
Abstract
Analyses characterizing canopy gaps are required to improve our understanding of spatial and structural variations in forest canopies and provide insight into ecosystem-level successional processes. Gap size frequency distributions (GSFD) are indicative of ecological processes and disturbance patterns. To date, GSFD in boreal forest ecosystems have not been systematically quantified over large areas using a single consistent data source. Herein we characterized GSFDs across the entirety of the Canadian boreal forest using transects of airborne laser scanning (ALS) data. ALS transects were representatively sampled within eight distinct Canadian boreal ecozones. Gaps were detected and delineated from the ALS-derived canopy height model as contiguous canopy openings ≥8 m2 with canopy heights ≤3 m. Gaps were then stratified by ecozone and forest type (i.e. coniferous, broadleaf, mixedwood, wetland-treed), and combinations thereof, and GSFDs were calculated for each stratum. GSFDs were characterized by the scaling parameter of the power-law probability distribution, lambda (λ) and Kolmogorov-Smirnov tests confirmed that GSFDs for each stratum followed a power-law distribution. Pairwise comparisons between ecozones, forest types, and combinations thereof indicated significant differences between estimates of λ. Scaling parameters were found to be more variable by ecozone (1.96–2.31) than by forest type (2.15–2.21). These results contrast those of similar studies done in tropical forest environments, whereby λ was found to be relatively consistent across a range of site types, geological substrates, and forest types. The geographic range considered herein is much larger than that of previous studies, and broad-scale patterns in climate, landforms, and soils that are reflected in the definition of unique ecozones, likely also influence gap characteristics.
Similar content being viewed by others


Introduction
Data characterizing the state and variability of large- and fine-scale forest disturbances is critical for evaluating forest dynamics and influencing current and future management strategies1. The need for these data is especially pertinent in extensively forested nations such as Canada, where boreal forests cover more than 270 Mha2,3, are predominantly remote, and are subject to seasonably variable and harsh climates4. Limited access to much of the Canadian boreal has constrained management intervention and industrial development5, leading to reliance on remotely sensed information to monitor, study, and generate synoptic data products for these ecosystems6. Generated using satellite based time-series, high-quality spatially explicit data detailing land cover7, disturbance8,9, and recovery1,10 have provided valuable insight into boreal forest dynamics and trends. These remote sensing based analyses serve to inform the ecological importance of biotic and abiotic disturbances, highlighting the implications of forest structural and physiological change at local to regional scales11,12.
Within the context of regional satellite based remote sensing studies, an abundance of research has focused on the prevalence and influence of spatially extensive abiotic and biotic disturbances such as stand replacing wildfires and non-stand replacing insect outbreaks6,7,10,13,14,15,16. Over boreal ecosystems, wildfire is recognized as a dominant driver for the spatial and structural conditions present, and have been shown to influence carbon and water dynamics17,18,19, create shifts in competitive wildlife use20,21, and in the long-term, rejuvenate ecosystem function through successional change22,23,24. Given the size of the Canadian boreal, substantial variability in fire regimes exist8, leading to differences in the manifestation and functional effect of fire events25.
In the absence of fire, other non-stand replacing disturbances are prevalent. Biotic disturbances including insect and fungal outbreaks, as well as abiotic events such as storm related wind throw and drought stress, result in widespread and important successional changes to forested ecosystems11,26,27. The role of these non-fire disturbances has been largely underemphasized, particularly in regards to their creation of canopy gaps and gap characteristics28. Canopy gaps are influential drivers of forest development26,28 and have been found to be highly variable in size29. Herein we refer to gaps as openings in the forest canopy originating from natural causes such as tree mortality, wind throw events, and branch fall30. Observed synchrony in the timing of gap formation suggests that landscape level events control gap dynamics31,32, with recent research indicating that their functional presence may not manifest prior to a few decades of development in boreal stands33.
Studies analyzing canopy gaps have largely been defined by a trade-off between the cost of data acquisition and the level of spatial and attributional detail for their delineation34. Until recently, canopy gaps were characterized via costly, time consuming, manual methodologies35,36,37, generally limiting extent and representation of research activity. The development of airborne laser scanning (ALS) and other remote sensing approaches has notably reduced barriers to characterizing canopy gaps over a range of forest environments33,34,36,38,39,40,41,42,43,44. The development of ALS has facilitated delineation of canopy gaps and their prevalence across forested landscapes41. Delineation of gaps is a straight-forward process, where minimum canopy height thresholds are established and used to delineate openings in ALS generated canopy height models (CHM)34. Following their delineation, gap attributes such as area, shape, and density are often derived. Vepakomma et al.45,46 for example investigated the potential to use multi-temporal ALS datasets in the eastern Canadian boreal forest to examine changes in gap size and shape over time. White et al.34 likewise tested the capacity of ALS to delineate canopy gaps in a coastal temperate rainforest in western Canada, outlining its enhanced accuracy in comparison to co-occurring digital aerial photogrammetric datasets.
