
Abstract
Button-press measures of response time (RT) and accuracy have long served a central role in psychological research. However, RT and accuracy provide limited insight into how cognitive processes unfold over time. To address this limitation, researchers have used hand-tracking techniques to investigate how cognitive processes unfold over the course of a response, are modulated by recent experience, and function across the lifespan. Despite the efficacy of these techniques for investigating a wide range of psychological phenomena, widespread adoption of hand-tracking techniques within the field is hindered by a range of factors, including equipment costs and the use of specialized software. Here, we demonstrate that the behavioral dynamics previously observed with specialized motion-tracking equipment in an Eriksen flanker task can be captured with an affordable, portable, and easy-to-assemble response box. Six-to-eight-year-olds and adults (N = 90) completed a computerized version of the flanker task by pressing and holding a central button until a stimulus array appeared. Participants then responded by releasing the central button and reaching to press one of two response buttons. This method allowed RT to be separated into initiation time (when the central button was released) and movement time (time elapsed between initiation and completion of the response). Consistent with previous research using motion-tracking techniques, initiation times and movement times revealed distinct patterns of effects across trials and between age groups, indicating that the method used in the current study presents a simple solution for researchers from across the psychological and brain sciences looking to move beyond RTs.
Similar content being viewed by others


Introduction
Button-press measures of response time (RT) and accuracy have served as cornerstones of quantitative research in psychology since the mid-twentieth century. However, such measures provide limited insight into how the processes underlying perception, cognition, and action unfold over time. Consequently, researchers have developed sophisticated computational1,2,3,4,5,6, neural7,8,9,10,11, and behavioral methods12,13,14,15,16,17 to shed light on how cognitive processes unfold over the course of a response (within-trial dynamics), are modulated by recent experience (cross-trial dynamics), and function across the life span (developmental dynamics).
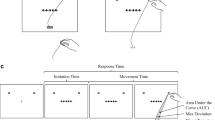
The limitations of button-press measures of RT and accuracy are illustrated by recent research using hand-tracking techniques to investigate the dynamics of attention and control18–22. In contrast to standard button-press measures, techniques such as mouse tracking and reach tracking provide insight into the temporal and spatial dynamics of performance by recording the path that a participant’s hand travels to reach a response target (see Fig. 1a,b). For example, these techniques allow RT to be deconstructed into measures of initiation time (IT; the time elapsed between stimulus onset and movement initiation) and movement time (MT; the time elapsed between movement initiation and response completion).

(a) Illustration of a mouse-tracking version of the flanker task. In the task, participants are instructed to respond according to a centrally presented target stimulus and to ignore surrounding distractor stimuli. Participants initiate each trial from a starting location at the bottom center of the display. Following stimulus presentation, participants navigate the mouse cursor to a response target located at the top of the screen. (b) Illustration of a reach-tracking version of the task. Participants initiate each trial by placing their finger on a designated starting marker place on the table in front of them. Following stimulus presentation, participants respond by reaching to touch a response target on the digital display. Hand movements can be recorded with optical tracking systems or electromagnetic tracking systems, both of which usually require the participant to wear a motion tracking sensor on their pointing finger. (c) Illustration of the button-release-and-press method used in the current study. Participants initiate each trial by holding down a “start” button at the bottom center of a response box. Following stimulus presentation, participants release the “start” button and respond by pressing one of two response buttons located toward the top of the response box. IT is measured as the time elapsed between stimulus presentation and the release of the “start” button, whereas MT is measured as the time elapsed between the release of the “start” button and the pressing of a response button. Portions of this figure were adapted from Erb40 with permission from the author.
Recent reach-tracking studies demonstrate that the patterns of effects observed in RTs in common congruency tasks such as the Eriksen flanker task23 reflect distinct patterns of effects in ITs and MTs18,24,25. In the flanker task, participants indicate which direction a centrally presented target arrow is pointing on each trial (see Fig. 1). On congruent trials, the target arrow is surrounded by distractor arrows that cue the same response (e.g., >>>>>). On incongruent trials, the distractor arrows cue a different response than the target arrow (e.g., <<><<). A congruency effect is standardly observed in the task, with higher error rates and slower response times on incongruent relative to congruent trials (I–C).
Additionally, a congruency sequence effect (CSE) (or Gratton effect)26 is frequently observed in the flanker task, with a larger congruency effect observed in trials preceded by a congruent trial (cI–cC, where the lowercase letter denotes the congruency of the previous trial) relative to trials preceded by an incongruent trial (iI–iC) (see Fig. 2a). Multiple studies have found that the CSE observed in two-response versions of the flanker task is driven by response repetition trials in which the response of the previous trial is required on the current trial (see Fig. 2b)24,27,28. In contrast, RTs on response alternation trials have been found to feature main effects of the current trial’s congruency (slower on incongruent trials than congruent trials) and the previous trial’s congruency (slower on trials preceded by an incongruent trial relative to trials preceded by a congruent trial).

(a) Illustration of the congruency sequence effect (CSE) observed in response times (RTs) in a two-response version of the flanker task, with a smaller congruency effect on trials preceded by an incongruent trial (iI and iC trials) than on trials preceded by a congruent trial (cI and cC trials). (b) Illustration of the RT effects observed in the same task when trials featuring the opposite response of the preceding trial (response alternation trials) are separated from trials featuring the same response as the preceding trial (response repetition trials). Note that the CSE pattern is only observed in response repetition trials.
