

Abstract
Interleukin-17 (IL-17)-producing helper T (TH17) cells are heterogenous and consist of nonpathogenic TH17 (npTH17) cells that contribute to tissue homeostasis and pathogenic TH17 (pTH17) cells that mediate tissue inflammation. Here, we characterize regulatory pathways underlying TH17 heterogeneity and discover substantial differences in the chromatin landscape of npTH17 and pTH17 cells both in vitro and in vivo. Compared to other CD4+ T cell subsets, npTH17 cells share accessible chromatin configurations with regulatory T cells, whereas pTH17 cells exhibit features of both npTH17 cells and type 1 helper T (TH1) cells. Integrating single-cell assay for transposase-accessible chromatin sequencing (scATAC-seq) and single-cell RNA sequencing (scRNA-seq), we infer self-reinforcing and mutually exclusive regulatory networks controlling different cell states and predicted transcription factors regulating TH17 cell pathogenicity. We validate that BACH2 promotes immunomodulatory npTH17 programs and restrains proinflammatory TH1-like programs in TH17 cells in vitro and in vivo. Furthermore, human genetics implicate BACH2 in multiple sclerosis. Overall, our work identifies regulators of TH17 heterogeneity as potential targets to mitigate autoimmunity.







Similar content being viewed by others


Data availability
Mouse sequence data have been deposited in the Gene Expression Omnibus under the accession codes GSE206175, GSE206304 and GSE206607. Human sequence data are under controlled access at Zenodo at https://doi.org/10.5281/zenodo.11186873 (ref. 82). Access to these data files is available upon request but is otherwise restricted to protect the privacy of the sample donors.
Code availability
No original code was generated for this study. Details on analysis packages used are available in the Methods.
References
Schnell, A., Littman, D. R. & Kuchroo, V. K. TH17 cell heterogeneity and its role in tissue inflammation. Nat. Immunol. 24, 19–29 (2023).
Schnell, A. et al. Targeting PGLYRP1 promotes antitumor immunity while inhibiting autoimmune neuroinflammation. Nat. Immunol. 24, 1908–1920 (2023).
Littman, D. R. & Rudensky, A. Y. TH17 and regulatory T cells in mediating and restraining inflammation. Cell 140, 845–858 (2010).
Bettelli, E., Oukka, M. & Kuchroo, V. K. TH-17 cells in the circle of immunity and autoimmunity. Nat. Immunol. 8, 345–350 (2007).
Blaschitz, C. & Raffatellu, M. TH17 cytokines and the gut mucosal barrier. J. Clin. Immunol. 30, 196–203 (2010).
Stockinger, B. & Omenetti, S. The dichotomous nature of T helper 17 cells. Nat. Rev. Immunol. 17, 535–544 (2017).
Lee, Y. et al. Induction and molecular signature of pathogenic TH17 cells. Nat. Immunol. 13, 991–999 (2012).
Ghoreschi, K. et al. Generation of pathogenic TH17 cells in the absence of TGF-β signalling. Nature 467, 967–971 (2010).
Gaublomme, J. T. et al. Single-cell genomics unveils critical regulators of TH17 cell pathogenicity. Cell 163, 1400–1412 (2015).
Wagner, A. et al. Metabolic modeling of single TH17 cells reveals regulators of autoimmunity. Cell 184, 4168–4185 (2021).
Hirota, K. et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol. 12, 255–263 (2011).
Lee, Y. K. et al. Late developmental plasticity in the T helper 17 lineage. Immunity 30, 92–107 (2009).
Schnell, A. et al. Stem-like intestinal TH17 cells give rise to pathogenic effector T cells during autoimmunity. Cell 184, 6281–6298 (2021).
Pawlak, M. et al. Induction of a colitogenic phenotype in TH1-like cells depends on interleukin-23 receptor signaling. Immunity 55, 1663–1679 (2022).
Harbour, S. N., Maynard, C. L., Zindl, C. L., Schoeb, T. R. & Weaver, C. T. TH17 cells give rise to TH1 cells that are required for the pathogenesis of colitis. Proc. Natl Acad. Sci. USA 112, 7061–7066 (2015).
