
Abstract
Juniperus thurifera is a key element of the forest communities in arid and semi-arid areas of the western Mediterranean. Previous genetic and morphological investigations suggested that Algerian populations are genetically more similar to European than to Moroccan populations and advocated their recognition at the variety rank. We aimed to investigate the spatial genetic structure in J. thurifera to verify the distinct character of the Algerian population in terms of the genetic breaks reported among several North African taxa. We also modelled species distributions since the Eemian to recognise the impact of past climatic changes on the current pattern of diversity and predict possible changes in species distribution in the future. Species-specific microsatellites were used in the analysis of 11 populations from Algeria, Morocco and Europe. We revealed the significant genetic distinctiveness of the Algerian populations from the Moroccan and European stands that may have important taxonomic and conservation implications. The diversity pattern revealed for J. thurifera reflects the east-west genetic splits reported among some North African plant and animal taxa and suggests an impact of shared historical processes. Additionally, modelling of the distribution allowed us to identify possible glacial refugia and their impact on the modern pattern of differentiation in J. thurifera. Reduction of species occurrence, especially in the European domain, is likely according to the future projections of the species distribution.
Similar content being viewed by others

Introduction
Compared to the northern Mediterranean, the genetic diversity and differentiation patterns of woody species in the southern part of the region that comprises North Africa is rather poorly explored1,2. However, in some studies that were focused on the evolutionary history of the North African tree species, the distinctiveness between Moroccan and Algerian/Tunisian populations emerges recurrently. In investigations on the cpDNA variability of Quercus canariensis3 and Q. ilex4 and mtDNA diversity of Pinus pinaster5, the Algerian populations were shown to have different haplotypes from those occurring in Morocco; a similar conclusion was drawn for Olea europea6. Furthermore, Moroccan stands of Cedrus atlantica and Alnus glutinosa are also genetically distinct from the Algerian populations7,8. The divergence revealed in A. glutinosa also included differences in ploidy level, such as Moroccan species displaying tetraploidy in comparison to the diploidy displayed by Algerian and European populations. Phylogeographic parallels among plants are accompanied by numerous examples among animal species9,10,11,12, and both advocate for a more general hypothesis of the phylogeographic convergence among North African biota. Such a pattern would assume an east-west split among North African biota, implying a common historical process in action.
Juniperus thurifera L. (thuriferous juniper) occurs in Europe and North Africa but the species range is greatly fragmented13,14,15,16,17. The major distribution area covers the mountains of the Iberian Peninsula, followed by the areas of the High and Middle Atlas Mountains, with a separate subpopulation in the Anti-Atlas in Morocco14,18,19,20. The species distribution has even been observed to extend into Spain through colonization of abandoned fields and abandonment of livestock operations21. Beyond the Iberian Peninsula, the species is considered rare in Europe, and it occurs in the Southern Alps, the Pyrenees and Corsica. In reference to the African range, the Moroccan populations are extremely degraded18,22, but the most isolated and confined populations of this species are in Algeria22.
In Algeria, J. thurifera is exclusively found in six locations in the Aurès Mountains, which are the eastern continuation of the Saharan Atlas Mountain range and are considered to be one of the 39 key biodiversity areas in northern Algeria23. The areas with the most inventories indicated the existence of relatively dense stands consisting of 3,500 to 8,000 mature individuals24. In addition to the climate-driven contraction of juniper’s range in Africa, its decline is strictly related to human pressure. First, it has declined as a result of overexploitation due to logging and wood harvesting, because juniper wood has been used in North Africa as a fuel for the last several millennia16. Second, maintaining a high level of livestock and traditional pasture culture in Mediterranean countries, which results in overgrazing, is also an important factor affecting the species decline in Algeria and Morocco25. Overgrazing and branch cutting negatively affect the regeneration ability of this species, since the organic matter that accumulates beneath the trees’ crowns is the most suitable site for recruitment16. A lack of such conditions hampers regeneration and leads to demographic collapse. Currently, all Algerian stands suffer from a lack of natural regeneration, and old individuals prevail in most of the populations, which probably also decreases their reproductive abilities. Generally, thuriferous juniper in Algeria produces seeds sporadically, and most of them are empty (Taib, personal observation), which suggests the occurrence of some serious environmental or genetic factors (or likely both) that interfere with the reproduction process, causing its failure. Additionally, in all stands, an imbalanced sex ratio is noted, and female trees represent a maximum of 20% of the population size. Since recruitment success is highest under female trees, an insufficient number of such trees in a population is another factor that negatively affects the regeneration process26.
The highly fragmented distribution of J. thurifera corresponds to its morphological and genetic differentiation, which led to subdivision into intra-specific taxa. The major taxonomic split follows the Ibero-African species distribution and resembles the isolating effect of the Strait of Gibraltar27,28,29. The morphological and genetic distinctiveness of populations from Europe and North Africa led to the recognition of the two subspecies: J. thurifera subsp. thurifera and J. thurifera subsp. africana (Maire) Romo et Boratyński20,30,31,32,33,34. Within the European species range, the geographic varieties are described, namely, J. thurifera var. gallica in the Alps and the Pyrenees, J. thurifera var. hispanica18 in Spain and J. thurifera var (chimovar.) corsicana Gauquelin, Idr. Hass. & P. Lebreton in Corse35. A suggestion to separate the Algerian relict populations under the name J. thurifera var. aurasiaca Véla & P. Schäf., var. nov. was made by Vela and Schäfer35 based on the geographic and morphological criteria. The distinctiveness of the Algerian stands was confirmed by Zeraib et al.36 in chemotaxonomic studies. However, the intermediate genetic character of a single Algerian population analysed by Terrab et al.37 suggested that it was more closely genetically related to the European stands.
According to the International Union for Conservation of Nature (IUCN), J. thurifera has been classified as being of least concern, which may be reasonable in the case of Spanish populations that occupy large areas38. However, in North Africa, species exhibits population fragmentation, reduced regeneration and strong decline in Algeria39. Particularly, investigations of Algerian populations are important because they may deliver information about the species’ genetic resources and reveal whether those populations form a distinct genetic lineage that should be recognized as a separate conservation unit. The very marginal location of the Algerian populations in the Aurès Mountains that practically precludes gene flow from other parts of the species’ geographic range may pose a threat to its long-term survival, especially in terms of ongoing climate change40. Moreover, genetic responses to climate-driven environmental changes will likely be based on standing genetic diversity, which requires the maintenance of high within-population variability41.
In this paper, genetic analysis was undertaken to examine the differences between the Algerian populations of J. thurifera in reference to the Moroccan and European stands, as previous studies were unconvincing37. We then asked two questions: (i) Does the spatial pattern of the genetic diversity of J. thurifera subsp. africana match the east-west genetic breaks reported for some North African organisms? and (ii) Does the intraspecific genetic differentiation have taxonomic and hence conservation implications for the Algerian populations? To predict the impact of climate change and evaluate the potential risks involved, species distribution modelling (SDM) was performed. Recently, this method has been widely used in simulations of future changes in species distribution patterns induced by climate change and has been recognized as a useful tool that may efficiently support conservation genetic studies and further planning of conservation activities42,43,44.
Materials and methods
Plant material
Nine natural populations of J. thurifera from Spain, France (Corsica) and Morocco (Middle and High Atlas Mountains) that were previously biometrically analysed by Boratyński et al.33 and two populations from Algeria (Aurès Mountains) were used in this study for the screening of polymorphisms at nuclear microsatellite loci (nSSR) (Table 1, Fig. 1A). However, permission for material collection in Chelia and Tafrent in Algeria was very restricted, and we were able to access only a very limited number of individuals (Table 1). Leaves were collected from 10–25 randomly distributed individuals from each population; 255 individuals were sampled in total.