Reach tracking investigations of the flanker task demonstrate that the CSE observed in RTs in two-response versions of the task (see the left panel of Fig. 3a) reflects the combination of two distinct patterns of effects: one observed in ITs and another observed in MTs18,24. Specifically, ITs reveal a main effect of current and previous congruency in response alternation and response repetition trials (see the left panel of Fig. 3b), whereas MTs reveal a main effect of current congruency in response alternation trials and a CSE in response repetition trials (see left panel of Fig. 3c). Erb et al.19 suggested that the patterns of effects observed in ITs and MTs reflect two dissociable processes underlying cognitive control: a threshold adjustment process involving the global inhibition of motor output in response to signals of conflict29,30,31,32,33,34, and a controlled selection process that regulates top-down attentional resources to bias competing response activations towards the correct response, respectively35,36. Thus, in addition to demonstrating that RT patterns commonly obscure underlying effects of interest15, collecting separate measures of IT and MT presents new opportunities to clarify how the processes that underlie decision behavior unfold over time in different tasks, age groups, and individuals24,25,37,38,39,40,41.

(a) Response time (RT), (b) initiation time (IT), and (c) movement time (MT) data from a two-response version of the flanker task with 135 participants (45 6- to 8-year-olds, 45 10- to 12-year-olds, and 45 adults) by Erb and Marcovitch24 collected using an electromagnetic reach-tracking system (left panel) and data obtained in the current study using a button-release-and-press method from adult participants (middle panel) and 6- to 8-year-old participants (right panel). Error bars denote standard errors. Data obtained by Erb and Marcovitch24 presented with permission from the authors.
Although measuring the temporal and spatial characteristics of hand movements (i.e., manual dynamics) has proven instrumental to studying topics across perceptual, cognitive, social, and developmental psychology17,18,19,20,21,22,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57, hand-tracking techniques remain comparatively rare in psychological research relative to standard button-press measures. This reliance on button-press measures can be attributed to a range of factors, including the cost and lack of portability associated with optical or electromagnetic motion-tracking systems, the use of specialized software, and ongoing discussions concerning the analysis and interpretation of hand-tracking data58,59,60,61.
In contrast to optical or electromagnetic motion-tracking systems, mouse tracking presents an easy to use, affordable, and portable solution for researchers who want to measure manual dynamics13,14,53,56,59. However, the extent to which mouse tracking can be used to capture the patterns of effects observed in ITs and MTs in reach-tracking studies remains unclear. This lack of clarity is partly because most mouse-tracking studies have used a dynamic starting procedure in which participants initiate their movement before the operative stimulus is presented, thereby preventing measurement of IT13,62. As such, ITs in tasks featuring dynamic starting procedures are commonly used to define exclusion criteria in mouse tracking paradigms, but are not commonly analyzed.
A recent mouse-tracking study by Ye and Damian63 used a static starting procedure in which participants initiated their movements after the operative stimulus was presented in three different congruency tasks, including the flanker task. Consistent with the reach-tracking studies reviewed above, the researchers observed a main effect of previous congruency, with slower initiation times on trials preceded by an incongruent as opposed to a congruent trial. However, no effect of current congruency was observed in initiation times, suggesting that mouse tracking may be less effective at capturing the different patterns of effects observed in ITs and MTs in reach-tracking tasks. Additionally, it is unclear how effective mouse-tracking tasks are for gathering data from children, given that children can vary greatly with regard to their skills with a computer mouse64. This consideration is especially relevant given that mouse-tracking studies require participants to perform visuomotor transformations to link their hand movements to the movements of a mouse cursor21.
In light of (a) the substantial barriers to adopting optical or electromagnetic motion-tracking techniques and (b) the potential limitations of mouse tracking outlined above, the current study investigates the extent to which the within-trial, cross-trial, and developmental dynamics observed with specialized hand-tracking systems24 can be captured using a button-release-and-press method in which participants respond by releasing a “start” button and then pressing one of two response buttons (see Fig. 1c)65,66,67,68,69. Demonstrating that this method can be used to capture the behavioral dynamics observed with more specialized systems would provide a simple, accessible solution for researchers across the behavioral and brain sciences looking to move beyond RTs.
Method
Preregistration and data availability
This study was preregistered through the Open Science Framework website on September 23rd, 2019 using the AsPredicted.org template (https://osf.io/bqsf3). We present our preregistration along with accompanying comments in Sect. 1 of the Supplementary Materials. The data and analysis files for the current study are available at https://osf.io/zpyvf/?view_only=761bd50ec9714aaba6b6fcc2101651ff. As detailed in the preregistration, the current study also collected eye-tracking data. For the purposes of this report, we focus on the hand movement data and report the full set of preregistered analyses (including analyses related to eye movements) in Sect. 2 of the Supplementary Materials.
Participants
The final sample consisted of 15 6-year-olds (M = 77.1 months, SD = 3.0, 8 female), 15 7-year-olds (M = 88.5 months, SD = 2.7, 8 female), 15 8-year-olds (M = 101.0 months, SD = 3.5, 5 female) and 45 adults (M = 20.7 years, SD = 2.0, 27 female). To be eligible for the study, participants had to be right-handed, have normal or corrected-to-normal vision, understand English, be able to perform normal reaching movements, and have no diagnosis of a social or cognitive impairment. Adults provided informed consent prior to testing. Child participants provided their assent, and informed consent was obtained from a legal guardian prior to testing. One additional adult was tested but excluded from the final dataset due to a software error. Three additional children were tested but excluded from the final dataset due to failure to complete the task. Adults received course credit or $15 NZD for participating. Child participants were compensated with a small prize worth approximately $5 (e.g., a plush toy). The families of participating children were also entered into a drawing for the chance to win a $25 supermarket voucher. Testing was conducted at the University of Auckland City campus and the study was approved by the University of Auckland Human Participants Ethics Committee (UAHPEC). Testing was performed in accordance with APA Ethical Principles.