Yosef, N. et al. Dynamic regulatory network controlling TH17 cell differentiation. Nature 496, 461–468 (2013).
Wang, C. et al. CD5L/AIM regulates lipid biosynthesis and restrains TH17 cell pathogenicity. Cell 163, 1413–1427 (2015).
Zheng, S., Hedl, M. & Abraham, C. Twist1 and Twist2 contribute to cytokine downregulation following chronic NOD2 stimulation of human macrophages through the coordinated regulation of transcriptional repressors and activators. J. Immunol. 195, 217–226 (2015).
Xiao, S. et al. Checkpoint receptor TIGIT expressed on Tim-1+ B cells regulates tissue inflammation. Cell Rep. 32, 107892 (2020).
Kim, S. V. et al. GPR15-mediated homing controls immune homeostasis in the large intestine mucosa. Science 340, 1456–1459 (2013).
Tortola, L. et al. High-dimensional T helper cell profiling reveals a broad diversity of stably committed effector states and uncovers interlineage relationships. Immunity 53, 597–613 (2020).
Meyer Zu Horste, G. et al. RBPJ controls development of pathogenic TH17 cells by regulating IL-23 receptor expression. Cell Rep. 16, 392–404 (2016).
Hiltensperger, M. et al. Skin and gut imprinted helper T cell subsets exhibit distinct functional phenotypes in central nervous system autoimmunity. Nat. Immunol. 22, 880–892 (2021).
Tantin, D. Oct transcription factors in development and stem cells: insights and mechanisms. Development 140, 2857–2866 (2013).
Ciofani, M. et al. A validated regulatory network for TH17 cell specification. Cell 151, 289–303 (2012).
Carr, T. M., Wheaton, J. D., Houtz, G. M. & Ciofani, M. JunB promotes TH17 cell identity and restrains alternative CD4+ T-cell programs during inflammation. Nat. Commun. 8, 301 (2017).
Bettelli, E. et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441, 235–238 (2006).
Bruder, D. et al. Neuropilin-1: a surface marker of regulatory T cells. Eur. J. Immunol. 34, 623–630 (2004).
Murai, M. et al. Interleukin 10 acts on regulatory T cells to maintain expression of the transcription factor Foxp3 and suppressive function in mice with colitis. Nat. Immunol. 10, 1178–1184 (2009).
Fragale, A. et al. IFN regulatory factor-1 negatively regulates CD4+ CD25+ regulatory T cell differentiation by repressing Foxp3 expression. J. Immunol. 181, 1673–1682 (2008).
Hu, X. & Ivashkiv, L. B. Cross-regulation of signaling pathways by interferon-γ: implications for immune responses and autoimmune diseases. Immunity 31, 539–550 (2009).
Trinchieri, G., Pflanz, S. & Kastelein, R. A. The IL-12 family of heterodimeric cytokines: new players in the regulation of T cell responses. Immunity 19, 641–644 (2003).
Wei, L. et al. Discrete roles of STAT4 and STAT6 transcription factors in tuning epigenetic modifications and transcription during T helper cell differentiation. Immunity 32, 840–851 (2010).
Granja, J. M. et al. ArchR is a scalable software package for integrative single-cell chromatin accessibility analysis. Nat. Genet. 53, 403–411 (2021).
Bravo Gonzalez-Blas, C. et al. cisTopic: cis-regulatory topic modeling on single-cell ATAC-seq data. Nat. Methods 16, 397–400 (2019).
Schep, A. N., Wu, B., Buenrostro, J. D. & Greenleaf, W. J. chromVAR: inferring transcription-factor-associated accessibility from single-cell epigenomic data. Nat. Methods 14, 975–978 (2017).
Moisan, J., Grenningloh, R., Bettelli, E., Oukka, M. & Ho, I. C. ETS-1 is a negative regulator of TH17 differentiation. J. Exp. Med. 204, 2825–2835 (2007).
Gilchrist, M. et al. Systems biology approaches identify ATF3 as a negative regulator of Toll-like receptor 4. Nature 441, 173–178 (2006).
De Nardo, D. et al. High-density lipoprotein mediates anti-inflammatory reprogramming of macrophages via the transcriptional regulator ATF3. Nat. Immunol. 15, 152–160 (2014).