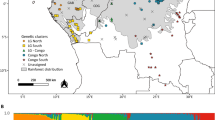
Location of the J. thurifera populations subjected to genetic analysis and populations genetic structure based on six nSSR loci: (A) – bar plots obtained from a discriminant analysis of principal components (DAPC) with genetic barriers obtained with Monmonier’s algorithm (bold lines); (B) – proportion of membership of each individual in the six assumed clusters (K = 6) according to a Bayesian approach estimated by STRUCTURE; (C) – estimation of the optimal number of genetic clusters following Evanno’s ΔK method (Evanno et al. 2005). Map prepared with QGIS.
DNA extraction, amplification and sequencing
Genomic DNA was extracted from dried leaves using the CTAB method45. A set of six nSSR markers originally developed for J. thurifera that were highly polymorphic according to Teixeira et al.34 were used: JT_01, JT_04, JT_30, JT_33, JT_40 and JT_46 (Supplementary Table S1). Two multiplex PCRs were performed in a final volume of 10 µL containing approximately 60 ng of template DNA, 1 U of Silver Hot Start DNA Polymerase (Syngen Biotech, Poland), 1 µM of each primer pair, 1x reaction buffer, 2.5 mM MgCl2, and water. The first multiplex reaction involving loci JT_01, JT_30 and JT_33 and the second one with loci JT_04, JT_40 and JT_46 were amplified in a Labcycler Basic thermocycler (SensoQuest, GmbH) with the following conditions: an initial denaturation step of 15 min at 95 °C, followed by 40 cycles of denaturation at 94 °C for 30 sec, annealing at a temperature specific to each multiplex for 90 sec (57 °C for multiplex I and 55 °C for multiplex II), and extension at 72 °C for 60 sec, with a final extension step at 72 °C for 10 min. PCR products were analysed using an AB 3130 Genetic Analyzer (Thermo Fisher Scientific, USA) capillary electrophoresis system with an internal size standard, GeneScan™ 500 LIZ®. Genotypes were scored using GeneMapper 4.0 software (Thermo Fisher Scientific, USA).
Data analysis
Genetic diversity and differentiation
J. thurifera is a tetraploid species34,46, which has important practical consequences for the scoring of genotypes and subsequent statistical analysis of nSSR data47,48. There is substantial ambiguity in genotype assignment due to the difficulty of resolving the number of allele copies in polyploids. This methodological issue leads to situations in which genotypes cannot be truly inferred based on phenotypes; therefore, neither allele frequencies nor genotype frequencies can be estimated, which are necessary to implement population genetic-based inferences47,49. Additionally, fixation of heterozygosity is a common feature, because genetically different isoloci may share the same alleles, which precludes statistical evaluation of deviation from the Hardy-Weinberg equilibrium or gene diversity level50.
GenoDive 2.0b23 software51, which enables population genetic analysis of polyploids by correcting for allele copy number ambiguity, was used for computations of basic diversity parameters, such as the number of alleles (N), average number of alleles (Na), effective number of alleles (Ne), number of private alleles (Pa) and heterozygosity within populations (Hs). The missing alleles in genotypes were filled with the option overall allele frequencies. The overall genetic differentiation among populations was assessed using the fixation index, Gst52, and tested with 9,999 permutations. Finally, pairwise population differentiation (Fst) was calculated from an analysis of molecular variance (AMOVA), which is exactly equivalent to Weir and Cockerham’s53 statistic, and the significance of Fst was tested with 9,999 permutations.
Geographic patterns of differentiation
We applied different multivariate analysis methods for detecting population structure that can be applied to polyploids48. First, AMOVA-based k-means clustering analysis54 implemented in GenoDive was applied. This method uses the AMOVA framework to calculate the among-cluster sum of squares and divides individuals into an a priori assigned number (k) of groups in such a way that the among-cluster sum of squares is maximized based on a simulated annealing algorithm. Analysis was performed on all generated datasets for k values from 2 to 9, using 20 repeats of the simulated annealing algorithm, with 50,000 MCMC steps. To estimate the most likely number of clusters (optimal k), the Bayesian information criterion (BIC) and Caliński and Harabasz pseudo-F statistic55 methods were used. The clustering results that were biologically justified were discussed, according to the recommendation of Meirmans54.
Second, to infer the geographical pattern of genetic differentiation and detect potential genetic admixture, a discriminant analysis of principal components (DAPC)56 implemented in package ‘adegenet’ in R 3.4.357,58 was used. This multivariate analysis provides an alternative to model-based analyses, because it is not limited by assumptions about the population genetic models; thus, it may serve as a more efficient tool for unravelling complex population structure56. Analysis was performed on a genetic dataset that was transformed to a binary matrix of the presence and absence of each allele without defining groups a priori. DAPC is a multivariate two-step procedure in which data are first transformed by principal component analysis (PCA) as a prior step to a discriminant analysis (DA). The function ‘find cluster’ was used to identify the optimal number of clusters based on the BIC. The number of principal components (PCs) to be retained for the DAPC was chosen according to the α score using the ‘optim.a.score’ function; the DAPC was conducted based on 12 PCs.
Monmonier’s maximum-difference algorithm implemented in BARRIER 2.259 was used to identify genetic discontinuities among populations. This approach was confirmed to be efficient in identification of intraspecific lineages60. The analysis was run using Nei’s genetic distance matrix and the geographical coordinates of each population. The significance of each inferred barrier was assessed by bootstrapping with 100 replications of the genetic distance matrix. SEQBOOT and GENDIST programmes implemented in the PHYLIP package 3.696 were used to obtain multiple matrices data for bootstrapping.
Finally, a model-based Bayesian clustering approach implemented in STRUCTURE 2.3.361 was used to determine the spatial genetic structure. A non-spatial admixture model with correlated allele frequencies was used. To deal with the 4n ploidy level that generates genotype ambiguity, we used a recessive alleles option as recommended by the software’s manual and recent paper of Meirmans48. Ten independent repetitions for each number of groups (K) ranging from 1 to 12 were performed with a burn-in set of 100,000 steps, followed by 200,000 MCMC iterations. The CLUMPAK platform62 was used to align replicated runs and average individual assignment probabilities for the most likely K-value and to estimate the optimal number of clusters according to Evanno’s ΔK method63.
To determine the hierarchical distribution of genetic variation, AMOVA implemented in GenoDive was performed for two defined configurations: among geographic regions (Europe, Morocco and Algeria) and groups revealed by clustering analysis conducted by DAPC (k = 4). The significance was tested with 9,999 permutations, and distances were calculated using the ploidy independent infinite alleles model (Rho). To test for isolation by distance (IBD), the geographic distance matrix was correlated with the genetic distances (Fst) using the standard Mantel test implemented in GenoDive and significance was assessed with 9,999 permutations.
Graphical presentation of genetic data was made with using the QGIS Geographic Information System64.
Modelling of species distributions
The species distribution model for J. thurifera was built using the maximum entropy approach implemented in MaxEnt 3.4.165,66. The data on occurrences of J. thurifera were obtained from the literature and personal observations (Supplementary Table S2). All data were carefully verified, and a total of 280 points were accepted for analysis19,37,46,67. Bioclimatic variables were retrieved from the WorldClim database68 and CHELSA 1.269 to construct models presenting the species’ current potential distribution based on climatic suitability (c.1960–1990; PRE) and that at the last glacial maximum (LGM; 22 ka BP). The CHELSE database has been shown to deliver more precise estimations in terms of precipitation than WorldClim, because the precipitation estimation algorithm is based on statistical downscaling of atmospheric temperatures, which delivers improved climatologies, especially in mountainous landscapes69. The spatial resolution of 19 bioclimatic variables68 (Supplementary Table S3 and S4) used to model the current species range was 30 arc-sec, and for the LGM scenario, the Community Climate System Model (CCSM4) with a 2.5 arc-min resolution was used70. Additionally, to predict future distributional changes in the species’ range, the RCP 2.6 scenario of climate change was tested for the CCSM4 model at a 2.5 arc-min resolution. This scenario assumes that before the year 2100, the atmospheric CO2 concentration will reach 440 ppm and increase the radiative forcing by 2.6 W/m2 as well as that the increase in global mean surface temperature by the end of the 21st century (2081–2100) will be 0.3 °C to 1.7 °C.