Apparatus
The task was displayed using a 53 cm × 29.6 cm digital monitor. Participants were seated at a small table facing the monitor. Participants made their responses using three buttons (3.0 cm in diameter) located toward the top left, top right, and bottom center of a 36.0 cm × 23.0 cm response box positioned centrally on the table in front of them (see Fig. 1c). Section 3 of the Supplementary Materials provides details regarding the construction of the response box, including instructions for researchers interested in constructing a custom response box. The task was designed and run using E-Prime® 3.0.
Procedure
Participants completed a computerized two-response version of the Eriksen flanker task featuring arrow stimuli (see Fig. 1). Participants initiated each trial by pressing and holding a central “start” button. After 1 s, the stimulus array appeared. Participants then made their response by releasing the “start” button and reaching to press either the right or left response button. Participants in all age groups were first given a verbal explanation of the task instructions. Child participants were shown a printed picture of the four possible stimulus arrays and were asked to demonstrate (using the response box) which response was correct in each case. The experimenter demonstrated the procedure for adult participants in the first two trials of the practice block. Participants then completed a block of 16 practice trials. During the practice trials, the experimenter (seated next to the participant) provided guidance and corrective instructions to participants in all age groups (e.g., “remember to look at the middle arrow” or “keep holding the start button down until the arrows appear”). A high tone (600 Hz for 200 ms) followed correct responses and a low tone (300 Hz for 200 ms) followed incorrect responses. There was no time limit for responses in the practice trials.
Adults completed the experimental blocks with a black curtain drawn between them and the experimenter. This curtain was left open for child participants after pilot testing revealed the need for continued monitoring and scaffolding from the experimenter (e.g., “remember to only press the buttons with your right hand”). Participants were presented with four blocks of 48 trials, for a total of 192 experimental trials. Twenty-one adult participants and 13 child participants were presented with 41 trials in the third block due to a coding error. Each block consisted of an equal number of congruent and incongruent trials, and an equal number of trials wherein the target arrow cued the left and right responses. Trial order was randomized for each block of trials. If the participant provided an incorrect response or took longer than 4 s to respond, the trial ended, and a low tone sounded. Correct responses in the experimental trials were not followed by any feedback. Participants were instructed to respond as quickly and accurately as possible.
At the end of each experimental block, the word “Rest” appeared in the centre of the screen. For adult participants, the experimenter verbally confirmed with the participant that they were ready to start the next block after approximately 5 s. Child participants were given the opportunity to take a break after each experimental block. The experimenter initiated the next block after the child confirmed that they were ready to continue. Trials containing an invalid response (e.g., the participant pressed the “start” button instead of the left or right button after stimulus presentation) and trials in which no response was provided within the 4 s limit, were excluded from analysis. Error rate, RT, IT and MT measures for each trial were obtained directly from E-prime® outputs. IT was defined as the time elapsed between the appearance of the stimulus array and release of the “start” button. MT was defined as the time elapsed between release of the “start” button and response completion. RT was computed for each trial by summing IT and MT.
Results
The first trials of each block was excluded from all analyses. Error rates were at floor for adult participants (M = 0.47%, SD = 0.01). Child participants made significantly more errors than adults [M = 4.96%, SD = 0.07, F(1, 88) = 18.51, p < .001]. Error rates for child participants also revealed a significant congruency effect F(1, 44) = 11.31, p = .002, with more errors in incongruent trials (M = 7.81%, SD = 0.13) relative to congruent trials (M = 2.18%, SD = 0.03). Trials featuring an error, and trials preceded by an error were excluded from the subsequent analyses. Log transformations were applied to RT, IT, and MT to minimize the effect of processing speed differences between children and adults. These measures were then analyzed using mixed analyses of variance (ANOVAs), featuring age group (children, adults) as a between-subjects factor, and previous congruency (c, i), current congruency (C, I), and response type (repetition, alternation) as within-subjects factors. Bonferroni corrections were applied to all post-hoc analyses to correct for multiple comparisons. Results of ANOVAs performed on log transformed and raw data are available in Sects. 4 and 5 of the Supplementary Materials, respectively. Effect sizes in the figures and text below is presented in milliseconds (without the log transformation) to aid interpretability.
In keeping with previous findings, RTs revealed a significant CSE [58 ms; F(1,88) = 32.08, p < .001, ηp2 = 0.27], with a larger congruency effect on trials preceded by a congruent trial (220 ms) than trials preceded by an incongruent trial (162 ms). The CSE was specific to response repetition trials [98 ms; F(1,88) = 38.02, p < .001, ηp2 = 0.30], with response alternation trials revealing main effects of current congruency [186 ms; F(1,88) = 133.34, p < .001, ηp2 = 0.60] and previous congruency [17 ms; F(1,88) = 12.12, p < .001, ηp2 = 0.12] but no interaction between the two, consistent with previous research (see Fig. 3a)24,27,28.
