna, Thunnus thynnus (L.) (Osteichthyes: Scombridae). J. Fish Dis. 34, 853–860 (2011).
Zohar, Y. et al. The bioactivity of gonadotropin-releasing hormones and its regulation in the gilthead seabream, Sparus aurata: In vivo and in vitro studies. Fish. Physiol. Biochem. 7, 59–67 (1989).
Zohar, Y., Tosky, M., Pagelson, G. & Finkelman, Y. Induction of spawning in the gilthead seabream, Sparus aurata, using wD-Ala6- Pro9NEtx-LHRH: Comparison with the use of HCG. Israeli J. Aquacult. Bamidgeh 41, 105–113 (1989).
Battaglene, S. C. & Selosse, P. M. Hormone-induced ovulation and spawning of captive and wild broodfish of the catadromous Australian bass, Macquaria novemaculeata (Steindachner), (Percichthyidae). Aqua Res. 27, 191–204 (1996).
Watanabe, T. & Vassallo-Agius, R. Broodstock nutrition research on marine finfish in Japan. Aquaculture 227, 35–61 (2003).
Zupa, R. et al. Comparative study of reproductive development in wild and captive-reared greater amberjack Seriola dumerili (Risso, 1810). PLoS ONE 12(1), e0169645 (2022).
Mylonas, C. C., Scott, A. P. & Zohar, Y. Plasma gonadotropin II, sex steroids, and thyroid hormones in wild striped bass (Morone saxatilis) during spermiation and final oocyte maturation. Gen. Comp. Endocrinol. 108, 223–236 (1997).
Mylonas, C. C., Woods, L. C. III., Thomas, P. & Zohar, Y. Endocrine profiles of female striped bass (Morone saxatilis) in captivity, during post-vitellogenesis and induction of final oocyte maturation via controlled-release GnRHa-delivery systems. Gen. Comp. Endocrinol. 110, 276–289 (1998).
Rosenfeld, H. et al. GnRHa-mediated stimulation of the reproductive endocrine axis in captive Atlantic bluefin tuna, Thunnus thynnus. Gen. Comp. Endocrinol. 175, 55–64 (2012).
Corriero, A. et al. Histological study of the effects of treatment with gonadotropin-releasing hormone agonist (GnRHa) on the reproductive maturation of captive-reared Atlantic bluefin tuna (Thunnus thynnus L.). Aquaculture 272, 675–686 (2007).
Mylonas, C. C. et al. Preparation and administration of gonadotropin-releasing hormone agonist (GnRHa) implants for the artificial control of reproductive maturation in captive-reared Atlantic bluefin tuna (Thunnus thynnus thynnus). Rev. Fish. Sci. 15, 183–210 (2007).
Mylonas, C. C. et al. Reproduction of hatchery-produced meagre Argyrosomus regius in captivity III. Comparison between GnRHa implants and injections on spawning kinetics and egg/larval performance parameters. Aquaculture 448, 44–53 (2015).
Fakriadis, I., Lisi, F., Sigelaki, I., Papadaki, M. & Mylonas, C. C. Spawning kinetics and egg/larval quality of greater amberjack (Seriola dumerili) in response to multiple GnRHa injections or implants. Gen. Comp. Endocrinol. 279, 78–87 (2019).
Fakriadis, I., Miccoli, A., Karapanagiotis, S., Tsele, N. & Mylonas, C. C. Optimization of a GnRHa treatment for spawning commercially reared greater amberjack Seriola dumerili: Dose response and extent of the reproductive season. Aquaculture 521, 735011 (2020).
Fakriadis, I. et al. Control of reproduction of greater amberjack Seriola dumerili reared in aquaculture facilities. Aquaculture 519, 734880 (2020).
Bauchot, M. L. Poisson osseux, Familie Carangidae. In Fiches FAO d’Identification des Espèces pour les Besoins de la Pêche (Révision 1). Méditerraneée et mar Noire. Zone de Pêche 37. Vol. II. Vertébrés (eds. Fischer, W. et al.). 1009–1030 (FAO, 1987).
Cervigon, F. Los Peces Marinos de Venezuela. Vol. 2 (Fundacion Cientıfica Los Roques, 1993).
Smith, C. L. National Audubon Society Field Guide to Tropical Marine Fishes of the Caribbean, the Gulf of Mexico, Florida, the Bahamas, and Bermuda (eds. Alfred, A.). Vol. 120 (Knopf, Inc, 1997).