A method for summarizing landscape level characteristics of canopy gaps using ALS delineated gaps is the calculation of gap size frequency distributions (GSFD), which describe the relative frequency of gaps across a range of gap size classes35. GSFDs are often found to follow a power-law distribution, wherein the negative slope resulting from plotting gap frequency against gap size classes on a log-log scale is characterized by lambda (λ), the scaling exponent of the power-law distribution38,47. In these studies λ has been used to compare GSFDs for areas with different geological substrates and forest types41,48. Values of λ have been theorized to have a range from 1.0–3.0 in tropical forests38,42,48,49,50, with a threshold of 2.0 being indicative of whether a forest is dominated by larger canopy gaps resulting from spatially extensive, stand-initiating disturbances (λ < 2.0), or smaller gaps, which may be indicative of low mortality, high growth dynamics (λ > 2.0) and small disturbances38.
To address the paucity of research focusing on boreal canopy gap characteristics and facilitate inter-comparisons of GSFDs from global forest types, we detected and delineated canopy gaps, and quantified and compared GSFDs within eight distinct ecological units (ecozones) and four forest types (coniferous, deciduous, mixedwood, wetland treed) in the boreal forest of Canada. To offer extensive spatial representation as well as required local forest structural detail, a 25,000 km network of ALS transect data were used, enabling a spatially explicit and consistent means of delineating canopy gaps, and consequently GSFDs across Canadian boreal regions.
The aims of this study were to investigate whether boreal GSFDs followed power-law distributions, and whether the relationship between gap size and frequency differed amongst ecozones and forest types. In order to address these aims we posed the following questions:
Do gaps in boreal ecozones and forest types follow a power-law distribution?
Gap size frequency analyses within various tropical forest environments have found that power-law distributions are common41,42,48. As no similar broad-scale studies have been conducted within boreal forest environments, we had no a priori expectations as to the form of gap size frequency distributions.
Do scaling exponents for power-law distributions (λ) differ amongst boreal ecozones and forest types?
The λ scaling exponent describes the relationship between gap size and frequency for power-law distributions. Previous studies in tropical forests have found minimal variation in scaling exponents across different sites, geological substrates, and forest types41,48. We therefore expected that λ values would not differ significantly among ecozones or forest types, confirming the consistency of scaling exponents across variable forest conditions.
In addition to these overarching research questions and hypotheses, we present descriptive statistics on gap size, shape, and density by ecozones and forest types. Finally, we considered GSFD for boreal forests with reference to values reported in the literature representing other forested regions with an aim to better understand similarities and differences in GSFDs and aid in the interpretation of our results in a broader context.
Materials and methods
Study area
The Canadian boreal forest is characterized by short cool summers, long cold winters, and is subject to variable temperature ranges3. Variations in climate as well as biotic and abiotic factors have led to the delineation of 6 boreal terrestrial ecozones51, the largest of which, the Taiga and Boreal Shield, are often divided into eastern and western sub-units to better reflect their eco-climatic conditions1,10. Proportions of managed land within ecozones vary, with Taiga Plains, Taiga Shield East and West, and Hudson Plains having <20%, Boreal Shield East and West <70%, and Boreal Plains <90% area considered to be managed1.
Airborne laser scanning data
Small-footprint, discrete return ALS data is an exhaustively researched technology with proven potential for characterizing forest structure and improving forest inventory information at multiple scales52,53. To leverage the benefits of this technology for canopy gap delineation, ALS data were acquired in the summer of 2010 by the Canadian Forest Service in collaboration with the Canadian Consortium for LiDAR Environmental Applications Research and the Applied Geomatics Research Group. Thirty-four transects averaging 700 km in length (~25,000 km in total) were acquired over all Canadian boreal forest ecozones (Fig. 1). Acquisition parameters for ALS transects are detailed in Table 1.

Study area map of Canadian boreal ecozones (Boreal Cordillera (BC), Taiga Plain (TP), Boreal Plain (BP), Taiga Shield West/East (TSW/E), Hudson Plain, and Boreal Shield West/East (BSW/E) (green), ALS transects acquired in 2010 (orange), and representative sections of transect used for canopy gap delineation (red).
ALS transects were processed using the LAStools54 programming suite, which enables fast and efficient reading, writing, and manipulation of ALS point clouds. In order to compute CHMs and canopy cover (% of ALS points > 2 m) layers for all ALS transects, point clouds were first divided into 1 km2 tiles using lastile and filtered to remove noise using lasnoise. Points were then classified as ground or non-ground points using lasground to extract a digital terrain model, and height normalized using lasheight. Normalized point clouds were used to generate CHMs at a 2 m resolution and canopy cover at 20 m resolution using lascanopy.
Selection of representative ALS transect sections
To ensure that areas of analysis encompassed as much variability in vegetation as possible, the MODIS MOD17A3 (https://lpdaac.usgs.gov/products/mod17a3v055/) gross primary productivity (GPP) product was used to delineate samples of ALS transects that were most representative of corresponding ecozones. MODIS derived GPP are well established to provide reliable vegetation productivity estimates55. Given that fluctuations in temperature and vegetation condition influence productivity estimates, individual years may not be representative of long-term productivity. To account for these variations, annual GPP estimates from 2005–2010 were averaged to align productivity with estimates of vegetation structure. Delineation of representative ALS transect sections was conducted using a probability distribution function where 30% of averaged MODIS GPP pixels intersecting ALS transects were randomly sampled with replacement over 10,000 iterations. Kolmogorov-Smirnov tests were then used to assess whether significant (a = 0.05) differences between bootstrapped GPP sample distributions and distributions of GPP for each ecozone existed. Transect sections maximizing the fit between ecozone GPP distributions and the sample distribution were considered representative of the ecozone and were selected for further processing. Representative transect sections used in this study are open data and include height percentiles, canopy cover, and coefficient of variation of height56.