The overall pattern observed in RTs reflected the sum of two distinct trial sequence effects in ITs and MTs. Specifically, ITs revealed main effects of current congruency [94 ms; F(1,88) = 161.74, p < .001, ηp2 = 0.65] and previous congruency [10 ms; F(1,88) = 23.14, p < .001, ηp2 = 0.21; see Fig. 3b]. This pattern of effects has been previously proposed to reflect a threshold adjustment process in which signals of conflict lead to higher response thresholds and, consequently, longer initiation times on incongruent trials and trials following an incongruent trial24. MTs revealed a significant interaction among current congruency, previous congruency, and response repetition type [F(1,88) = 18.59, p < .001, ηp2 = 0.17]. Follow-up tests revealed a main effect of current congruency on response alternation trials [91 ms; F(1,88) = 70.92, p < .001, ηp2 = 0.44]. In contrast, response repetition trials revealed a significant interaction between current and previous congruency [F(1,88) = 45.88, p < .001, ηp2 = 0.34]. Follow-up tests revealed slower MTs on cI-r trials (454 ms) than iI-r trials (402 ms), with a significantly smaller difference between cC-r (317 ms) and iC-r (332 ms) trials (see Fig. 3c; t(89) = 6.77, p < .001). Erb and Marcovitch24 also observed this pattern of effects in MTs and attributed the difference between cI-r and iI-r trials to feature-integration effects occurring when the stimulus and response features of the previous trial partially overlap (cI-r trials) or fully overlap (iI-r trials) with those of the current trial28,70,71.
Several developmental differences of note were also observed. The congruency effect observed in RTs decreased significantly with age [children: 323 ms; adults: 61 ms; F(1,88) = 46.02, p < .001, ηp2 = 0.34; see Fig. 3a], with a significant age-related decrease in both ITs (children: 162 ms; adults: 27 ms; see Fig. 3b) and MTs (children: 161 ms; adults: 35 ms; see Fig. 3c). The size of the CSE also significantly decreased with age in both RTs [children: 104 ms; adults: 12 ms; F(1,88) = 12.72, p = .001, ηp2 = 0.13] and MTs [children: 61 ms; adults: 11 ms; F(1,88) = 7.36, p = .008, ηp2 = 0.08]. Children’s performance in the current study diverged from the findings of Erb and Marcovitch24 in two ways. Firstly, children’s ITs in the current study revealed a CSE [F(1,44) = 10.14, p = .003, ηp2 = 0.19], and did not reveal an effect of previous congruency on incongruent trials, indicating that patterns in ITs and MTs are less distinct in children than adults. Secondly, we did not observe the four-way interaction in MTs reported by Erb and Marcovitch24 in which age-related decreases in the CSE were specific to response repetition trials.
Discussion
The results of the current study demonstrate that a simple button-release-and-press method can be used to capture distinct patterns of effects in ITs and MTs that are otherwise obscured in RTs. Specifically, RTs revealed main effects of current and previous congruency in response alternation trials and a CSE in response repetition trials, consistent with previous reach-tracking and button-press research24,27,28. The pattern observed in RTs was demonstrated to reflect distinct patterns of effects in ITs and MTs, consistent with previous work indicating that these measures can be used to capture the functioning of the threshold adjustment process and controlled selection process, respectively18,24. Thus, our findings indicate that a simple, affordable, and portable response box can be used to capture distinct patterns of effects in RTs, ITs, and MTs in a manner comparable to specialized optical or electromagnetic tracking systems.
In contrast to the reach tracking results from Erb and Marcovitch24, the button-release-and-press method used in the current study proved to be less effective for capturing distinct effects in ITs and MTs in children. We suspect that this difference emerged because children in the current study often looked down to the response box before initiating their response, possibly delaying their movement and resulting in a blending of the typical IT and MT patterns. This possibility is supported by the observation that children spent a smaller proportion of time during experimental trials with their eyes on-screen (M = 0.86, SD = 0.25) than adults (M = 0.99, SD = 0.05). To address this, future studies may incorporate additional practice using the button box for child participants prior to experimental trials. Alternatively, to reduce the possibility of split attention and to make the procedure more similar to past hand-tracking research with children, future studies could have children respond by reaching to images on a touchscreen display instead of separate buttons.
Although the patterns of effects observed in adults in the current study correspond closely to the patterns observed in previous research using electromagnetic motion-tracking systems, the strongest demonstration of the button-release-and-press method would come from a study in which the same group of participants complete the same tasks with both methods. As noted in the “Introduction”, it is not clear at present whether mouse tracking can be used to deconstruct RTs in the same manner as the button-release-and-press method tested in the current study63. Consequently, future research should directly compare mouse tracking, 3-dimensional reach tracking, and the button-release-and-press method in tasks targeting attention and control.
A recent study by Moher and Song72 compared 3-dimensional reach tracking, mouse tracking, and stylus tracking in a task in which a target stimulus would occasionally change locations after the participated initiated a movement. The researchers found that the three methods generated results that were largely comparable. However, hand movements measured with the 3-dimensional reach-tracking system revealed (a) faster initiation latencies than movements measured with a stylus and (b) more curved movement trajectories than movements measured with a stylus or computer mouse. These observations led Moher and Song to conclude that, “3D reach trackers may be ideal for observing fast, subtle changes in internal decision-making processes compared to other devices” (p. 2558). Although the button-release-and-press method used in the current study does not capture the spatial dynamics of responding, the results of Moher and Song suggest that 3-dimensional hand movements like those required in the current study may be particularly useful for capturing initiation time effects.
Our findings present important implications for research across the behavioral and brain sciences that collects button-press measures of RT. For example, many of the standardized cognitive assessments used to measure developmental and individual differences73 could incorporate a button-release-and-press method to collect measures of IT and MT. This approach also presents an alternative solution to the challenges of interpreting developmental and individual differences in performance when speed-accuracy trade-off effects may be present74,75 given that the method allows participants to detect and override incorrect responses, resulting in very low error rates.
This approach can also be easily paired with electroencephalography (EEG) to shed new light on the link between the brain and behavior. Electromagnetic position and orientation recording systems such as those used by Erb and Marcovitch24 are sensitive to ferromagnetic materials and may therefore be difficult to pair with EEG. Although researchers have incorporated more continuous measures of behavior into EEG studies using electromyography (EMG)8,76 mouse tracking77, and dynamometers78,79, the neural correlates of the threshold adjustment and controlled selection processes captured with reach tracking are currently unclear. The button-release-and-press method used in the current study is compatible with any equipment that can be used with a standard keyboard, including EEG. Consequently, our research group is currently using the approach to target neural activity preceding movement initiation in EEG investigations of the flanker23 and Simon tasks80.