The bootstrapping analysis was conducted with 100 replicates, the convergence threshold was set to 10–5, and the maximum number of iterations was 10,000. The ‘random seed’ option was applied, 20% of the data were set aside as test points, and the output was set to logistic. To reveal areas of long-term species persistence, such as Pleistocene refugia, we also modelled the Eemian (c. 120–140 ka BP) theoretical range of J. thurifera in Europe and Africa. However, the Eemian projection was available only with WorldClim. We performed modelling of the present and LGM distributions using occurrence data (1) jointly for African and European populations, (2) for African populations only and (3) for European populations only using both climatic datasets. As a criterion of model accuracy, the receiver operating characteristic (ROC) curve and value of the area under the curve (AUC) were used71,72. The results of the analyses were visualized in QGIS 2.18.2064.
In addition, the relative influence of each bioclimatic variable on the current distribution of J. thurifera was verified by PCA analysis in order to characterize ecological differences between the African and European populations. Analysis was performed on dataset generated for 583 occurrence records of species (Supplementary Table S5) based on bioclimatic raster retrieved from CHELSA using SAGA GIS software73. Function ‘vif” from package usdm in R was used to calculate the variance inflation factor (VIF). Ten climatic variables, which had the highest VIF values (> 300), were removed from the analysis to avoid autocorrelations. PCA analysis was run using the ‘prcomp’ function implemented in R.
Graphical presentation of modelling results was made using the QGIS Geographic Information System.
Results
Genetic diversity and differentiation
All loci used were polymorphic, and the number of alleles ranged from 12 (JT_04) to 44 (JT_33), for a total of 144 alleles detected across the 255 individuals analysed (Supplementary Table S1). In terms of allelic diversity, the Algerian populations were less variable (average Na = 12.500; Ne = 5.985) than the European (average Na = 13.166; Ne = 5.576) and Moroccan populations (average Na = 13.500; Ne = 5.460) (Table 1). In all populations (except for MO_1 and MO_4), private alleles were detected, but the largest number (10) was noted in the European populations. The heterozygosity within populations (Hs) ranged from 0.687 (FR_2) to 0.809 (AL_2), with an average value of 0.772. A comparison between the European part of the J. thurifera range and the African populations indicated that the population from Algeria had the highest significant average value of within-population diversity (Hs = 0.808; P = 0.133), but this value was not significantly higher than the other values (Table 1).
The overall genetic differentiation among populations was low but highly significant (Gst = 0.026; P < 0.001). Accordingly, the populations’ pairwise Fst values ranged from 0.002 (between populations MO_1 and MO_3) to 0.059 (between populations FR_2 and AL_1) and were significant (P ≤ 0.001) in most populations (Table 2).
Geographic pattern of differentiation
AMOVA-based k-means clustering analysis identified k = 4 as the most likely number of clusters, and the pseudo-F statistic (Table 1 and Table 3) indicated clear genetic discontinuity among separate geographic regions. The European, Moroccan and Algerian populations formed three discrete groups. Substructure was detected within the European population; specifically, one Corsican population (FR_2) was detached from the Spanish populations. According to the BIC, populations were assigned to nine distinct groups (k = 9) with no clear geographic pattern (Table 1 and Table 3). This alternative partitioning indicated further substructure of populations within the main regions of the species’ range. Most populations were placed in separate clusters; only the SP_3 and FR_1 populations and two Moroccan populations (MO_1 and MO_3) shared the same clusters (cluster I and II, respectively).
DAPC identified the highest support for k = 4 based on the BIC and confirmed the pattern of differentiation revealed by the AMOVA-based K-means analysis. A clear geographic pattern of differentiation with moderate sharing of gene pools among inferred clusters was detected (Fig. 1A). It was also confirmed at the individual level on the ordination plot of the first two principal components out of 24 included (Fig. 2). Accordingly, all populations from the European part of the species’ range (SP and FR) were mostly assigned to clusters I and III with comparable average membership coefficients (44% and 37%, respectively). DAPC indicated further subdivision of the African populations into two groups with limited sharing of gene pools. The Moroccan populations were grouped mostly into clusters III and IV (average of 26% and 63%, respectively), whereas the Algerian population was put into cluster II (average 65%). Within the latter group, the most distinct population was Chelia (AL_1), which consisted of individuals showing limited genetic affinities to different clusters as compared with the other populations (cluster II, 83%).

Ordination plot for the first two principal component axes resulting from a discriminant analysis of principal components (DAPC) for each individual, ellipses indicate their assignment to the genetic clusters inferred. The low-right graph indicates the variance explained by the principal component axes used for DAPC (dark grey).
According to STRUCTURE, the best K describing spatial genetic structure in analysed populations of J. thurifera was K = 6 (Fig. 1B,C). This result shows mostly the distinctiveness of the Algerian populations from the European and Moroccan stands. The Algerian populations formed rather homogenous group and were mostly assigned to cluster II with moderate gene exchange with other inferred clusters. All remaining populations were rather intermixed, but some general pattern could be inferred. The Moroccan populations were grouped mostly into clusters III and IV. Populations from the European part of the species’ range showed high gene admixture, but the Spanish populations were grouped mostly into clusters I and V and the Corsican population into clusters I and IV (Fig. 1B).
According to the Barrier analysis, three strongly supported genetic boundaries were identified among J. thurifera populations (Fig. 1A). Again, Algerian populations were indicated to be of a distinct character. They were clearly separated from the Spanish and Corsican (100% bootstrap value) and Moroccan (85% bootstrap value) populations. Additionally, barrier analysis also divided the two Algerian populations (98% bootstrap value) that were studied.
AMOVA showed that most of the genetic variation (94%) occurred within populations, while the variation attributable to geographic regions and genetic clusters defined by the clustering analysis accounted for 3.6% and 4.5% of the total variation, respectively (Table 4). A Mantel test revealed a significant positive correlation between the genetic and geographic distances (r = 0.334, P = 0.005).
Modelling of species distributions
The MaxEnt model accuracy for present predictions of the J. thurifera distribution for WorldClim, expressed by the AUC, reached 0.974 for the combined European and African stands, 0.983 for European-only stands and 0.990 for African-only stands, indicating high model performance. Similarly high accuracy was obtained with the CHELSA dataset—0.972 for both African and European records, 0.983 for European-only records and 0.990 for African-only records. The most important factors limiting the current species range modelled jointly for the European and African datasets using WorldClim were the mean temperature of the coldest quarter (BIO11 – relative contribution: 28.6%), minimum temperature of the coldest month (BIO6 – relative contribution: 22.8%), mean temperature of the driest quarter (BIO9 – relative contribution: 11.4%) and precipitation of the warmest quarter (BIO18 – relative contribution: 10.2%) (Supplementary Table S3). The set of factors used to model the current distribution of the African subspecies was different only in terms of the number of important bioclimatic variables and their input (Supplementary Table S3). Nevertheless, BIO11 remained the most important (31.7%), the second critical variable was BIO18 (22.8%), and the remaining factors were of marginal influence. For the European range, some differences also appeared with respect to the strength of the variables. Contrary to the results of the joint analysis, the most important bioclimatic variable was BIO6 (24.0%), while BIO11 was the second most important (17.1%). In reference to the two remaining variables, BIO9 was less important (7.7%), and BIO18 made a relative contribution comparable to that in the joint analysis (11.6%).