Canopy gap delineation
To isolate forested areas within representative ALS transect sections, only areas meeting the definition for forest used by the Food and Agriculture Organization (FAO) of the United Nations were used in our analysis57 (i.e. areas ≥ 0.5 ha, canopy cover ≥ 10%, and height ≥ 5 m). CHMs and canopy cover (% of ALS pts > 2 m) layers were used to delineate areas within transects meeting threshold specifications.
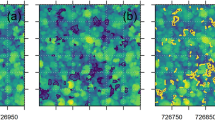
Following delineation of forested areas within representative ALS transects, a height threshold of 3 m40 was applied to CHMs to differentiate between gap and no-gap pixels. All pixels with heights ≤ 3 m were identified as gaps (Fig. 2). Connected gap pixels were considered single contiguous gaps. A maximum canopy gap size was not defined as has been presented in previous literature34 in order to include all gaps within delineated forested areas regardless of their area coverage and complexity of shape. Following delineation, each gap was attributed a respective ecozone, as well as assigned a forest type (coniferous, broadleaf, mixedwood, or wetland-treed) based on the disturbance informed Virtual Land Cover Engine (VLCE), a Landsat derived, spatially extensive, temporally dense, and flexible framework for mapping land cover7.

Graphical description of applied canopy gap delineation and gap size frequency distribution (GSFD) methodology.
In order to characterize properties of detected gaps, a number of spatial characteristics including gap area, shape index, and distance to nearest gap were computed. Shape index to characterize gap shape complexity was calculated using Eq. 1.
A shape index of 1 describes a perfectly circular gap. As shape index increases, the gap boundary becomes increasingly complex. Distance to nearest gap indicates gap density and was calculated as the distance between the bounding boxes of a gap and its closest neighbour.
Gap size frequency distributions
Gap size frequency distributions for all boreal ecozones and forest types were calculated using the approach of Hanel et al.58, which employs Bayesian parameter estimation utilizing maximum likelihood estimators that calculate λ across the entire range of reasonably accessible values59,60. This method also integrates a two-sided Kolmogorov-Smirnov goodness of fit test to determine whether GSFDs follow a power-law distribution. A key parameter when calculating GSFDs is the minimum included gap size. In this analysis we tested the impact of minimum gap size by including gaps ranging from 4 m2 (i.e. a single 2 m CHM cell) to a maximum of 80 m2. The number of samples included in each bootstrap (500, 1000, 2000, 5000, and 10000) was also assessed.
A smallest gap size of 8 m2 with 5000 samples per bootstrap iteration was chosen due to consistency between bootstrapped λ estimates. These parameters were used for quantifying GSFDs for ecozones, forest type, and their combinations. Estimates of λ were not generated where the minimum number of gap samples was not met.
Bootstrapped estimates of λ provided distributions that facilitated the two-sided Kolmogrov-Smirnov test for each ecozone, forest type, and ecozone/forest type combination to determine whether distributions conform to that of a power-law. Kruskal-Wallis and Dunn’s post-hoc tests were applied to assess whether ecozones, forest type, and their pairings showed significantly (a = 0.05) different λ estimates.
Results
Delineated gap features
A total of 1,037,358 gaps were included with a mean gap area of 57.7 m2. Hudson Plain and Taiga Shield East ecozones had the largest median gap sizes relative to other ecozones (Fig. 3a). Taiga Shield East had the largest inter-quartile range for gap area and shape index (Fig. 3b). Distance to nearest gap was greatest for deciduous stands, however was relatively consistent across forest types with a coefficient of variation of 38.3% (Fig. 3c). Wetland-treed gaps were found to have a higher median gap area and larger interquartile range for shape index (Fig. 3d,e). Coniferous gaps were found to have the smallest inter-quartile range for gap size. Deciduous gaps had the largest median, and greatest variability, in distance to nearest gap (Fig. 3f).

Summaries of gap area (a,d) (m2), shape index (b,e), and distance to nearest gap (c,f) (m) for ecozones (Boreal Cordillera (BC), Boreal Plains (BP), Boreal Shield East (BSE), Boreal Shield West (BSW), Hudson Plain (HP), Taiga Plain (TP), Taiga Shield East (TSE), and Taiga Shield West (TSW)) and forest types (Coniferous (C), Deciduous (D), Mixedwood (M), and Wetland-treed (WT)).