Nakada, M. Yellowtail and related species culture. In Encyclopedia of Aquaculture (ed. Stickney, R.). 1007–1036 (Wiley, 2000).
Nakada, M. Capture-based aquaculture of yellow-tail. In Capture-Based Aquaculture. Global Overview (eds. Lovatelli, A. & Holthus, P. F.). 199–215 (FAO, 2008).
Corriero, A., Wylie, M. J., Nyuji, M., Zupa, R. & Mylonas, C. C. Reproduction of greater amberjack (Seriola dumerili) and other members of the family Carangidae. Rev. Aquacult. 13, 1781–1815 (2021).
Micale, V., Genovese, L. & Greco, S. Gonadal development in cultured amberjack Seriola dumerili (Risso, 1810). Anim. Biol. 7, 125–130 (1998).
Mylonas, C. C., Papandroulakis, N., Smboukis, A., Papadaki, M. & Divanach, P. Induction of spawning of cultured greater amberjack (Seriola dumerili) using GnRHa implants. Aquaculture 37, 141–154 (2004).
Pousis, C. et al. The observed oogenesis impairment in greater amberjack Seriola dumerili (Risso, 1810) reared in captivity is not related to an insufficient liver transcription or oocyte uptake of vitellogenin. Aquac. Res. 49, 243–252 (2018).
Pousis, C. et al. Vitellogenin receptor and fatty acid profiles of individual lipid classes of oocytes from wild and captive-reared greater amberjack (Seriola dumerili) during the reproductive cycle. Theriogenology 140, 73–83 (2019).
Lavecchia, A. et al. Dysregulation of testis mRNA expression levels in hatchery-produced vs wild greater amberjack Seriola dumerili. Sci. Rep. 13, 13662 (2023).
Ventriglia, G. et al. Effects of different hormonal treatments on spermatogenesis advancement in hatchery-produced greater amberjack Seriola dumerili (Risso 1810). Gen. Comp. Endocr. 348, 114447 (2024).
Kožul, V., Skaramuca, B., Kraljević, M., Dulčić, J. & Glamuzina, B. Age, growth and mortality of the Mediterranean amberjack Seriola dumerili (Risso 1810) from the south-eastern Adriatic Sea. J. Appl. Ichthyol. 17, 134–141 (2001).
Anonymous. Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 55, 251–257 (1998).
Metcalfe, J. D. & Craig, J. F. Ethical justification for the use and treatment of fishes in research: An update. J. Fish Biol. 78(2), 393–394 (2011).
EU. Directive 2010/63/EU of the European parliament and the council of 22 September 2010 on the protection of animals used for scientific purposes. In Official Journal of the European Union L 276/33 (Animal Use): Animal Protection (2010).
Corriero, A., Zupa, R., Mylonas, C. C. & Passantino, L. Atresia of ovarian follicles in fishes, and implications and uses in aquaculture and fisheries. J. Fish Dis. 44, 1271–1291 (2021).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890 (2018).
Dobin, A. et al. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15(12), 550 (2014).
Sherman, B. T. et al. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists. Nucleic Acids Res. 50(W1), W216–W221 (2022).
Ge, S. X., Jung, D. & Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 36, 2628–2629 (2020).
Kanehisa, M. & Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 29, 28–35 (2020).
Lancerotto, S. et al. Timing of puberty in F1-generation hatchery-produced greater amberjack (Seriola dumerili). Gen. Comp. Endocr. 347, 114414 (2024).
Pors, S. E. et al. Initial steps in reconstruction of the human ovary: Survival of pre-antral stage follicles in a decellularized human ovarian scaffold. Hum. Reprod. 34(8), 1523–1535 (2019).
Hassani, F. et al. Downregulation of extracellular matrix and cell adhesion molecules in cumulus cells of infertile polycystic ovary syndrome women with and without insulin resistance. Cell J. 21(1), 35–42 (2019).
Shibuya, M. Vascular endothelial growth factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: A crucial target for anti- and pro-angiogenic therapies. Genes Cancer 2(12), 1097–1105 (2011).
Osz, K., Ross, M. & Petrik, J. The thrombospondin-1 receptor CD36 is an important mediator of ovarian angiogenesis and folliculogenesis. Reprod. Biol. Endocrinol. 12, 21 (2014).