Conclusion
RTs have long served as a foundational behavioral measure in the psychological and brain sciences. Although the current study focused on the dynamics of cognitive control, the ability to deconstruct RTs into ITs and MTs could provide important insights into the cognitive processes underlying a wide range of phenomena, including perception, memory, language, numerical cognition, social reasoning, and decision making. Additionally, the release-and-press method used in the current study can be paired with other techniques such as eye tracking and EEG to shed new light on how manual, oculomotor, and neural dynamics are linked81,82. Although hand-tracking techniques have demonstrated great potential for studying the mind in action17,18,19,20,21,22,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57, these techniques have yet to be widely adopted within psychological research. The results of the current study demonstrate that a simple, portable, and low-cost button-release-and-press method can provide a robust, accessible solution for researchers looking to move beyond response times.
References
Brown, S. & Heathcote, A. A ballistic model of choice response time. Psychol. Rev. 12(1), 117–128. https://doi.org/10.1037/0033-295x.112.1.117 (2005).
De Boeck, P. & Jeon, M. An overview of models for response times and processes in cognitive tests. Front. Psychol. 10, 1–10. https://doi.org/10.3389/fpsyg.2019.00102 (2019).
Panis, S., Schmidt, F., Wolkersdorfer, M. P. & Schmidt, T. Analyzing response times and other types of time-to-event data using event history analysis: A tool for mental chronometry and cognitive psychophysiology. i-Percept 11(6), 1–24. https://doi.org/10.1177/2041669520978673 (2020).
Rouder, J., Province, J., Morey, R., Gomez, P. & Heathkote, A. The lognormal race: A cognitive-process model of choice and latency with desirable psychometric properties. Psychometrika 80, 491–513. https://doi.org/10.1007/s11336-013-9396-3 (2015).
Ratcliff, R., Smith, P. L., Brown, S. D. & McKoon, G. Diffusion decision model: Current issues and history. Trends Cogn. Sci. 20(4), 260–281. https://doi.org/10.1016/j.tics.2016.01.007 (2016).
Servant, M. & Evans, N. J. A diffusion model analysis of the effects of aging in the Flanker Task. Psychol. Aging. 35(6), 831–849. https://doi.org/10.1037/pag0000546.supp (2020).
Clawson, A., Clayson, P. E., Keith, C. M., Catron, C. & Larson, M. J. Conflict and performance monitoring throughout the lifespan: An event-related potential (ERP) and temporospatial component analysis. Biol. Psychol. 124, 87–99. https://doi.org/10.1016/j.biopsycho.2017.01.012 (2017).
Coles, M. G., Gratton, G., Bashore, T. R., Eriksen, C. W. & Donchin, E. A psychophysiological investigation of the continuous flow model of human information processing. J. Exp. Psychol. Hum. Percept. Perform. 11(5), 529–553. https://doi.org/10.1037/0096-1523.11.5.529 (1985).
Lo, S. L. A meta-analytic review of the event-related potentials (ERN and N2) in childhood and adolescence: Providing a developmental perspective on the conflict monitoring theory. Dev. Rev. 48, 82–112. https://doi.org/10.1016/j.dr.2018.03.005 (2018).
Schneider, D., Zickerick, B., Thönes, S. & Wascher, E. Encoding, storage, and response preparation—distinct EEG correlates of stimulus and action representations in working memory. Psychophysiology 57(6), 1–15. https://doi.org/10.1111/psyp.13577 (2020).
Servant, M., White, C., Montagnini, A. & Burle, B. Linking theoretical decision-making mechanisms in the Simon task with electrophysiological data: A model-based neuroscience study in humans. J. Cogn. Neurosci. 28(10), 1501–1521. https://doi.org/10.1162/jocn_a_00989 (2016).
Ficarella, S. C., Rochet, N. & Burle, B. Becoming aware of subliminal responses: an EEG/EMG study on partial error detection and correction in humans. Cortex 120, 443–456. https://doi.org/10.1016/j.cortex.2019.07.007 (2019).
Freeman, J. B. & Ambady, N. MouseTracker: Software for studying real-time mental processing using a computer mouse-tracking method. Behav. Res. 42(1), 226–241. https://doi.org/10.3758/brm.42.1.226 (2010).
Scherbaum, S. & Dshemuchadse, M. Psychometrics of the continuous mind: Measuring cognitive sub-processes via mouse tracking. Mem. Cogn. 48(3), 436–454. https://doi.org/10.3758/s13421-019-00981-x (2019).
Song, J. H. & Nakayama, K. Hidden cognitive states revealed in choice reaching tasks. Trends Cogn. Sci. 13(8), 360–366. https://doi.org/10.1016/j.tics.2009.04.009 (2009).
Weissman, D. H. Let your fingers do the walking: Finger force distinguishes competing accounts of the congruency sequence effect. Psychon. Bull. Rev. 26(5), 1619–1626. https://doi.org/10.3758/s13423-019-01626-5 (2019).
Wispinski, N. J., Gallivan, J. P. & Chapman, C. S. Models, movements, and minds: Bridging the gap between decision making and action. Ann. N. Y. Acad. Sci. 1426(1), 30–51. https://doi.org/10.1111/nyas.13973 (2018).
Erb, C. D., Moher, J., Sobel, D. M. & Song, J. H. Reach tracking reveals dissociable processes underlying cognitive control. Cognition 152, 114–126. https://doi.org/10.1016/j.cognition.2016.03.015 (2016).