Gap size frequency distributions
Two-sided Kolmogorov-Smirnov tests indicated that the GSFDs for all ecozones, forest types and combinations thereof conformed to a power-law distribution (Fig. 4). Mean λ for all ecozones was 2.18, with the Boreal Plains having the highest (λ = 2.31) and Taiga Shield East having the lowest (λ = 1.96). All ecozone λ estimates were greater than 2.0, except Taiga Shield East. Mean λ for forest types were highest for conifers (λ = 2.21) and lowest for wetland-treed (λ = 2.15; Table 2). All ecozones paired with coniferous forest types had sufficient gap samples to conduct bootstrapping. Taiga Plains, Boreal Plains, and Boreal Shield East ecozones were the only ecozones with sufficient gap samples to calculate λ for all ecozone-forest type combinations (Table 2).

Gap size frequency distributions (GSFD) in eight boreal ecozones and four forest types. Dark grey markers represent the GSFD for each ecozone/forest type, while light grey markers represent GSFD for all ecozones/forest types combined. Axes are logarithmic. Kolmogorov-Smirnov test statistics (KS), critical test statistics (KScrit), and λ are the mean values from iterative bootstrapping.
Kruskall-Wallis and Dunn’s post-hoc tests were conducted for ecozones, forest types, and combinations thereof. All ecozones were found to have significantly different λ distributions (Fig. 5a). Forest types except conifer and deciduous had significantly different distributions of λ (Fig. 5b).

Comparison of bootstrapped distributions of λ for (a) ecozone (Boreal Cordillera (BC), Boreal Plains (BP), Boreal Shield East (BSE), Boreal Shield West (BSW), Hudson Plain (HP), Taiga Plain (TP), Taiga Shield East (TSE), and Taiga Shield West (TSW)), and (b) forest type (Coniferous (C), Deciduous (D), Mixedwood (M), and Wetland-treed (WT). Letters above boxplots indicate Dunn’s post-hoc test results denoting whether distributions were significantly different (a = 0.05).
Combinations of ecozone and forest type showed a range of λ results (Fig. 6). Gaps within coniferous and mixedwood forests in the Taiga Shield East ecozone were found to have the two lowest overall λ values. These two pairings were the only distributions with λ values below 2.0. Distributions for coniferous Hudson Plains, wetland-treed Boreal Shield West, and coniferous Boreal Cordillera shared similar distributions. Coniferous Taiga Shield West and both deciduous and mixedwood Boreal Plains shared similar distributions of λ. The coniferous Boreal Plains pairing was found to have the highest estimated λ distribution.

Comparison of bootstrapped distributions of λ for ecozone-forest type pairings. Letters above each boxplot indicate Dunn’s post-hoc test results denoting whether distributions were significantly different (a = 0.05) from one another. Boreal Cordillera (BC), Boreal Plains (BP), Boreal Shield East (BSE), Boreal Shield West (BSW), Hudson Plain (HP), Taiga Plain (TP), Taiga Shield East (TSE), and Taiga Shield West (TSW).
Discussion
In this study, we delineated canopy gaps within representative sections of ALS transects to compute GSFDs across a range of forest conditions in Canadian boreal forest ecozones. We found that the number and size of gaps, as well as GSFDs were different for ecozones, forest types, and their pairing. Our results represent the first spatially extensive quantification of GSFDs within a boreal forest context, complementing that of existing studies primarily focused on tropical forest ecosystems. The results presented herein can provide insight for investigations of boreal forest dynamics and improve understanding of similarities and differences amongst global forest systems. Comparisons between boreal and tropical GSFDs, with reference to ecological foundations and gap formation theory, can inform our understanding of forest dynamics in these different regions.
The Canadian boreal, though variable by region, is dominated by stand replacing wildfires1 with highly variable patch sizes61. Fires create landscape-level heterogeneity of forest structure, which are subject to smaller scale insect attacks or individual/group mortality events in their absence. Tropical forests such as those found in the Amazon region have markedly different climatic conditions, species composition, structure, and disturbance regimes from boreal forests. Espírito-Santo et al.62 estimated that small disturbances (<0.1 ha) are much more probable than large events (≥5 ha) in the Amazon region. Low probabilities of large disturbances in these forests contrast markedly to Canadian boreal ecozones where historically, wildfire has burned an average of 1.5 Mha annually (based on data representing 1985–2010)1. These spatially extensive disturbances were excluded from this analysis. Patches impacted by wildfire in Canada’s boreal are commonly 2–5 km2 in size (with some exceeding 1000 km2)8. Any recent wildfires (i.e. since 1985) within the bounds of ALS transects that did not meet the required thresholds for forest height, canopy cover, and minimum size were not considered in our analysis. With this context in mind, we discuss results and outcomes of this study with reference to our initial research questions and hypotheses.
Do gaps in boreal ecozones and forest types follow a power-law distribution?
Similar to results reported across a range of conditions in tropical forests, we found that gap size frequency in boreal forests followed a power-law distribution. Identifying power-law distributions for ecological features such as canopy gaps provides insight into the nature of gap formation processes such as tree mortality and local stand level disturbance. Changes in gap size were found to result in a similarly proportioned change in gap frequency, and that small canopy gaps were more prevalent than large canopy gaps within boreal forest ecozones and forest types.
Do scaling exponents for power-law distributions (λ) differ amongst boreal ecozones and forest types?