Brown, H. M. & Russell, D. L. Blood and lymphatic vasculature in the ovary: Development, function and disease. Hum. Reprod. Update 20, 29–39 (2014).
Zhu, B., Pardeshi, L., Chen, Y. & Ge, W. Transcriptomic analysis for differentially expressed genes in ovarian follicle activation in the zebrafish. Front. Endocrinol. 9, 00593 (2018).
Pousis, C. et al. Expression of vitellogenin receptor gene in the ovary of wild and captive Atlantic bluefin tuna (Thunnus thynnus). Anim. Reprod. Sci. 132, 101–110 (2012).
Babio, L., Damsteegt, E. L. & Lokman, P. M. Lipoprotein receptors in ovary of eel, Anguilla australis: Molecular characterisation of putative vitellogenin receptors. Fish Physiol. Biochem. 49(1), 117–137 (2023).
Münsterberg, A. & Lovell-Badge, R. Expression of the mouse anti-Müllerian hormone gene suggests a role in both male and female sexual differentiation. Development 113(2), 613–624 (1991).
Silva, M. S. B. & Giacobini, P. New insights into anti-Müllerian hormone role in the hypothalamic–pituitary–gonadal axis and neuroendocrine development. Cell. Mol. Life Sci. 78, 1–16 (2021).
Malone, S. A. et al. Defective AMH signaling disrupts GnRH neuron development and function and contributes to hypogonadotropic hypogonadism. eLife 8, e47198 (2019).
Yatsenko, S. A. & Rajkovic, A. Genetics of human female infertility. Biol. Reprod. 101, 549–566 (2019).
Mohammadzadeh, P. & Amberg, G. C. AXL/Gas6 signaling mechanisms in the hypothalamic–pituitary–gonadal axis. Front. Endocrinol. 14, 1212104 (2023).
Lu, C. et al. Inhibin A inhibits follicle-stimulating hormone (FSH) action by suppressing its receptor expression in cultured rat granulosa cells. Mol. Cell. Endocrinol. 298, 48–56 (2009).
Lu, H., Zhao, C., Zhu, B., Zhang, Z. & Ge, W. Loss of inhibin advances follicle activation and female puberty onset but blocks oocyte maturation in zebrafish. Endocrinology 161(12), bqaa184 (2020).
Lu, D. L., Peegel, H., Mosier, S. M. & Menon, K. M. Loss of lutropin/human choriogonadotropin receptor messenger ribonucleic acid during ligand-induced down-regulation occurs post transcriptionally. Endocrinology 132(1), 235–240 (1993).
Menon, K. M., Munshi, U. M., Clouser, C. L. & Nair, A. K. Regulation of luteinizing hormone/human chorionic gonadotropin receptor expression: A perspective. Biol. Reprod. 70(4), 861–866 (2004).
Baker, S. J. C. & Van Der Kraak, G. ADAMTS1 is regulated by the EP4 receptor in the zebrafish ovary. Gen. Comp. Endocr. 311, 13835 (2021).
Russell, D. L., Brown, H. M. & Dunning, K. R. ADAMTS proteases in fertility. Matrix Biol. 44–46, 54–63 (2015).
Duan, J. et al. Low-dose exposure of silica nanoparticles induces cardiac dysfunction via neutrophil-mediated inflammation and cardiac contraction in zebrafish embryos. Nanotoxicology 10, 575–585 (2016).
Fricker, L. D., Margolis, E. B., Gomes, I. & Devi, L. A. Five decades of research on opioid peptides: Current knowledge and unanswered questions. Mol. Pharmacol. 98, 96–108 (2020).
Kaynard, A. H., McMurray, C. T., Douglass, J., Curry, T. E. & Melner, M. H. Regulation of prodynorphin gene expression in the ovary: Distal DNA regulatory elements confer gonadotropin regulation of promoter activity. Mol. Endocrinol. 6, 2244–2256 (1992).
Baird, M. A., Hsu, T. Y., Wang, R., Juarez, B. & Zweifel, L. S. κ opioid receptor-dynorphin signaling in the central amygdala regulates conditioned threat discrimination and anxiety. eNeuro 8(1), ENEURO.0370-20.2020 (2021).
Garg, A., Patel, B., Abbara, A. & Dhillo, W. S. Treatments targeting neuroendocrine dysfunction in polycystic ovary syndrome (PCOS). Clin. Endocrinol. 97(2), 156–164 (2022).