Estimations made with the CHELSA dataset indicated a different set of factors that had the highest contribution to the model (Supplementary Table S4). The major difference relates to BIO14 (precipitation of the driest month). It was defined as the second most important variable affecting distribution of the African populations (28.7%), while WorldClim computations marginalized it (Supplementary Table S3). This bioclimatic variable had a very limited impact on the European distribution (3.1%), according to CHELSA. BIO6 remained significant, and its relative contribution was comparable in joint analysis (21.7%) to modelling using WorldClim (22.8%). However, using CHELSA, it was definitely higher for African-only (38.6%) as compared to European-only (10.3%) in comparison to WorldClim-based estimations (Supplementary Table S3). Contrary to WorldClim, modelling with CHELSA indicated that BIO1 (annual mean temperature) was also significant for African range (13.9% of contribution), while it was defined as being marginal in the case of WorldClim-based analysis. BIO11, which was indicated as affecting the African range at most according to WorldClim-based analysis (31.7%), turned out to be important for European-only modelling (17.6%), and for African populations only at negligible level (1.7%). The same situation was observed with BIO18, which had a high relative contribution for European-only estimations (16.7%) and low for African-only estimations (2.9%).
The theoretical contemporary range of J. thurifera modelled using both climatic datasets were very similar. Simulations of the range made jointly for the European and African data covered mainly the area of the Iberian Peninsula, with the most favourable conditions in the Central System and the Penibaetic System at the southern fringe of the Iberian Peninsula (Fig. 3A–C and, Supplementary Fig. S1, Fig. S2A–C). The same high suitability (65%) was also indicated for the southwestern Alps. In these areas, numerous records of species occurrence are available. Less favourable areas for J. thurifera were defined in the Pyrenees. Also, the Massif Central seems to possess habitats climatically suitable for the species, but no observations confirm its growth there. In reference to Africa, the High Atlas is the only location that may highly support the occurrence of J. thurifera, while in Algeria, only very limited areas in the Aurès Mountains offer suitable habitats. However, modelling the range separately for the African and European data revealed profound modification of expected distributions (Fig. 3B,C). Based on the European dataset, MaxEnt supported observed locations of the species in the Iberian Peninsula and in the Alps, but the African range was considered marginal (Supplementary Fig. S2). In contrast, modelling the distribution using African occurrences strongly supported the observed species occurrence in the High Atlas, and the suitability in the Aurès Mountains was much lower.

Theoretical current range of J. thurifera, estimated using MaxEnt based on raster data from the CHELSA database: (A) – European and African records; (B) – African-only records; (C) – European-only records. Map prepared with QGIS.
Modelling the last interglacial distribution (Eemian) jointly for the European and African datasets (WorldClim) suggested a profound reduction of occurrence for J. thurifera (Fig. 4A). Only current core range areas in the Iberian Peninsula, the Alps and the High Atlas and Anti-Atlas were indicated as climatically suitable but with very low support (c. 5%). Analysis of the LGM based on WorldClim using all occurrence records (Fig. 4B) and that using only the African data (data not shown) failed to identify any suitable locations for J. thurifera during this period. Similarly, attempts to reconstruct the species’ range during the LGM using the CHELSA dataset gave no results for any of the combinations used (data not shown). The LGM distribution outlined based on the European-only data retrieved from WorldClim was vestigial to the current one (Fig. 4C). Suitable conditions may have existed in the Iberian Peninsula, probably in the Alps and to some extent in the Anti-Atlas and High Atlas. Generally, the species was likely severely limited (Fig. 4C). However, a dramatic reduction in the species distribution is projected under future climatic conditions based on WorldClim data (Supplementary material Fig. S3). Except for limited areas in the High Atlas, the species practically disappears. Projections made using climatic records from CHELSA are more optimistic (Fig. 5A–C). It predicts some reduction of the range when the datasets for both subspecies are considered. The niche modelling based on the Africa-only dataset indicated that, for African subspecies, the areas of the Penibaetic System in southern Spain may potentially deliver climatically suitable habitats in the future (Fig. 5B; suitability> 65%).

Theoretical range of J. thurifera in the past, estimated using MaxEnt based on raster data from WorldClim database: (A) – Eemian (c. 120–140 ka BP); (B) – LGM (c. 22,000 years ago), all records; (C) – LGM, European-only records. Map prepared with QGIS.

Theoretical range of J. thurifera in the future, estimated using MaxEnt based on raster data from the CHELSA database (year 2070): (A) – European and African records; (B) – African-only records; (C) – European-only records. The RCP 2.6 scenario of climate change was used for the CCSM4 model. Map prepared with QGIS.
PCA analysis of bioclimatic dataset gained from CHELSA supported the ecological differentiation among European and African populations of J. thurifera. Accordingly, all species occurrence records were divided into two main groups according to the first two principal components (PC1 and PC2), which explained 70.9% of the total variation (Fig. 6). The component PC1 (45% of the total variation) was negatively correlated with isothermality (BIO3) and precipitation seasonality (BIO15) and positively correlated with variables that refer to precipitation (BIO12, BIO13, BIO18 and BIO19). The temperature variability (BIO6 and BIO8) and precipitation variables (BIO13, BIO15 and BIO19) contributed mainly to the PC2 component (25.9%) (Fig. 6; Supplementary material Table S5). However, in terms of climatic preferences, it seems that Algerian populations are more similar to European stands.

Influence of 19 bioclimatic variables on the current distribution of J. thurifera according to principal component analysis (PCA) based on 280 species occurrence data (acronyms of bioclimatic variables as in Table S3).
Discussion
North African phylogeographic congruencies include J. thurifera subsp. africana
The most striking result in our study was the recurrent genetic distinctiveness of the Algerian juniper populations from the Moroccan stands, as revealed by different analysis used in this study (Figs. 1 and 2). The pairwise Fst comparisons supported this distinct genetic character of the Algerian stands in reference to Spanish and Moroccan populations, with the latter two being more related to each other. General differentiation between European and African populations of J. thurifera (Fig. 1), was comparable to that obtained by Teixteira et al.34 but much lower than that revealed by Terrab et al.37 (3.6% here vs. 21.44% in Terrab et al.37). This difference probably reflects the different types of markers used in the latter study, particularly the dominant nature of AFLP markers. However, we observed higher differentiation (4.5%) the four genetic clusters detected (Fig. 1A), which points to a significant geographically driven pattern of differentiation in J. thurifera. The influence of geographic location was also supported by significant IBD. This result is certainly an effect of the spatial structure of the species’ natural range, being divided into a few isolated distributional domains in the complex mountainous landscape that currently limits gene flow and likely has hampered it in the past (Fig. 1A).
Our results on Moroccan-Algerian genetic split in J. thurifera subsp. africana are consistent with a few other intra-species diversification profiles noted among western Mediterranean plant and animal species that indicate shared spatial genetic structure5,7,8. The question of whether the genetic split in J. thurifera subsp. africana exemplifies the vicariance model or reflects separate migration pathways to eastern and western North Africa remains unanswered. In other words, is the Moroccan-Algerian divergence a secondary or primary phenomenon in the evolutionary history of J. thurifera in Africa?
According Jimenez et al.20, the colonization of Africa by the ancestor of modern J. thurifera might have taken place from the Iberian Peninsula via the Strait of Gibraltar. However, this conclusion was based solely on investigations of the Spanish and Moroccan stands. The earliest diversification time estimated for J. thurifera by Mao et al.74 was in the Oligocene/Miocene period (31.7–16.4 Ma). This matches the prominent geological rearrangements in the region that left detectable signals in the genetic structure of some species (e.g., Quercus suber75). The most recent estimates date to the Miocene/Pliocene (5.9–3.7 Ma), which is also acknowledged as a period of high diversification in the Mediterranean triggered by climatic changes76. According to geological reconstructions77, in both of these periods, especially in the Miocene, independent colonization of the African continent from the western and eastern directions was feasible. The largest number of private alleles that we recorded in Spanish populations (Table 1) confirms their long-term persistence and thus probably the general ancestral nature of the European gene pool in reference to the African one. However, this pattern does not preclude the independent two-directional colonization of Africa from Europe.