Erb, C. D., Smith, K. A. & Moher, J. Tracking continuities in the flanker task: From continuous flow to movement trajectories. Atten. Percept. Psychophys. 83(2), 731–747. https://doi.org/10.3758/s13414-020-02154-4 (2021).
Freeman, J. B. Doing psychological science by hand. Curr. Dir. Psychol. Sci. 27(5), 315–323. https://doi.org/10.1177/0963721417746793 (2018).
Gallivan, J. P. & Chapman, C. S. Three-dimensional reach trajectories as a probe of real-time decision-making between multiple competing targets. Front. Neurosci. 8, 1–19. https://doi.org/10.3389/fnins.2014.00215 (2014).
Gallivan, J. P., Chapman, C. S., Wolpert, D. M. & Flanagan, J. R. Decision-making in sensorimotor control. Nat. Rev. Neurosci. 19(9), 519–534. https://doi.org/10.1038/s41583-018-0045-9 (2018).
Eriksen, B. A. & Eriksen, C. W. Effects of noise letters upon the identification of a target letter in a nonsearch task. Percept. Psychophys. 16(1), 143–149. https://doi.org/10.3758/bf03203267 (1974).
Erb, C. D. & Marcovitch, S. Deconstructing the Gratton effect: Targeting dissociable trial sequence effects in children, pre-adolescents, and adults. Cognition 179, 150–162. https://doi.org/10.1016/j.cognition.2018.06.007 (2018).
Erb, C. D., Touron, D. R. & Marcovitch, S. Tracking the dynamics of global and competitive inhibition in early and late adulthood: Evidence from the flanker task. Psychol. Aging 35(5), 729–743. https://doi.org/10.1037/pag0000435.supp (2020).
Gratton, G., Coles, M. G. & Donchin, E. Optimizing the use of information: Strategic control of activation of responses. J. Exp. Psychol. Gen. 121(4), 480–506. https://doi.org/10.1037/0096-3445.121.4.480 (1992).
Mayr, U., Awh, E. & Laurey, P. Conflict adaptation effects in the absence of executive control. Nat. Neurosci. 6(5), 450–452. https://doi.org/10.1038/nn1051 (2003).
Nieuwenhuis, S. et al. Accounting for sequential trial effects in the flanker task: Conflict adaptation or associative priming?. Mem. Cognit. 34(6), 1260–1272. https://doi.org/10.3758/bf03193270 (2006).
Aron, A. R., Robbins, T. W. & Poldrack, R. A. Inhibition and the right inferior frontal cortex: One decade on. Trends Cogn. Sci. 18(4), 177–185. https://doi.org/10.1016/j.tics.2013.12.003 (2014).
Cavanagh, J. F. et al. Subthalamic nucleus stimulation reverses mediofrontal influence over decision threshold. Nat. Neurosci. 14(11), 1462–1467. https://doi.org/10.1038/nn.2925 (2011).
Diesburg, D. A. & Wessel, J. R. The Pause-then-Cancel model of human action-stopping: Theoretical considerations and empirical evidence. Neurosci. Biobehav. Rev. 129, 17–34. https://doi.org/10.31234/osf.io/vp6es (2021).
Frank, M. J. Hold your horses: A dynamic computational role for the subthalamic nucleus in decision making. Neural Netw. 19(8), 1120–1136. https://doi.org/10.1016/j.neunet.2006.03.006 (2006).
Muralidharan, V., Aron, A. R. & Schmidt, R. Transient beta modulates decision thresholds during human action-stopping. Neuroimage 254, 119145. https://doi.org/10.1101/2021.07.05.447605 (2022).
Tatz, J. R., Soh, C. & Wessel, J. R. Common and unique inhibitory control signatures of action-stopping and attentional capture suggest that actions are stopped in two stages. J. Neurosci. 41(42), 8826–8838. https://doi.org/10.1523/jneurosci.1105-21.2021 (2021).
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S. & Cohen, J. D. Conflict monitoring and cognitive control. Psychol. Rev. 108(3), 624. https://doi.org/10.1037/0033-295x.108.3.624 (2001).
Shenhav, A., Botvinick, M. M. & Cohen, J. D. The expected value of control: An integrative theory of anterior cingulate cortex function. Neuron 79(2), 217–240. https://doi.org/10.1016/j.neuron.2013.07.007 (2013).
Erb, C. D. et al. Linking the dynamics of cognitive control to individual differences in working memory capacity: Evidence from reaching behavior. J. Exp. Psychol. Learn. Mem. Cogn. 47(9), 1383–1402. https://doi.org/10.1037/xlm0001018.supp (2021).
Erb, C. D., McBride, A. G. & Marcovitch, S. Associative priming and conflict differentially affect two processes underlying cognitive control: Evidence from reaching behavior. Psychon. Bull. Rev. 26(4), 1400–1410. https://doi.org/10.3758/s13423-019-01576-y (2019).
Erb, C. D., Moher, J., Song, J. H. & Sobel, D. M. Reach tracking reveals dissociable processes underlying inhibitory control in 5- to 10-year-olds and adults. Dev. Sci. 21(2), 114–126. https://doi.org/10.1111/desc.12523 (2018).
Erb, C. D. The developing mind in action: Measuring manual dynamics in childhood. J. Cogn. Dev. 19(3), 233–247. https://doi.org/10.1080/15248372.2018.1454449 (2018).
Erb, C. D. & Aschenbrenner, A. J. Multiple expectancies underlie the congruency sequence effect in confound-minimized tasks. Acta Psychol 198, 102869. https://doi.org/10.1016/j.actpsy.2019.102869 (2019).