Uno, T., Ishizuka, M. & Itakura, T. Cytochrome P450 (CYP) in fish. Environ. Toxicol. Pharm. 34, 1–13 (2012).
Strushkevich, N. et al. Structural basis for pregnenolone biosynthesis by the mitochondrial monooxygenase system. Proc. Natl. Acad. Sci. USA 108(25), 10139–10143 (2011).
Corbin, C. J. et al. Isolation of a full-length cDNA insert encoding human aromatase system cytochrome P-450 and its expression in nonsteroidogenic cells. Proc. Natl. Acad. Sci. USA 85(23), 8948–8952 (1988).
Fakriadis, I. et al. Gonadotropin expression, pituitary and plasma levels in the reproductive cycle of wild and captive-reared greater amberjack (Seriola dumerili). Gen. Comp. Endocr. 350, 114465 (2024).
Lubzens, E., Young, G., Bobe, J. & Cerdà, J. Oogenesis in teleosts: How fish eggs are formed. Gen. Comp. Endocrinol. 165, 36789 (2010).
Cui, Y. et al. Loss of the peroxisome proliferation-activated receptor gamma (PPARgamma) does not affect mammary development and propensity for tumor formation but leads to reduced fertility. J. Biol. Chem. 277, 17830–17835 (2002).
Dupont, J., Reverchon, M., Cloix, L., Froment, P. & Ramé, C. Involvement of adipokines, AMPK, PI3K and the PPAR signaling pathways in ovarian follicle development and cancer. Int. J. Dev. Biol. 56(10–12), 959–967 (2012).
Jiang, Y. et al. Hedgehog pathway inhibition causes primary follicle atresia and decreases female germline stem cell proliferation capacity or stemness. Stem Cell Res. Ther. 10, 198 (2019).
Moreno Acosta, O. D., Boan, A. F., Hattori, R. S. & Fernandino, J. I. Notch pathway is required for protection against heat stress in spermatogonial stem cells in medaka. Fish Physiol. Biochem. 49, 487–500 (2023).
Upadhyay, M., Kuna, M., Tudor, S., Martino Cortez, Y. & Rangan, P. A switch in the mode of Wnt signaling orchestrates the formation of germline stem cell differentiation niche in Drosophila. PLoS Genet. 14(1), 7154 (2018).
Yavropoulou, M. P., Maladaki, A. & Yovos, J. G. The role of Notch and Hedgehog signaling pathways in pituitary development and pathogenesis of pituitary adenomas. Hormones 14(1), 5–18 (2015).
Corriero, A. et al. Germ cell proliferation and apoptosis during different phases of swordfish (Xiphias gladius L.) spermatogenetic cycle. J. Fish Biol. 70, 83–99 (2007).
Chaudhary, G. R. et al. Necroptosis in stressed ovary. J. Biomed. Sci. 26, 11 (2019).
Acknowledgements
The authors are grateful to Mr Tasos Raftopoulos of Argosaronikos Fishfarming S.A. (Greece) for providing the hatchery-produced greater amberjack and for assistance during biological sampling. Thanks are also expressed to Mr Peppe, Giovanni and Vincenzo Billeci, and all the crew of the purse-seine fishing vessel ‘Graziella’ (Italy) for their friendly collaboration and hospitality on board during the sampling of wild greater amberjack. Thanks are due to Rezart Cuko for his technical support in the preparation of histological slides.
Funding
This work was funded by HORIZON EUROPE European Research Council (Grant no. 862658) and by ELIXIR-IT though the empowering project ELIXIRNextGenIT (Grant no. IR0000010).
Author information
Authors and Affiliations
Contributions
Project ideation: C.P., C.C.M., G.P. and A.C. Biological sampling: C.P., C.C.M., R.Z. and A.C. Molecular analyses: C.D.V. and C.M. Elaboration of RNAseq data: A.L., L.M., S.N.C. and E.P. Histological analysis: R.Z. and G.V. Figure preparation: A.L., R.Z and A.C. Funding: C.C.M., G.P. and A.C. Manuscript writing (first draft): A.L. and A.C. Manuscript review: all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lavecchia, A., De Virgilio, C., Mansi, L. et al. Comparison of ovarian mRNA expression levels in wild and hatchery-produced greater amberjack Seriola dumerili. Sci Rep 14, 18034 (2024). https://doi.org/10.1038/s41598-024-69091-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-69091-2
- Springer Nature Limited