Zhang, H. et al. An IL-27-driven transcriptional network identifies regulators of IL-10 expression across T helper cell subsets. Cell Rep. 33, 108433 (2020).
Filen, S. et al. Activating transcription factor 3 is a positive regulator of human IFNγ gene expression. J. Immunol. 184, 4990–4999 (2010).
Hudson, W. H. et al. Proliferating transitory T cells with an effector-like transcriptional signature emerge from PD-1+ stem-like CD8+ T cells during chronic infection. Immunity 51, 1043–1058 (2019).
Yao, C. et al. BACH2 enforces the transcriptional and epigenetic programs of stem-like CD8+ T cells. Nat. Immunol. 22, 370–380 (2021).
Bettelli, E. et al. Myelin oligodendrocyte glycoprotein-specific T cell receptor transgenic mice develop spontaneous autoimmune optic neuritis. J. Exp. Med. 197, 1073–1081 (2003).
Schnell, A. Stem-like T cells in cancer and autoimmunity. Immunol. Rev. https://doi.org/10.1111/imr.13356 (2024).
Roychoudhuri, R. et al. BACH2 represses effector programs to stabilize Treg-mediated immune homeostasis. Nature 498, 506–510 (2013).
Igarashi, K., Kurosaki, T. & Roychoudhuri, R. BACH transcription factors in innate and adaptive immunity. Nat. Rev. Immunol. 17, 437–450 (2017).
Grant, F. M. et al. BACH2 drives quiescence and maintenance of resting Treg cells to promote homeostasis and cancer immunosuppression. J. Exp. Med. 217, e20190711 (2020).
Mouri, K. et al. Prioritization of autoimmune disease-associated genetic variants that perturb regulatory element activity in T cells. Nat. Genet. 54, 603–612 (2022).
McAllister, K. et al. Identification of BACH2 and RAD51B as rheumatoid arthritis susceptibility loci in a meta-analysis of genome-wide data. Arthritis Rheum. 65, 3058–3062 (2013).
Perga, S. et al. Anti-inflammatory genes associated with multiple sclerosis: a gene expression study. J. Neuroimmunol. 279, 75–78 (2015).
Yang, L., Chen, S., Zhao, Q., Sun, Y. & Nie, H. The critical role of BACH2 in shaping the balance between CD4+ T cell subsets in immune-mediated diseases. Mediators Inflamm. 2019, 2609737 (2019).
International Multiple Sclerosis Genetics Consortium. Multiple sclerosis genomic map implicates peripheral immune cells and microglia in susceptibility. Science 365, eaav7188 (2019).
Guo, M. et al. Dissection of multiple sclerosis genetics identifies B and CD4 T cells as driver cell subsets. Genome Biol. 23, 127 (2022).
Javierre, B. M. et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell 167, 1369–1384 (2016).
Schmiedel, B. J. et al. Impact of genetic polymorphisms on human immune cell gene expression. Cell 175, 1701–1715 (2018).
Lee, Y. & Kuchroo, V. Defining the functional states of TH17 cells. F1000Res 4, 132 (2015).
Kim, E. H. et al. BACH2 regulates homeostasis of Foxp3+ regulatory T cells and protects against fatal lung disease in mice. J. Immunol. 192, 985–995 (2014).
Roychoudhuri, R. et al. BACH2 regulates CD8+ T cell differentiation by controlling access of AP-1 factors to enhancers. Nat. Immunol. 17, 851–860 (2016).
Herndler-Brandstetter, D. et al. KLRG1+ effector CD8+ T cells lose KLRG1, differentiate into all memory T cell lineages, and convey enhanced protective immunity. Immunity 48, 716–729 (2018).
Karmaus, P. W. F. et al. Metabolic heterogeneity underlies reciprocal fates of TH17 cell stemness and plasticity. Nature 565, 101–105 (2019).
Afzali, B. et al. BACH2 immunodeficiency illustrates an association between super-enhancers and haploinsufficiency. Nat. Immunol. 18, 813–823 (2017).
Vahedi, G. et al. Super-enhancers delineate disease-associated regulatory nodes in T cells. Nature 520, 558–562 (2015).
Aschenbrenner, D. et al. An immunoregulatory and tissue-residency program modulated by c-MAF in human TH17 cells. Nat. Immunol. 19, 1126–1136 (2018).