Our results confirmed that λ varied more amongst ecozones than forest types. Ecozones were all found to exhibit significant differences in λ, and had greater variability in scaling exponents. Forest types were found to exhibit less variability in λ, and significant differences in λ were not found between coniferous and deciduous forest types.
A commonly implemented heuristic in tropical forests is that GSFDs with λ values below 2.0 are dominated by larger gaps, created through spatially extensive disturbances, whereas GSFDs with λ values above 2.0 are dominated by smaller gaps and localized disturbance processes38,41,47. Tropical studies have reported λ values ranging from 1.1–1.638 and 1.9–3.147 in Amazonian forests, and 1.66–1.9963 and 1.8–2.648 in Costa Rican and Hawaiian forests respectively. Asner et al.41, likewise computed GSFDs in the Peruvian Amazon for a range of canopy depth classes (≥1 and ≥20 m), geologic, topographic, and physiognomic descriptors with largely consistent findings of λ values between 1.70–2.03. Within the context of these previous studies in tropical forest environments, results for λ presented herein for the Canadian boreal are 1.96–2.31 for ecozones and 2.15–2.21 for forest types. Scaling parameters were found to be relatively consistent across varying conditions in tropical forests41,48. However, we found significant differences in λ values among ecozones and some forest types, and greater variation among ecozones than forest types. Herein, the geographical range across which we evaluated gap size frequency distributions was much broader than that of past studies, suggesting that variations in climate, landforms, and soil types among others, which are inherent to the definition of Canada’s terrestrial ecozones, also influence gap characteristics.
Gap formation theory helps to support similarities among forest type GSFDs. In the event of tree mortality and windthrow, larger, higher biomass trees are more likely to result in larger canopy gaps64,65, and corresponding lower λ values. Boreal forests are predominantly short, have low diversity, and are comprised of regular crown shapes66. Short growing seasons in boreal forests reduce annual growth potential, generally resulting in smaller trees67. Reasonably similar estimates of scaling parameters among boreal forest types, especially conifer, deciduous, and mixedwood stands indicate similarities in the distribution of canopy gap sizes. Of all forest types considered, wetland-treed forests had the lowest values of λ and therefore greatest prevalence of larger canopy gaps. Larger proportions of these gaps are likely driven by prevalence of surface water, high soil moisture, and low nutrient edaphic conditions common to wet-land forest environments. Values of λ below 2.0 were also found for conifer and mixed-wood forests in the Taiga Shield East ecozone. This region is known for its harsh long winters, abundance of water features, and shallow wet soils, potentially leading to more spatially scattered, lower density, forest stands. Larger median gap sizes and shape indices for both the Taiga Shield East ecozone and wetland-treed forest type further reinforce these findings.
In the tropics, relative consistency in GSFDs has been attributed to light availability. Studies have modeled how forest canopy gaps develop in tropical forests using cellular automata theory68,69, which addresses how complex systems approaching a critical state will self-organize into similar spatial patterns. To this end, tropical forest species have been found to exhibit opportunistic and plastic growth patterns forming intricate and irregular tessellations70. These growth patterns result in tree canopies filling most of the tri-dimensional canopy space, which explains the gap-scaling processes across landscapes within a particular region41,70. While there is still debate surrounding the high degree of scaling consistency in forest ecosystems, resource limitation such as light has been thought to be the cause of this homogeneity in tropical forests41.
Tropical and temperate forests experience drastic differences in light levels between gap and non-gap areas. In boreal forests, gap and non-gap areas experience similar light distributions likely due to the presence of small gaps that are fairly evenly spaced71. Power-law distributions for boreal GSFDs and distance to nearest gap metrics presented herein help to confirm this. Research in boreal and temperate forests found spatial patterns to be clustered in young stands and more regular in older aged stands72,73. In these forests, competition between tree species drives forest patterns away from spatial clustering towards regularity74,75. Trees are opportunistic in their growth and will fill available canopy space76. This crown plasticity is particularly pronounced in mixed-species forests77. In boreal forests, we posit that this opportunistic growth is the primary reason for consistency in scaling parameters among forest types.
Conclusions
Findings from this study provide a useful baseline that quantifies the variability in GSFDs in the Canadian boreal forest. The relationship between gap size and frequency for all ecozones and forest types were found to fit a power-law distribution. Scaling parameters varied more by ecozone than by forest type, suggesting that broad-scale patterns in climate, landforms, and soils that are inherent to the definition of ecozones, also influence gap characteristics. Findings herein facilitate a comprehensive understanding and inter-comparison among global forest types that can help form the basis of additional hypotheses with respect to canopy gap size and frequency across space and time. Quantification of regional differences in boreal GSFDs provide insights regarding variations in ecological processes, including the manifestation of gap phase dynamics, as well as informing forest management practices that seek to emulate natural disturbance patterns.
References
White, J. C., Wulder, M. A., Hermosilla, T., Coops, N. C. & Hobart, G. W. A nationwide annual characterization of 25 years of forest disturbance and recovery for Canada using Landsat time series. Remote Sens. Environ. 194, 303–321 (2017).