Dotan, D., Pinheiro-Chagas, P., Al Roumi, F. & Dehaene, S. Track it to crack it: Dissecting processing stages with finger tracking. Trends Cogn. Sci. 23(12), 1058–1070. https://doi.org/10.1016/j.tics.2019.10.002 (2019).
Dshemuchadse, M., Scherbaum, S. & Goschke, T. How decisions emerge: action dynamics in intertemporal decision making. J. Exp. Psychol. Gen. 142(1), 93. https://doi.org/10.1037/a0028499 (2013).
Faulkenberry, T. J., Witte, M. & Hartmann, M. Tracking the continuous dynamics of numerical processing: A brief review and editorial. J. Numer. Cogn. 4(2), 271–285. https://doi.org/10.31234/osf.io/pruz7 (2018).
Freeman, J. B., Dale, R. & Farmer, T. A. Hand in motion reveals mind in motion. Front. Psychol. 2, 59–64. https://doi.org/10.3389/fpsyg.2011.00059 (2011).
Incera, S. & McLennan, C. T. Mouse tracking reveals that bilinguals behave like experts. Biling. Lang. Cogn. 19(3), 610–620. https://doi.org/10.1017/s1366728915000218 (2016).
Incera, S. & McLennan, C. T. Bilingualism and age are continuous variables that influence executive function. Neuropsychol. Dev. Cogn. B Aging Neuropsychol. Cogn. 25(3), 443–463. https://doi.org/10.1080/13825585.2017.1319902 (2018).
Ingram, J. N., Körding, K. P., Howard, I. S. & Wolpert, D. M. The statistics of natural hand movements. Exp. Brain Res. 188(2), 223–236. https://doi.org/10.1007/s00221-008-1355-3 (2008).
Miall, R. C. & Reckess, G. Z. The cerebellum and the timing of coordinated eye and hand tracking. Brain Cogn. 48(1), 212–226. https://doi.org/10.1006/brcg.2001.1314 (2002).
Resulaj, A., Kiani, R., Wolpert, D. M. & Shadlen, M. N. Changes of mind in decision-making. Nature 461(7261), 263–266. https://doi.org/10.1038/nature08275 (2009).
Scherbaum, S., Dshemuchadse, M., Fischer, R. & Goschke, T. How decisions evolve: The temporal dynamics of action selection. Cognition 115(3), 407–416. https://doi.org/10.1016/j.cognition.2010.02.004 (2010).
Scherbaum, S., Frisch, S. & Dshemuchadse, M. A bird in the hand isn’t good for long: Action dynamics reveal short-term choice impulses in intertemporal choices. Exp. Psychol. 65(1), 23–31. https://doi.org/10.1027/1618-3169/a000385 (2018).
Schoemann, M., O’Hora, D., Dale, R. & Scherbaum, S. Using mouse cursor tracking to investigate online cognition: Preserving methodological ingenuity while moving toward reproducible science. Psychon. Bull. Rev. 28(3), 766–787. https://doi.org/10.31234/osf.io/4ku26 (2021).
Song, J. H. Abandoning and modifying one action plan for alternatives. Philos. Trans. R. Soc. Lond. B Biol. Sci. 372(1718), 20160195. https://doi.org/10.1098/rstb.2016.0195 (2017).
Stillman, P. E., Medvedev, D. & Ferguson, M. J. Resisting temptation: Tracking how self-control conflicts are successfully resolved in real time. Psychol. Sci. 28(9), 1240–1258. https://doi.org/10.1177/0956797617705386 (2017).
Stillman, P. E., Shen, X. & Ferguson, M. J. How mouse-tracking can advance social cognitive theory. Trends Cogn. Sci. 22(6), 531–543. https://doi.org/10.1016/j.tics.2018.03.012 (2018).
Sullivan, N., Hutcherson, C., Harris, A. & Rangel, A. Dietary self-control is related to the speed with which attributes of healthfulness and tastiness are processed. Psychol. Sci. 26(2), 122–134. https://doi.org/10.1177/0956797614559543 (2015).
Calcagnì, A., Lombardi, L., D’Alessandro, M. & Freuli, F. A state space approach to dynamic modeling of mouse-tracking data. Front. Psychol. 10, 1–16. https://doi.org/10.3389/fpsyg.2019.02716 (2019).
Hehman, E., Stolier, R. M. & Freeman, J. B. Advanced mouse-tracking analytic techniques for enhancing psychological science. Group Process. Intergroup Relat. 18(3), 384–401. https://doi.org/10.1177/1368430214538325 (2015).
Kieslich, P. J., Schoemann, M., Grage, T., Hepp, J. & Scherbaum, S. Design factors in mouse-tracking: What makes a difference?. Behav. Res. 52(1), 317–341. https://doi.org/10.3758/s13428-019-01228-y (2020).
Wulff, D. U., Haslbeck, J. M., Kieslich, P. J., Henninger, F. & Schulte-Mecklenbeck, M. Mouse-tracking: Detecting types in movement trajectories. In A Handbook of Process Tracing Methods (eds Schulte-Mecklenbeck, M. et al.) 131–145 (Routledge, 2019). https://doi.org/10.31234/osf.io/6edca.
Scherbaum, S. & Kieslich, P. J. Stuck at the starting line: How the starting procedure influences mouse-tracking data. Behav. Res. Methods 50(5), 2097–2110. https://doi.org/10.3758/s13428-017-0977-4 (2018).
Ye, W. & Damian, M. F. Exploring task switch costs in a color-shape decision task via a mouse tracking paradigm. J. Exp. Psychol. Hum. Percept. Perform. 48(1), 8–20. https://doi.org/10.1037/xhp0000975 (2022).