Brinkman, E. K., Chen, T., Amendola, M. & van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 42, e168 (2014).
Corces, M. R. et al. Lineage-specific and single-cell chromatin accessibility charts human hematopoiesis and leukemia evolution. Nat. Genet. 48, 1193–1203 (2016).
Picelli, S. et al. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 9, 171–181 (2014).
Kurtulus, S. et al. Checkpoint blockade immunotherapy induces dynamic changes in PD-1–CD8+ tumor-infiltrating T cells. Immunity 50, 181–194 (2019).
Singer, M. et al. A distinct gene module for dysfunction uncoupled from activation in tumor-infiltrating T cells. Cell 166, 1500–1511 (2016).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP–seq (MACS). Genome Biol. 9, R137 (2008).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
McLean, C. Y. et al. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 28, 495–501 (2010).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Li, B. et al. Cumulus provides cloud-based data analysis for large-scale single-cell and single-nucleus RNA-seq. Nat. Methods 17, 793–798 (2020).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902 (2019).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
McCarthy, D. J., Chen, Y. & Smyth, G. K. Differential expression analysis of multifactor RNA-seq experiments with respect to biological variation. Nucleic Acids Res. 40, 4288–4297 (2012).
DiSpirito, J. R. et al. Molecular diversification of regulatory T cells in nonlymphoid tissues. Sci. Immunol. 3, eaat5861 (2018).
Murtagh, F. & Legendre, P. Ward’s hierarchical agglomerative clustering method: which algorithms implement Ward’s criterion? J. Classification 31, 274–295 (2014).
Schnell, A., Thakore, P., Notarbartolo, S. & Sallusto, F. Chromatin-based mechanisms of diversification of regulatory and pro-inflammatory states reveal a role for BACH2 in TH17-based autoimmunity. Zenodo https://zenodo.org/records/11186874 (2024).
Acknowledgements
We thank the Broad Genomics Platform for help with bulk RNA-seq sample processing, L. Gaffney for graphical help in figure preparation and M. Collins for critical feedback on the manuscript. We thank all members of the laboratories of V.K.K. and A.R. for feedback. We thank J. Xia and H. Stroh for technical support. A. Schnell was supported by a German Academic Scholarship Foundation (Studienstiftung des Deutschen Volkes) PhD fellowship. P.I.T. was supported by a National Institutes of Health F32 Ruth L. Kirschstein Postdoctoral Fellowship (5F32AI138458). This work was supported by the National Institutes of Health (R01NS045937, R01NS30843, R01AI144166, P01AI073748, P01AI039671 and P01AI056299; V.K.K.) and the Klarman Cell Observatory and HHMI (A.R.). F.S. was supported by the Swiss National Science Foundation (189331). F.S. and the Institute for Research in Biomedicine are supported by the Helmut Horten Foundation. N.A.P. was supported, in part, by the National Multiple Sclerosis Society (JF-1808-32223 and RG-1707-28657).