Brandt, J. P. The extent of the North American boreal zone. 161, 101–161 (2009).
Brandt, J. P., Flannigan, M. D., Maynard, D. G., Thompson, I. D. & Volney, W. J. A. An introduction to Canada’s boreal zone: ecosystem processes, health, sustainability, and environmental issues. Environ. Rev. 21, 207–226 (2013).
Sulla-Menashe, D., Woodcock, C. E. & Friedl, M. A. Canadian boreal forest greening and browning trends: An analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environ. Res. Lett. 13 (2018).
Andrew, M. E., Wulder, M. A. & Coops, N. C. Identification of de facto protected areas in boreal Canada. Biol. Conserv. 146, 97–107 (2012).
Hermosilla, T. et al. Mass data processing of time series Landsat imagery: pixels to data products for forest monitoring. Int. J. Digit. Earth 8947, 1–20 (2016).
Hermosilla, T., Wulder, M. A., White, J. C., Coops, N. C. & Hobart, G. W. Disturbance-Informed Annual Land Cover Classification Maps of Canada’s Forested Ecosystems for a 29-Year Landsat Time Series. Can. J. Remote Sens. 44, 67–87 (2018).
Coops, N. C., Hermosilla, T., Wulder, M. A., White, J. C. & Bolton, D. K. A thirty year, fine-scale, characterization of area burned in Canadian forests shows evidence of regionally increasing trends in the last decade. PLoS One 13, 1–19 (2018).
Pickell, P. D., Coops, N. C., Gergel, S. E., Andison, D. W. & Marshall, L. Evolution of Canada’ s Boreal Forest Spatial Patterns as Seen from Space. 1–20, https://doi.org/10.1371/journal.pone.0157736 (2016).
Pickell, P. D., Hermosilla, T., Frazier, R. J., Coops, N. C. & Wulder, M. A. Forest recovery trends derived from Landsat time series for North American boreal forests Forest recovery trends derived from Landsat time series for North American boreal forests. Int. J. Remote Sens. 1161, 138–149 (2016).
Kneeshaw, D., Bergeron, Y. & Kuuluvainen, T. Forest ecosystem structure and disturbance dynamics across the circumboreal forest. (The Sage Handbook of Biogeography, 2011).
Thom, D. & Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 91, 760–781 (2016).
Hermosilla, T., Wulder, M. A., White, J. C., Coops, N. C. & Hobart, G. W. Regional detection, characterization, and attribution of annual forest change from 1984 to 2012 using Landsat-derived time-series metrics. Remote Sens. Environ. 170, 121–132 (2015).
Hermosilla, T., Wulder, M. A., White, J. C., Coops, N. C. & Hobart, G. W. An integrated Landsat time series protocol for change detection and generation of annual gap-free surface reflectance composites. Remote Sens. Environ. 158, 220–234 (2015).
Wolter, P. T., Townsend, P. A., Sturtevant, B. R. & Kingdon, C. C. Remote sensing of the distribution and abundance of host species for spruce budworm in Northern Minnesota and Ontario. Remote Sens. Environ. 112, 3971–3982 (2008).
Franklin, S. E., Fan, H. & Guo, X. Relationship between landsat TM and SPOT vegetation indices and cumulative spruce budworm defoliation. Int. J. Remote Sens. 29, 1215–1220 (2008).
Balshi, M. S. et al. The role of historical fire disturbance in the carbon dynamics of the pan-boreal region: A process-based analysis. J. Geophys. Res. Biogeosciences 112, 1–18 (2007).
Bond-Lamberty, B., Peckham, S. D., Ahl, D. E. & Gower, S. T. Fire as the dominant driver of central Canadian boreal forest carbon balance. Nature 450, 89–92 (2007).
Mkhabela, M. S. et al. Comparison of carbon dynamics and water use efficiency following fire and harvesting in Canadian boreal forests. Agric. For. Meteorol. 149, 783–794 (2009).
Dalerum, F., Boutin, S. & Dunford, J. S. Wildfire effects on home range size and fidelity of boreal caribou in Alberta, Canada. Can. J. Zool. 85, 26–32 (2007).
Hobson, K. A. & Schieck, J. Changes in Bird Communities in Boreal Mixedwood Forest: Harvest and Wildfire Effects over 30 Years. Ecol. Appl. 9, 849–863 (2012).
Pyne, S. The Ecology of Fire. Nat. Educ. Knowl. 3 (2010).
Bowman, D. M. J. S. et al. Fire in the Earth System. Science (80-.). 481, 481–484 (2009).
Bond, W. J. & Keeley, J. E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 20, 387–394 (2005).
de Groot, W. J. et al. A comparison of Canadian and Russian boreal forest fire regimes. For. Ecol. Manage. 294, 23–34 (2013).
Mccarthy, J. W. & Weetman, G. Age and size structure of gap-dynamic, old-growth boreal stands in Newfoundland. Silva Fenn. 40 (2006).
Whitmore, T. C. Canopy Gaps and the Two Major Groups of Forest Trees. Ecology 70, 536–538 (1989).
McCarthy, J. Gap dynamics of forest trees: A review with particular attention to boreal forests. Environ. Rev. 59, 1–59 (2001).