In contrast to our results, in the study of Terrab et al.37, closer genetic relationships of a single Algeria population to European ones were inferred, especially in reference to the Iberian range (Fig. 1 in Terrab et al.36). The two Algerian populations studied here clearly separated from the populations from the other regions, although they showed very limited admixture of the Spanish and Moroccan gene pools (Fig. 1). This genetic overlap might be interpreted as evidence of a common origin20 or ancient gene flow. Terrab et al.36 argued that long-distance dispersal events were the causal factors of the closer clustering of the single Algerian population to European ones. Inspection of a single herbarium individual from Algeria suggests high morphological similarity to European J. thurifera subsp. thurifera35. Although probable, this issue definitely requires more comprehensive morphometric analysis, including more populations and individuals from the Algerian range. However, the influence of the European gene pool needs to be considered in the case of the Algerian populations of J. thurifera. In fact, genetic connections across the Mediterranean Sea between North Africa and the central Mediterranean (Europe) have been demonstrated previously for P. pinaster5, Laurus nobilis78, Carex panormitana79 and J. phoenicea80, which suggests the complex evolutionary history of plant species in this part of the Mediterranean. Even more puzzling is the genetic affinity of Algerian and Tunisian populations of A. glutinosa to a single Scottish stand and not to geographically much closer Moroccan populations8. In terms of such unexpected genetic proximity, the authors concluded independent (including temporal) colonization of the western and eastern parts of North Africa by ancestral populations of A. glutinosa. Accordingly, Algerian and Tunisian populations are more recent colonizers, while Moroccan populations are defined as relicts. The diversification scenario proposed for the North African firs also involves two temporally and spatially independent colonization routes from Europe to North Africa81. The first one, depicted for the ancestor of Abies pinsapo and A. maroccana, would have run from the Iberian Peninsula and might have been related to the evolution of the Betic-Rif orogenic belt since the Miocene82. The second route dated to the Pliocene-Pleistocene would have involved colonization from the Apennine Peninsula and subsequent vicariance, resulting in the evolution of A. numidica81. Ultimately, the problem of colonization routes and their temporal aspects in the evolutionary history of J. thurifera subsp. africana remains unsolved in this study since solving it would require molecular dating and better sampling of the Algerian range. However, there is still ample evidence to consider the bidirectional colonization of North Africa, which is a convincing argument explaining the west-east genetic differentiation of J. thurifera subsp. africana and other plant and animal species in the region.
Pleistocene/Holocene changes in species ranges: possible adaptation-driven divergence?
The impact of the Pleistocene glaciations, particularly the last glacial period, on the genetic patterns in North Africa is far less recognized than that on the European part of the Mediterranean, but it is invoked as a factor inducing patterns of endemism and differentiation29,76,83. Our species ecological niche modelling revealed interesting but very puzzling results. Surprisingly, projections made for the periods spanning the time from the Eemian (c. 120 ka BP) to the current time clearly suggest that the widest theoretical species occurrence is the one under current climatic conditions (Fig. 3A–C and, Supplementary Fig. S1, Fig. S2A–C). Meanwhile, it is commonly assumed that the rise of Holocene temperatures was the factor responsible for the reduction and fragmentation of the ranges of many Juniperus species occurring in the Mediterranean, and the environmental conditions during the glacial period promoted range expansion20,34,44. Another unexpected result from our projections concerns the mutually exclusive distribution patterns obtained when using the African-only or European-only dataset on species locations that may indicate climate-based adaptive divergence (Fig. 4B,C; Supplementary Fig. S2A–C).
Climatic conditions during the last glacial period led to the formation of glaciers in North African mountains, especially in the High Atlas84, while lower precipitation did not create suitable conditions for extended glacier formation in the Aurès Mountains85. Due to insufficient snow accumulation, relatively high annual temperatures and low precipitation at the highest points, glaciers are currently absent from the Atlas Mountains86. Consequently, the formation of glaciers in the High Atlas during the last glacial period would have required either (1) a significant decrease in the annual temperatures (by c. 7–8 °C) and the current level of precipitation or (2) more precipitation than currently observed and a smaller decrease in the mean annual temperatures86. J. thurifera subsp. africana prefers warm and dry habitats and exhibits strong continentalism; it is limited by the influence of the Atlantic climate19. Hence, irrespective of the climatic reconstruction supporting glacier formations in the High Atlas, the conditions for occurrence of J. thurifera subsp. africana were inadequate in North Africa. Consequently, MaxEnt failed to identify any climatically suitable conditions for the persistence of this taxon during the LGM in Africa based on current African-only locations using both climatic datasets (Fig. 4). However, the joint analysis of the European and African records made on the WorldClim dataset indicated some limited areas of species persistence in the Iberian Peninsula and West Alps but with very low suitability (c. 5%) (Fig. 4B). When using only the European records, in addition to the Iberian mountains and the Alps, optimal conditions for the species also appeared in the High Atlas and Aurès Mountains which could serve as refugia (Fig. 4C). Additional regions, such as the Pyrenees and Corsica, from which J. thurifera is reported today, were also indicated as suitable for refugia (Fig. 4C). Currently, this species occurs in Europe in areas with a total annual precipitation of at least 200 mm and a mean annual temperature of 8–10 °C87. Previous estimates of the potential distribution of several Mediterranean tree species from the Iberian Peninsula showed that the LGM range of J. thurifera was greatly limited and matched the one we modelled in our work88. It seems that, generally, climatic conditions for J. thurifera during the last glacial period were less suitable than the current theoretical ones, and its distribution expanded in the Holocene82. The factors affecting species occurrence in Europe during the LGM were related to temperature. The most significant for species occurrence were BIO6 (minimum temperature of the coldest month; 24.8%) and BIO11 (mean temperature of the coldest quarter; 18.2%). Current populations of African subspecies adapted to dry and hot climates could not be present during the LGM likely because of too low temperatures. The most important climatic factor affecting the distribution of J. thurifera subsp. africana (in fact a lack of its occurrence at that time) was BIO11 (31.2%). However, on the second position BIO18 (precipitation of the warmest quarter) was ranked (25.0%), which may suggest that it was also too arid during the LGM for current African taxa. According to Cheddadi et al.83, the mean annual temperatures in North Africa during the glacial period could have even been 15 °C lower than today, and the mean annual precipitation did not exceed 300 mm. In this way, our findings contradict previous assumptions made by some authors that the last cold period favoured a wider occurrence of this species20,37. Additionally, there is a compelling similarity between the Eemian (Fig. 4A,C. 120–140 ka BP) and glacial theoretical distribution (Fig. 4C) that suggests continuation of its occurrence in the core range in the Central System in the Iberian Peninsula, the West Alps, the High Atlas and the Aurès Mountains.
Given the mutually exclusive ranges projected with only African or only European records (Fig. 3), we hypothesize that climate-induced adaptation was one of the possible driving factors of the Ibero-African genetic differentiation recurrently revealed for J. thurifera, including in this work. According to the proposed scenario, spatial isolation in refugia defined with MaxEnt in Central System in Iberian Peninsula and Atlas Mountains in Africa (Fig. 4C) and the selective forces related to climatic changes in North Africa allowed genetic differentiation to arise and then be reinforced by genetic drift due to limited gene flow among different domains of the species distribution. First, MaxEnt models suggest that today’s European and African lineages are characterized by differential climatic requirements (Fig. 6, Supplementary Table S3 and Table S4). The most important climatic variables differ, and it seems that the African linage evolved towards enhanced drought resistance, as it is adapted to a dry continental climate with hot summers and low winter temperatures19 (Fig. 6, Supplementary Table S4). Second, the lack of climatic conditions favourable for the current African lineage, since at least the last interglacial, suggests its more recent evolution. Actually, for some species, the Quaternary glacial cycles were shown to be significant drivers of adaptation-based genetic divergence89,90,91. For J. thurifera, the crucial factor was probably the climate changes in North Africa throughout the Holocene. Reconstruction of the vegetation changes in the Middle Atlas based on Lake Sidi Ali, located at an altitude of 2,080 m above sea level, revealed the first occurrence of Cupressaceae (and so Juniperus) in the early Holocene and its expansion in the mid-Holocene92, which was cooler than the early Holocene. However, the general aridification trend in North Africa throughout the Holocene prevailed, and Juniperus withdrew at the expense of Cedrus only in more humid periods92. Our hypothesis of climate-driven differentiation between the European and African lineages is also somewhat supported at the morphological level. The biometric analysis revealed generally smaller dimensions of structures (cones and scales) and a smaller number of seeds per cone in the Moroccan populations as compared to in the European ones, which might be ascribed to adaptation to the severe arid climate of the Atlas Mountains33. However, this issue requires deeper investigations involving searching for genomic signals of adaptation91.