Author information
Authors and Affiliations
Contributions
P.I.T., A. Schnell, V.K.K. and A.R. conceived the study. P.I.T., A. Schnell, O.R.-R., V.K.K. and A.R. designed the experiments and interpreted the results. P.I.T. and A. Schnell performed the in vitro T cell culture experiments with help from A. Singaraju. A. Schnell performed the in vivo mouse experiments with assistance from Y.H. P.I.T. and A. Schnell performed the ATAC-seq and RNA-seq experiments, with assistance from M.Z., E.C., S.Z., C.W., V. Singh, V. Sankar and O.R.-R. P.I.T. designed and performed the computational analysis, and L.H. performed the network analysis with assistance and guidance from A. Schnell, V.K.K. and A.R. R.K.K. and D.K. helped with flow cytometry and cell sorting. S.M. and J.D.B. helped with the scATAC-seq experiments. S.N. and F.S. helped with the acquisition of human T cell samples. N.A.P. provided the human BACH2 GWAS data. The manuscript was written by A. Schnell and P.I.T. and was edited by V.K.K. and A.R. with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
A.R. is a cofounder of and equity holder in Celsius Therapeutics, an equity holder in Immunitas and was a Scientific Advisory Board member of Thermo Fisher Scientific, Syros Pharmaceuticals, Neogene Therapeutics and Asimov until 31 July 2020. A.R. has been an employee of Genentech (member of the Roche Group) since August 2020. From 22 March 2021, P.I.T. has been an employee of Genentech. A.R. and P.I.T. have equity in Roche. From 19 October 2020, O.R.-R. has been an employee of Genentech and has equity in Roche. O.R.-R. is a co-inventor on patent applications filed by the Broad Institute for inventions related to single-cell genomics. She has given numerous lectures on the subject of single-cell genomics to a wide variety of audiences and, in some cases, has received remuneration to cover time and costs. V.K.K. is also a cofounder of, has an ownership interest in and is a member of the Scientific Advisory Board for Celsius Therapeutics, Tizona Therapeutics, Bicara Therapeutics and Larkspur Biosciences. The financial interests and conflicts of V.K.K. are managed by Brigham and Women’s Hospital and Partners Health Care system. None of these companies provided support for this work. J.D.B. holds patents related to ATAC-seq and is a Scientific Advisory Board member of Camp4 and seqWell. The other authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks Christina Zielinski and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: N. Bernard, in collaboration with the Nature Immunology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Bulk RNA-seq of in vitro differentiated Th17 cells.
(a) Gating strategy for FACS. Exemplary plot of in vitro differentiated npTh17 cells at 72 h sorting singlet viable CD4+ cells (all, black) and singlet viable CD4+ IL-17A-GFP+ cells (GFP+, green). (b) Bulk ATAC-seq of in vitro differentiated npTh17 and pTh17 cell populations with rest and restimulation. PCA of the first (x axis) and second (y axis) PCs of chromatin profiles (dots) of npTh17 (circles) and pTh17 (triangles) cell populations profiles at 72 h differentiation (pink), followed by 48 h resting phase (green), and after 48 h restimulation (blue). (c,d,e) ATAC-seq of in vitro cultured human Th17 cells. Representative example (c) of the cytokines produced by IFN-γ+IL-10– and IFN–γ– IL-10+ Th17 clone pools at Day 0 and Day 3 upon 5 h of PMA/Iono stimulation. PCA (d, first (x axis) and third (y axis) PCs) of ATAC-seq profiles (e, top 1,000 peaks from PC3, rows, -Z score of normalized counts) of in vitro cultured human IL-10− IFNγ+ (pink) or IL-10+ IFNγ- (blue) Th17 cells unstimulated (0 hr; circles and light grey) or re-stimulated with anti-CD3/-CD28 for 6 h (6 h; triangles and mid-grey) or 3 days (72 h, squares and dark grey). Selected genes associated with the ATAC-seq peaks are labeled on left. (f,g) Bulk RNA-seq of in vitro differentiated npTh17 and pTh17 cell populations. PCA (f) of cell profiles and expression (g, z-score of normalized counts, color bar) of the union of the top 200 differentially expressed genes (g, rows) at 24 h, 48 h, or 72 h (FDR < 0.05, rank ordered in C by fold change, 415 genes) in npTh17 (blue) and pTh17 (blue) cells populations from Il17aGFP reporter mice either after sorting for GFP+ cells (light color) or without sorting (all viable cells, dark color).
Extended Data Fig. 2 Comparison of in vitro and in vivo Th17 cell ATAC-seq peaks.