Pham, A. T., Grandpré, L., De, Gauthier, S. & Bergeron, Y. Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec. Can. J. For. Res. 34, 353–364 (2004).
Bonnet, S., Gaulton, R., Lehaire, F. & Lejeune, P. Canopy gap mapping from airborne laser scanning: An assessment of the positional and geometrical accuracy. Remote Sens. 7, 11267–11294 (2015).
Périgon, S. Dynamique de trouées dans de vieux peuplements résineux de la Côte-Nord, Québec. (2006).
Gauthier, S. et al. Ecosystem management in the boreal forest. (Presses de l’Université du Québec, 2009).
Senécal, J. & Doyon, F. C. M.-. Management implications of varying gap detection height thresholds and other canopy dynamics processes in temperate deciduous forests. For. Ecol. Manage. 410, 84–94 (2018).
White, J. C., Tompalski, P., Coops, N. C. & Wulder, M. A. Comparison of airborne laser scanning and digital stereo imagery for characterizing forest canopy gaps in coastal temperate rainforests. Remote Sens. Environ. 208, 1–14 (2018).
Runkle, J. R. Patterns of Disturbance in Some Old-Growth Mesic Forests of Eastern North America. Ecology 63, 1533–1546 (1982).
Vepakomma, U., St-Onge, B. & Kneeshaw, D. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Environ. 112, 2326–2340 (2008).
Battles, J. J., Fahey, T. J. & Harney, E. M. B. Spatial patterning in the canopy gap regime of a subalpine Abies-Picea forest in the northeastern United States. J. Veg. Sci. 6, 807–814 (2006).
Fisher, J. I., Hurtt, G. C., Thomas, R. Q. & Chambers, J. Q. Clustered disturbances lead to bias in large-scale estimates based on forest sample plots. Ecol. Lett. 11, 554–563 (2008).
Vehmas, M., Packalén, P., Maltamo, M. & Eerikäinen, K. Using airborne laser scanning data for detecting canopy gaps and their understory type in mature boreal forest. Ann. For. Sci. 68, 825–833 (2011).
Kane, V. R. et al. Patch dynamics and the development of structural and spatial heterogeneity in Pacific Northwest forests. Can. J. For. Res. 41, 2276–2291 (2011).
Asner, G. P. et al. Forest Canopy Gap Distributions in the Southern Peruvian Amazon. PLoS One 8 (2013).
Goulamoussène, Y., Bedeau, C., Descroix, L., Linguet, L. & Hérault, B. Environmental control of natural gap size distribution in tropical forests. Biogeosciences 14, 353–364 (2017).
D’Aoust, V., Kneeshaw, D. & Bergeron, Y. Characterization of canopy openness before and after a spruce budworm outbreak in the southern boreal forest. Can. J. For. Res. 34, 339–352 (2004).
Foody, G. M., Jackson, R. G. & Quine, C. P. Potential improvements in the characterization of forest canopy gaps caused b… Agriculture 49 (2003).
Vepakomma, U., St-Onge, B. & Kneeshaw, D. Response of a boreal forest to canopy gap openings – assessing vertical and horizontal tree growth with multi-temporal lidar data. Ecol. Appl. 21, 99–121 (2011).
Vepakomma, U., Kneeshaw, D. & Fortin, M. J. Spatial contiguity and continuity of canopy gaps in mixed wood boreal forests: Persistence, expansion, shrinkage and displacement. J. Ecol. 100, 1257–1268 (2012).
Lloyd, J., Gloor, E. U. & Lewis, S. L. Are the dynamics of tropical forests dominated by large and rare disturbance events? Ecol. Lett. 12, 19–21 (2009).
Kellner, J. R. & Asner, G. P. Convergent structural responses of tropical forests to diverse disturbance regimes. Ecol. Lett. 12, 887–897 (2009).
Boyd, D. S., Hill, R. A., Hopkinson, C. & Baker, T. R. Landscape-scale forest disturbance regimes in southern Peruvian Amazonia. Ecol. Appl. 23, 1588–1602 (2013).
Lobo, E. & Dalling, J. W. Effects of topography, soil type and forest age on the frequency and size distribution of canopy gap disturbances in a tropical forest. Biogeosciences 10, 6769–6781 (2013).
Ecological Stratification Working Group. A National Ecological Framework for Canada. Environment doi:Cat. No. A42-65/1996E; ISBN 0-662-24107-X (1995).
Wulder, M. A., Bater, C., Coops, N. C., Hilker, T. & White, J. The role of LiDAR in sustainable forest management. For. Chron. 84, 807–826 (2008).
White, J. C. et al. A best practices guide for generating forest inventory attributes from airborne laser scanning data using an area-based approach. For. Chron. 89, 50 (2013).
Isenburg, M. LAStools. Retrieved from http://lastools.org (2019).
Phillips, L. B., Hansen, A. J. & Flather, C. H. Evaluating the species energy relationship with the newest measures of ecosystem energy: NDVI versus MODIS primary production. Remote Sens. Environ. 112, 4381–4392 (2008).