Concluding remarks and perspectives
In this work, we demonstrated that Algerian populations of J. thurifera are a genetically distinct lineage (Figs. 1 and 2) and could be classified as J. thurifera var. aurasiaca, as previously suggested35. This species is one more excellent example of the extremely high intra-species genetic diversity noted in the North African region, representing another hotspot of evolution within the Mediterranean region. Unfortunately, this high biodiversity is presumed to face great environmental challenges that may lead to the irreversible loss of many species and thus plant community depletion. Ultimately, such a loss will affect ecosystem functioning and human well-being. In the African range of J. thurifera, the species is currently subjected to strong human pressure, fragmentation and reproductive failure16. All of these effects are negatively reinforced by ongoing aridification of the region, which makes populations vulnerable to additional disturbances and thereby exposes them to extinction risk. There are well-known examples of species in North Africa that are already trapped in the extinction vortex, such as endemic Cupressus or Abies species29,76. The most pessimistic projections of the future species distribution confirmed the serious reduction and almost extinction of J. thurifera (Supplementary Fig. S3). The predicted climatic deterioration is expected to affect the species’ range not only in Africa but also in Europe, leaving only a few locations of probable occurrence. Currently, J. thurifera is a key component of the high mountain forest communities of arid and semi-arid areas in North Africa and the Iberian Peninsula. A definitely more optimistic scenario was obtained using the CHELSA dataset (Fig. 5). The difference between CHELSA-based and WorldClim-based projections likely stems from the more precise performance of the first one69. Particularly, the precipitation algorithm that works in CHELSA includes orographic predictors such as wind fields or valley exposition with a subsequent bias correction that makes it work better and more reliable in case of the mountainous landscapes. Consequently, our CHELSA-based computations also indicated a reduction of the species’ range but not in such a drastic scale as WorldClim-based computations. In this case, the European part of the range was predicted to be more vulnerable for future climate changes than the African part of the range (Fig. 5 and Supplementary Fig. S3). The core range of the species in the Iberian mountains with the highest suitability (65%) was reduced profoundly (Fig. 5C). For African-based modelling, MaxEnt holds approximately the same areas of distribution as current ones and even projected the appearance of the suitable locations in the most southern fringes of the Iberian Peninsula (Fig. 5B). This particular result yearns for attention, as it allows the idea of assisted migration to be considered in the future to enhance the adaptability of the European population of the species.
Using a combined approach in which genetic analysis was supplemented with ecological niche modelling, we denoted the areas of the current species distribution that have constituted long-term refugia for J. thurifera since at least the Eemian (Fig. 4). Therefore, these areas represent the evolutionary heritage of the species and should thus be prioritized in conservation programmes93. Refugia are areas that have facilitated species survival during past climatic and environmental crises. However, as shown by our modelling, the buffering capacity of these future refugial areas in Central System in Iberian Peninsula (Fig. 5C) can be limited in the case of J. thurifera under expected environmental transformation. If the current trend in climate change continues40, these areas may no longer be efficient in supporting the species’ survival. Therefore, the evolutionary value of the Algerian populations should be immediately acknowledged by including them in a protected areas network, because they are more isolated and spatially confined than Moroccan stands.
As a final remark, we need to stress that predictions of future changes in species distribution done in this work are solely based on the climatic variables. Although they set the environmental boundaries of the species’ current or possible performance, they ignore local adaptations and so species evolvability. Meanwhile including evolutionary potential may change the predictions and fates of species94. However, genetic adaptation that may help to mitigate the climate-induced species and population demographic crisis leading to local extinctions require high levels of genetic diversity stored in populations and maintaining genetic connectivity among them. The more crucial is to focus conservation activity on species such as the fragmented and isolated J. thurifera subsp. africana in Algeria.
References
Nieto Feliner, G. Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect Plant Ecol 16, 265–278 (2014).
Bouahmed, A., Vessella, F., Schirone, B., Krouchi, F. & Derridj, A. Modeling Cedrus atlantica potential distribution in North Africa across time: new putative glacial refugia and future range shifts under climate change. Reg Environ Change https://doi.org/10.1007/s10113-019-01503-w (2019).
Petit, R. J. et al. Identification of refugia and post-glacial colonisation routes of European white oaks based on chloroplast DNA and fossil pollen evidence. For Ecol Manage 156, 49–74 (2002).
Lumaret, R., Mir, C., Michaud, H. & Raynal, V. Phylogeographical variation of chloroplast DNA in holm oak (Quercus ilex L.). Mole Ecol 11, 2327–2336 (2002).
Burban, C. & Petit, R. J. Phylogeography of maritime pine inferred with organelle markers having contrasted inheritance. Mol Ecol 12, 1487–1495 (2003).
Besnard, G., Khadari, B., Baradat, P. & Bervillé, A. Olea europaea (Oleaceae) phylogeography based on chloroplast DNA polymorphism. Theoret Appl Genetics 104, 1353–1361 (2002).
Terrab, A. et al. Phylogeography of North African Atlas cedar (Cedrus atlantica, Pinaceae): Combined molecular and fossil data reveal a complex Quaternary history. Am J Bot 95, 1262–1269 (2008).
Lepais, O. et al. High Genetic Diversity and Distinctiveness of Rear-Edge Climate Relicts Maintained by Ancient Tetraploidisation for Alnus glutinosa. PLOS ONE 8, e75029 (2013).
Naciri, Y., Cavat, F. & Jeanmonod, D. Silene patula (Siphonomorpha, Caryophyllaceae) in North Africa: A test of colonisation routes using chloroplast markers. Mol Phylo Evol 54, 922–932 (2010).
Nicolas, V., Mataame, A., Crochet, P.-A., Geniez, P. & Ohler, A. Phylogeographic patterns in North African water frog Pelophylax saharicus (Anura: Ranidae). J Zool Syst Evol Res 53, 239–248 (2015).
Lalis, A. et al. Phylogeography and demographic history of Shaw’s Jird (Meriones shawii complex) in North Africa. Biol J Linn Soc 118, 262–279 (2016).
Beddek, M. et al. Comparative phylogeography of amphibians and reptiles in Algeria suggests common causes for the east-west phylogeographic breaks in the Maghreb. PLOS ONE 13, e0201218 (2018).
Gauquelin, T., Bertaudière-Montès, A., Badri, W. & Montès, N. Sex ratio and sexual dimorphism in mountain dioecious thuriferous juniper (Juniperus thurifera L., Cupressaceae). Bot J Linn Soc 138, 237–244 (2002).
Farjon, A. A handbook of the world’s conifers. (Brill, 2010).
Farjon, A. & Filer, D. An atlas of the world’s conifers: an analysis of their distribution, biogeography, diversity, and conservation status. (Brill, 2013).
Gauquelin, T., Bertaudiere, V., Montes, N., Badri, W. & Asmode, J.-F. Endangered stands of thuriferous juniper in the western Mediterranean basin: ecological status, conservation and management. Biodiv Conserv 8, 1479–1498 (1999).