Lane, A. E. & Ziviani, J. M. Factors influencing skilled use of the computer mouse by schoolaged children. Comput. Educ. J. 55, 1112–1122. https://doi.org/10.1016/j.compedu.2010.05.008 (2010).
Chevalier, N., Kelsey, K. M., Wiebe, S. A. & Espy, K. A. The temporal dynamic of response inhibition in early childhood: An ERP study of partial and successful inhibition. Dev. Neuropsychol. 39(8), 585–599. https://doi.org/10.1080/87565641.2014.973497 (2014).
Cragg, L., Fox, A., Nation, K., Reid, C. & Anderson, M. Neural correlates of successful and partial inhibitions in children: An ERP study. Dev. Psychobiol. 51(7), 533–543. https://doi.org/10.1002/dev.20391 (2009).
Cragg, L. & Nation, K. Go or no-go? Developmental improvements in the efficiency of response inhibition in mid-childhood. Dev. Sci. 11(6), 819–827. https://doi.org/10.1111/j.1467-7687.2008.00730.x (2008).
Lim, C. E. & Cho, Y. S. Response mode modulates the congruency sequence effect in spatial conflict tasks: Evidence from aimed-movement responses. Psychol. Res. https://doi.org/10.1007/s00426-020-01376-3 (2020).
Narang, V. & Laycock, J. R. D. Psychomotor testing of oncall anaesthetists. Anaesthesia 41(8), 868–869. https://doi.org/10.1111/j.1365-2044.1986.tb13136.x (1986).
Hommel, B. Event files: Feature binding in and across perception and action. Trends Cogn. Sci. 8(11), 494–500. https://doi.org/10.1016/j.tics.2004.08.007 (2004).
Hommel, B., Proctor, R. W. & Vu, K. P. L. A feature-integration account of sequential effects in the Simon task. Psychol. Res. 68(1), 1–17. https://doi.org/10.1007/s00426-003-0132-y (2004).
Moher, J. & Song, J. H. A comparison of simple movement behaviors across three different devices. Atten. Percept. Psychophys. 81(7), 2558–2569. https://doi.org/10.3758/s13414-019-01856-8 (2019).
Zelazo, P. D. et al. II. NIH toolbox cognition battery (CB): Measuring executive function and attention. Monogr. Soc. Res. Child Dev. 78(4), 16–33. https://doi.org/10.1111/mono.12032 (2013).
Burgoyne, A. P. & Engle, R. W. Attention control: A cornerstone of higher-order cognition. Curr. Dir. Psychol. Sci. 29(6), 624–630. https://doi.org/10.1177/0963721420969371 (2020).
Draheim, C., Mashburn, C. A., Martin, J. D. & Engle, R. W. Reaction time in differential and developmental research: A review and commentary on the problems and alternatives. Psychol. Bull. 145(5), 508–535. https://doi.org/10.1037/bul0000192 (2019).
Gratton, G., Coles, M. G., Sirevaag, E. J., Eriksen, C. W. & Donchin, E. Pre- and poststimulus activation of response channels: A psychophysiological analysis. J. Exp. Psychol. Hum. Percept. Perform 14(3), 331–344. https://doi.org/10.1037/0096-1523.14.3.331 (1988).
Tafuro, A., Vallesi, A. & Ambrosini, E. Cognitive brakes in interference resolution: A mouse-tracking and EEG co-registration study. Cortex 133, 188–200. https://doi.org/10.1016/j.cortex.2020.09.024 (2020).
Holroyd, C. B., Dien, J. & Coles, M. G. Error-related scalp potentials elicited by hand and foot movements: Evidence for an output-independent error-processing system in humans. Neurosci. Lett. 242(2), 65–68. https://doi.org/10.1016/s0304-3940(98)00035-4 (1998).
Ridderinkhof, K. R. & van der Molen, M. W. When global information and local information collide: A brain potential analysis of the locus of interference effects. Biol. Psychol. 41(1), 29–53. https://doi.org/10.1016/0301-0511(95)05125-t (1995).
Simon, J. R. The effects of an irrelevant directional cue on human information processing. In Advances in Psychology Vol 65 (eds Proctor, R. W. & Reeve, T. G.) 31–86 (North-Holland, 1990). https://doi.org/10.1016/s0166-4115(08)61218-2.
Holroyd, C. B., Yeung, N., Coles, M. G. & Cohen, J. D. A mechanism for error detection in speeded response time tasks. J. Exp. Psychol. Gen. 134(2), 163–191. https://doi.org/10.1037/0096-3445.134.2.163 (2005).
Scharinger, C., Soutschek, A., Schubert, T. & Gerjets, P. When flanker meets the n-back: What EEG and pupil dilation data reveal about the interplay between the two central-executive working memory functions inhibition and updating. Psychophysiology 52(10), 1293–1304. https://doi.org/10.1111/psyp.12500 (2015).
Funding
This article was funded by University of Auckland (Grant no. Performance-Based Research Fund).
Author information
Authors and Affiliations
Contributions
K.A.S. and C.D.E. generated the research question. All authors contributed to the preregistration and project development. S.M. programmed the task, designed, and built the response box. K.A.S. and S.M. collected the data under the supervision of A.M.E.H. and C.D.E. K.A.S. and C.D.E. conducted the data analysis and wrote the majority of the manuscript, with critical revisions provided by S.M. and A.M.E.H. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Smith, K.A., Morrison, S., Henderson, A.M.E. et al. Moving beyond response times with accessible measures of manual dynamics. Sci Rep 12, 19065 (2022). https://doi.org/10.1038/s41598-022-20579-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-20579-9
- Springer Nature Limited