Wulder, M. A. et al. Lidar plots — a new large-area data collection option: Context, concepts, and case study. Can. J. Remote Sens. 38, 600–618 (2012).
Food and Agriculture Organization of the United Nations. The Forest Resources Assessment Programme: Terms and Defninitions (2012).
Hanel, R., Corominas-Murtra, B., Liu, B. & Thurner, S. Fitting power-laws in empirical data with estimators that work for all exponents. PLoS One 12, e0170920 (2017).
Bauke, H. Parameter estimation for power-law distributions by maximum likelihood methods. Eur. Phys. J. B 58, 167–173 (2007).
Clauset, A., Shalizi, C. R. & Newman, M. E. J. Power-law distributions in empirical data. 51, 661–703 (2007).
San-Miguel, I., Andison, D. W. & Coops, N. C. Characterizing historical fire patterns as a guide for harvesting planning using landscape metrics derived from long term satellite imagery. For. Ecol. Manage. 399, 155–165 (2017).
Espírito-Santo, F. D. B. et al. Size and frequency of natural forest disturbances and the Amazon forest carbon balance. Nat. Commun. 5, 1–6 (2014).
Kellner, J. R., Clark, D. B. & Hubbell, S. P. Pervasive canopy dynamics produce short-term stability in a tropical rain forest landscape. Ecol. Lett. 12, 155–164 (2009).
Oliver, C. D. & Larson, B. C. Forest stand dynamics: updated edition. Forest stand dynamics: updated edition (John Wiley and sons, 1996).
Brokaw, N. V. L. Gap-Phase Regeneration in a Tropical Forest. Ecology 66, 682–687 (1985).
Palace, M., Keller, M., Asner, G. P., Hagen, S. & Braswell, B. Amazon Forest Structure from IKONOS Satellite Data and the Automated Characterization of Forest Canopy Properties. Biotropica 40, 141–150 (2008).
Price, D. T. et al. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 21, 322–365 (2013).
Sole, R. V. & Manrubia, S. C. Are Rainforests Self-organized in a Cirtical State? J. theor Biol. 173, 31–40 (1995).
Bak, P. & Chen, K. Self-Organized Criticality. Sci. Am. 264, 46–53 (1991).
Herwitz, S. R., Slye, R. E. & Turton, S. M. Long-Term Survivorship and Crown Area Dynamics of Tropical Rain Forest Canopy Trees. Ecology 81, 585–597 (2000).
Bartemucci, P., Coates, K. D., Harper, K. A. & Wright, E. F. Gap disturbances in northern old-growth forests of British Columbia, Canada. J. Veg. Sci. 13, 685–696 (2002).
Moeur, M. Characterizing spatial patterns of trees using stem-mapped data. For. Sci. 39, 756–775 (1993).
Brassard, B. W. & Chen, H. Y. H. Stand Structural Dynamics of North American Boreal Forests. CRC. Crit. Rev. Plant Sci. 25, 115–137 (2006).
Szwagrzyk, J. & Czerwczak, M. Spatial Patterns of Trees in Natural Forests of East-Central-Europe. J. Veg. Sci. 4, 469–476 (1993).
Chokkalingam, U. & White, A. Structure and spatial patterns of trees in old-growth northern hardwood and mixed forests of northern Maine. Plant Ecol. 156, 139–160 (2001).
Muth, C. C. & Bazzaz, F. A. Tree canopy displacement and neighborhood interactions. Can. J. For. Res. 33, 1323–1330 (2003).
Jucker, T., Bouriaud, O. & Coomes, D. A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Funct. Ecol. 29, 1078–1086 (2015).
Acknowledgements
Components of this research were funded by the Canadian Forest Service, and a NSERC Discovery grant to Nicholas C. Coops. Chris Hopkinson (previously of Nova Scotia Community College and now with the University of Lethbridge) is thanked for his transect project partnership and his leadership of the national Canadian Consortium for Lidar Environmental Applications Research (C-CLEAR) which was critical in obtaining the research data used in this study. Components of this research was undertaken as part of the “National Terrestrial Ecosystem Monitoring System (NTEMS): Timely and detailed national cross-sector monitoring for Canada” project jointly funded by the Canadian Space Agency (CSA) Government Related Initiatives Program (GRIP) and the Canadian Forest Service (CFS) of Natural Resources Canada. Christopher Bater (previously of UBC and now with the Government of Alberta) is thanked for his analysis efforts and insights into the processing of the lidar transects, and Paul Pickell for statistical advice in the MODIS GPP comparison. Trevor Milne of Gaiamatics is thanked for assisting with the development of customized code for processing the long lidar transect files. We thank all reviewers and editorial members for their commitment to improving this manuscript.
Author information
Authors and Affiliations
Contributions
T.G. and P.T. conducted analysis. T.G. wrote text and created figures with editorial help from all other authors (N.C.C., J.C.W., M.A.W., P.T. and M.S.).
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Goodbody, T.R.H., Tompalski, P., Coops, N.C. et al. Uncovering spatial and ecological variability in gap size frequency distributions in the Canadian boreal forest. Sci Rep 10, 6069 (2020). https://doi.org/10.1038/s41598-020-62878-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-62878-z
- Springer Nature Limited