Aymerich, P. & Villar, L. Sur les peuplements de Genévrier thurifère (Juniperus thurifera L.) en Haut Pallars (Pyrénées centrales, Catalogne). Présentation géobotanique et écologique. Ecologia Mediterranea 39, 13 (2013).
Auclair, L. Le genévrier thurifère Juniperus thurifera L. Géant de l’Atlas. Forêt Mediterranéenne 14, 306–314 (1993).
Romo Díez, A. & Boratyński, A. Chorology of Juniperus thurifera (Cupressaceae) in Morocco. Dendrobiology 54, 41–50 (2005).
Jiménez, J. F., Werner, O., Sánchez-Gómez, P., Fernández, S. & Guerra, J. Genetic variations and migration pathway of Juniperus thurifera L. (Cupressaceae) in the western Mediterranean region. Israel J Plant Sci 51, 11–22 (2003).
DeSoto, L., Olano, J. M., Rozas, V. & De la Cruz, M. Release of Juniperus thurifera woodlands from herbivore-mediated arrested succession in Spain. App Veg Science 13, 15–25 (2010).
Gauquelin, T. et al. Le Genévrier thurifère, espèce partagée au Nord et au Sud de la Méditerranée. Forêt Meditérranéenne 3, 227–240 (2012).
Benhouhou, S., Yahi, N. & Vela, E. Key Biodiversity Areas (KBAs) for plants in the Mediterranean region in Conserving wild plants in the South and East Mediterranean region (eds. Valderrábano, M., Gil, T., Heywood, V. & de Montmollin, B.) 53–60 (IUCN & IUCN Centre for Mediterranean Cooperation, 2018).
Farjon, A. Juniperus thurifera. The IUCN Red List of Threatened Species 2013: e.T42255A2967372, https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T42255A2967372.en (2013).
Barbero, M., Bonin, G., Loisel, R. & Quézel, P. Changes and disturbances of forest ecosystems caused by human activities in the western part of the mediterranean basin. Vegetatio 87, 151–173 (1990).
De Peña, M., García-González, M. D. & De Pedro, R. Adéquation du microhabitat pour la germination de Juniperus thurifera L. dans les populations naturelles de Genévrier thurifère en Castilla et León (Espagne). Forêt Meditérranéenne 34, 1–14 (2012).
Jaramillo-Correa, J. P. et al. The Strait of Gibraltar as a major biogeographic barrier in Mediterranean conifers: a comparative phylogeographic survey. Mol Ecol 19, 5452–5468 (2010).
Lavergne, S., Hampe, A. & Arroyo, J. In and out of Africa: how did the Strait of Gibraltar affect plant species migration and local diversification? J. Biogeogr. 40, 24–36 (2013).
Dering, M. et al. Genetic diversity and inter-specific relations of western Mediterranean relic Abies taxa as compared to the Iberian A. alba. Flora 209, 367–374 (2014).
Gauquelin, T., Idrissi Hassani, L. M. & Lebreton, P. Le genévrier thurifère, Juniperus thurifera L. (Cupressaceae): Analyse biométrique et biochimique; propositions systématiques. Ecologia Mediterranea 3/4, 31–42 (1988).
Adams, R. P. et al. Geographic Variation in the Leaf Oils and DNA Fingerprints (RAPDs) of Juniperus thurifera L. from Morocco and Europe. J Essent Oil Res 15, 148–154 (2003).
Romo, A. & Boratyński, A. Nomenclatural note on Juniperus thurifera subsp. africana (Cupressaceae). Ann Bot Fenn 44, 72–75 (2007).
Boratyński, A. et al. Morphological differentiation supports the genetic pattern of the geographic structure of Juniperus thurifera (Cupressaceae). Plant Syst Evol 299, 773–784 (2013).
Teixeira, H., Rodríguez-Echeverría, S. & Nabais, C. Genetic Diversity and Differentiation of Juniperus thurifera in Spain and Morocco as Determined by SSR. PLOS ONE 9, e88996 (2014).
Vela, E. & Schäfer, P. A. Typification de Juniperus thurifera var. africana Maire, délimitation taxonomique et conséquences nomenclaturales sur le Genévrier thurifère d’Algérie. Ecologia mediterranea 39, 69–80 (2013).
Zeraib, A., Ramdani, M., Boudjedjou, L., Chalard, P. & Figuredo, G. Characterization and chemosystematics of Algerian thuriferous juniper (Juniperus thurifera L.). J Appl Bot Food Qua 87, 249–255 (2014).
Terrab, A., Schönswetter, P., Talavera, S., Vela, E. & Stuessy, T. F. Range-wide phylogeography of Juniperus thurifera L., a presumptive keystone species of western Mediterranean vegetation during cold stages of the Pleistocene. Mol Phylogenet Evol 48, 94–102 (2008).
IUCN. The IUCN Red List of Threatened Species. Version 2019-3. Downloaded on 12, December 2019.
Villar, L. The incense juniper, an old and very useful tree for man in western Mediterranean area. Forêt Meditérranéenne 1, 7–16 (2013).
Guiot, J. & Cramer, W. Climate change: The 2015 Paris Agreement thresholds and Mediterranean basin ecosystems. Science 354, 465–468 (2016).
Orr, H. A. & Unckless, R. L. The Population Genetics of Evolutionary Rescue. PLOS Genetics 10, e1004551 (2014).
Wan, D.-S. et al. The Quaternary evolutionary history, potential distribution dynamics, and conservation implications for a Qinghai-Tibet Plateau endemic herbaceous perennial, Anisodus tanguticus (Solanaceae). Ecol Evol 6, 1977–1995 (2016).
Qin, A. et al. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Global Ecol Conserv 10, 139–146 (2017).
Walas, Ł. et al. Past, present, and future geographic range of an oro-Mediterranean Tertiary relict: the Juniperus drupacea case study. Reg Environ Change 19, 1507–1520 (2019).
Doyle, J. J. & Doyle, J. L. Isolation of plant DNA from fresh tissue. Focus 2, 13–15 (1990).
Romo, A. et al. Genome size and ploidy levels in highly fragmented habitats: the case of western Mediterranean Juniperus (Cupressaceae) with special emphasis on J. thurifera L. Tree Genet Genom 9, 587–599 (2013).
Dufresne, F., Stift, M., Vergilino, R. & Mable, B. K. Recent progress and challenges in population genetics of polyploid organisms: an overview of current state-of-the-art molecular and statistical tools. Mol Ecol 23, 40–69 (2014).
Meirmans, P. G., Liu, S. & van Tienderen, P. H. The Analysis of Polyploid Genetic Data. J Hered 109, 283–296 (2018).
Clark, L. V. & Jasieniuk, M. polysat: an R package for polyploid microsatellite analysis. Mol Ecol Resour 11, 562–566 (2011).
Obbard, D. J., Harris, S. A. & Pannell, J. R. Simple allelic-phenotype diversity and differentiation statistics for allopolyploids. Heredity 97, 296–303 (2006).
Meirmans, P. G. & Tienderen, P. H. V. genotype and genodive: two programs for the analysis of genetic diversity of asexual organisms. Mol Ecol Notes 4, 792–794 (2004).
Nei, M. Molecular evolutionary genetics. (Columbia University Press, 1987).
Weir, B. S. & Cockerham, C. C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 38, 1358–1370 (1984).
Meirmans, P. G. AMOVA-Based Clustering of Population Genetic Data. J Hered 103, 744–750 (2012).
Caliński, T. & Harabasz, J. A dendrite method for cluster analysis. Comm Stat 3, 1–27 (1974).
Jombart, T., Devillard, S. & Balloux, F. Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genetics 11, 94 (2010).
Jombart, T. adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24, 1403–1405 (2008).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. (2013).
Manni, F., Guerard, E. & Heyer, E. Geographic Patterns of (Genetic, Morphologic, Linguistic) Variation: How Barriers Can Be Detected by Using Monmonier’s Algorithm. Human Biology 76, 173–190 (2004).
Xu, T.-T. et al. Allopatric divergence, demographic history, and conservation implications of an endangered conifer Cupressus chengiana in the eastern Qinghai-Tibet Plateau. Tree Genet Genomes 13, 100 (2017).
Pritchard, J. K., Stephens, M. & Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 155, 945–959 (2000).
Kopelman, N. M., Mayzel, J., Jakobsson, M., Rosenberg, N. A. & Mayrose, I. Clumpak: a program for identifying clustering modes and packaging population structure inferences across. Mol Ecol Resour 15, 1179–1191 (2015).
Evanno, G., Regnaut, S. & Goudet, J. Detecting the number of clusters of individuals using the software structure: a simulation study. Mol Ecol 14, 2611–2620 (2005).
QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project (2012).
Phillips, S. J., Dudík, M. & Schapire, R. E. A maximum entropy approach to species distribution modeling in Proceedings of the Twenty-First International Conference on Machine Learning, https://doi.org/10.1145/1015330.1015412 (2004).
Elith, J. et al. A statistical explanation of MaxEnt for ecologists. Divers Distrib 17, 43–57 (2011).
UICN. Programme UICN pour l’Afrique du nord: Etude d’Elevage et du Pastoralisme au sein du Parc National de Belezma (Genévrier Thurifère). (République Algérienne, Ministère de l’Agriculture, 2001).
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. & Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25, 1965–1978 (2005).
Karger, D. N. et al. Climatologies at high resolution for the earth’s land surface areas. Scientific Data 4, 170122 (2017).
Gent, P. R. et al. The Community Climate System Model Version 4. J Climate 24, 4973–4991 (2011).
Wang, Z., Chang, Y. I., Ying, Z., Zhu, L. & Yang, Y. A parsimonious threshold-independent protein feature selection method through the area under receiver operating characteristic curve. Bioinformatics 23, 2788–2794 (2007).
Mas, J.-F., Soares Filho, B., Pontius, R. G., Farfán Gutiérrez, M. & Rodrigues, H. A Suite of Tools for ROC Analysis of Spatial Models. ISPRS International Journal of Geo-Information 2, 869–887 (2013).
Conrad, O. et al. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci Model Dev 8, 1991–2007 (2015).
Mao, K., Hao, G., Liu, J., Adams, R. P. & Milne, R. I. Diversification and biogeography of Juniperus (Cupressaceae): variable diversification rates and multiple intercontinental dispersals. New Phyto 188, 254–272 (2010).
Magri, D. et al. The distribution of Quercus suber chloroplast haplotypes matches the palaeogeographical history of the western Mediterranean. Mol Ecol 16, 5259–5266 (2007).
Sękiewicz, K. et al. Phylogenetic and biogeographic insights into long-lived Mediterranean Cupressus taxa with a schizo-endemic distribution and Tertiary origin. Bot J Linn Soc 188, 190–212 (2018).
Popov, S. V. et al. Lithological-Paleogeographic maps of Paratethys - 10 maps Late Eocene to Pliocene. Available at: https://www.schweizerbart.de/publications/detail/isbn/9783510613700/%23. (Accessed: 25th July 2019) (2004)
Marzouki, H. et al. Population Genetic Structure of Laurus nobilis L. Inferred From Transferred Nuclear Microsatellites. Silvae Genet 58, 270–276 (2009).
Benítez‐Benítez, C., Escudero, M., Rodríguez‐Sánchez, F., Martín‐Bravo, S. & Jiménez‐Mejías, P. Pliocene–Pleistocene ecological niche evolution shapes the phylogeography of a Mediterranean plant group. Mol Ecol 27, 1696–1713 (2018).
Sánchez-Gómez, P. et al. Genetic structure and phylogeography of Juniperus phoenicea complex throughout Mediterranean and Macaronesian regions: different stories in one. Ann For Sci 75, 75 (2018).
Sánchez-Robles, J. M. et al. Phylogeography of SW Mediterranean firs: Different European origins for the North African Abies species. Mol Phylogen Evol 79, 42–53 (2014).
Rosenbaum, G., Lister, G. S. & Duboz, C. Reconstruction of the tectonic evolution of the western Mediterranean since the Oligocene. J Virtual Explorer 8, 107–130 (2002).
Cheddadi, R. et al. Putative glacial refugia of Cedrus atlantica deduced from Quaternary pollen records and modern genetic diversity. J Biogeogr 36, 1361–1371 (2009).
Hughes, P. D., Fenton, C. R. & Gibbard, P. L. Quaternary Glaciations of the Atlas Mountains, North Africa. in Developments in Quaternary Sciences 15, 1065–1074 (2011).
Hughes, P. D., Woodward, J. C. & Gibbard, P. L. Quaternary glacial history of the Mediterranean mountains. Progress in Physical Geography: Earth and Environment 30, 334–364 (2006).
Hannah, G., Hughes, P. D. & Gibbard, P. L. Pleistocene plateau ice fields in the High Atlas, Morocco. Geological Society, London, Special Publications 433, 25–53 (2017).
González, A. G., García-Viñas, J. I., Saura, S., Caudullo, G. & de Rigo, D. Juniperus thurifera in Europe: distribution, habitat, usage and threats. In European Atlas of Forest Tree Species (eds J. San-Miguel-Ayanz, D. de Rigo, G. Caudullo, T. Houston Durrant, & A. Mauri) 0195e6+ (Luxembourg, 2016).
Garzón, M. B., Dios, R. Sde & Ollero, H. S. Predictive modelling of tree species distributions on the Iberian Peninsula during the Last Glacial Maximum and Mid-Holocene. Ecography 30, 120–134 (2007).
Mayol, M. et al. Adapting through glacial cycles: insights from a long-lived tree (Taxus baccata). New Phytol 208, 973–986 (2015).
Gugger, P. F., Ikegami, M. & Sork, V. L. Influence of late Quaternary climate change on present patterns of genetic variation in valley oak, Quercus lobata Née. Mole Ecol 22, 3598–3612 (2013).
Ma, Y. et al. Ancient introgression drives adaptation to cooler and drier mountain habitats in a cypress species complex. Commun Biol 2, 1–12 (2019).
Campbell, J. F. E. et al. Environmental Drivers of Holocene Forest Development in the Middle Atlas, Morocco. Front Ecol Evol 5, 113 (2017).
Rodríguez-Sánchez, F., Pérez-Barrales, R., Ojeda, F., Vargas, P. & Arroyo, J. The Strait of Gibraltar as a melting pot for plant biodiversity. Quater Sci Rev 27, 2100–2117 (2008).
Keppel, G. & Wardell-Johnson, G. W. Refugia: keys to climate change management. Global Change Biol 18, 2389–2391 (2012).
Acknowledgements
This study was supported by the Institute of Dendrology, Polish Academy of Sciences and Ecole Nationale Supérieure Agronomique, Algeria.
Author information
Authors and Affiliations
Contributions
A.T. performed plant material collection and laboratory works; A.M., A.B. and A.R delivered material; A.Ch. conducted laboratory works; K.S., Ł.W. and M.D performed statistical analysis; A.T., K.S., M.D. wrote the main manuscript text; M.D gave the idea of studies. All Authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Taib, A., Morsli, A., Chojnacka, A. et al. Patterns of genetic diversity in North Africa: Moroccan-Algerian genetic split in Juniperus thurifera subsp. africana. Sci Rep 10, 4810 (2020). https://doi.org/10.1038/s41598-020-61525-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-61525-x
- Springer Nature Limited
This article is cited by
-
The influence of climate and population density on Buxus hyrcana potential distribution and habitat connectivity
Journal of Plant Research (2023)
-
Environmental regionalization and endemic plant distribution in the Maghreb
Environmental Monitoring and Assessment (2022)
-
High genetic diversity and low future habitat suitability: will Cupressus atlantica, endemic to the High Atlas, survive under climate change?
Regional Environmental Change (2020)