(a,b) Gene expression profiles of CNS-infiltrating Th17 cells. (a) PCA of bulk RNA-seq profiles from viable CD45+ TCRb+ CD4+ IL17-A-GFP+ dLN-derived (teal) and CNS-derived (pink) Th17 cell populations. dLN and CNS samples are matched for each biological replicate (shapes, n = 5). (b) Relative expression (z-score, color bar) of the 1,565 genes (rows) differentially expressed (FDR < 0.05, |log2(fold change|) between dLN- (teal) and CNS- (pink) derived Th17 cells (rows). (c) Chromatin accessibility (z-score of normalized counts, rows) in the top 5,000 DACRs (rows, FDR < 0.05) between dLN- (teal) and CNS- (pink) derived Th17 cell populations, ranked by ranked by log2(fold change). (d-h) GPR15 is associated with Th17 cell pathogenicity. (d, e) Expression (y axis, normalized counts by RNA-seq) of Gpr15 in different CD4+ T cell subsets differentiated in vitro (d, x axis; n = 4) and in Th17 cells from the dLN (n = 6) and CNS (n = 7) of EAE mice (e, x axis). (f) Clinical EAE score (y axis, mean with ± SEM) over time (x axis) in Gpr15-/- (blue) and WT (grey) mice. (g, h) Clinical EAE score (n = 5 Gpr15-/- and 4 WT mice) at day 18 (g, y axis, mean ± SEM) and frequency of IL-17A+ IFNγ+ CD4+ T cells in the CNS by flow cytometry (n = 4 mice) (h, y axis, mean ± SEM) in Gpr15-/- and WT mice. Unpaired two-tailed t-test. (i) Expression of signature cytokines in in vitro differentiated Th1 cells, npTh17 cells, pTh17 cells, and Treg cells. Illustrative flow cytometry plots are shown. (j, k) Gene expression profiles of CD4+ T cell subsets. (j) PCA of RNA-seq profiles of in vitro differentiated Th0 cells (yellow), npTh17 cells (blue), pTh17 cells (purple), Th1 cells (red), and Treg cells (green) at 72 h. (k) Relative expression (z-score) of the top 500 genes (rows) differentially expressed (FDR < 0.05) between any two cell-type conditions (1,556 genes total) in cells of each type (columns), with genes ordered by k-mean clustering (k = 8) into cell type specific clusters. Key genes are highlighted on the right.
Extended Data Fig. 3 ATAC-seq peaks across CD4+ T cell subsets.
(a) RNA expression level (left, y axis, normalized transcript counts) and enrichment of ATAC-seq signal over background (right, y axis, log fold) of Il10rb (chr16:91,420,666-91,445,000), Il9 (chr13:56,505,989-56,508,498), Maf (chr8:115,567,448-115,569,757), Il17a (chr1:20,727,418-20,762,267) loci in in each CD4+ T cell subset. Gene bodies are displayed on the bottom. In cases of distal peaks the associated gene is indicated (right: downstream). (b, c) Impact of Th1− and Treg-polarizing conditions on npTh17 and pTh17 cells. Fraction on IL-17A+ (b, left, y axis), IFNγ+ (b, right, y axis), FoxP3+ (c, left, y axis), and IL-10+ (c, right, y axis) cells out of CD4+ cells after npTh17 and pTh17 cells were in vitro differentiated for 3 days, rested (for 2 days) and restimulated (2 days) with the same cytokines (‘npTh17’, ‘pTh17’) or with IL-12 (b) or TGFβ (c) (n = 3 mice). Bona fide Th1 cells (b) and Tregs (c) were included as controls. Data are presented as mean ± SEM and an unpaired two-tailed t-test was performed.
Extended Data Fig. 4 Time course chromatin analysis of Th17 and Th1 differentiation.
(a) Expression of the marker cytokines at 48 h. Flow cytometry plots for expression of IL-17A (y axis) and IFNγ (x axis) at 48 h during the time course experiment. (b) Time and activation are captured by independent axes of variation (PCs) in both bulk ATAC- and RNA-seq. PCA plot of PC1 and PC2 of bulk ATAC-seq (left) and bulk RNA-seq (right) of naïve and in vitro differentiated npTh17 (blue), pTh17 (purple) and Th1 (red) cells at 0 h, 1 h, 6 h, 12 h, 20 h, and 48 h (grey scale). (c) Shared chromatin regions close during Th17 and Th1 differentiation. Venn diagram of chromatin regions that close by 48 h in npTh17 (blue), pTh17 (purple) or Th1 (red) cells. (d, e) Distribution of TSS enrichment (d, y axis) and number of fragments (e, y axis) for each scATAC-seq cluster (x axis) (as in Fig. 5b); n = 2 biological replicates per treatment group, npTh17, pTh17, or Th1. The box plots display the upper quartile value (upper bound of box), the median value (center line of box), and the lower quartile value (lower bound of box), as well as the upper extreme (top line) and lower extreme (bottom line).
Extended Data Fig. 5 ScATAC-seq and scRNA-seq of Th17 cells.
(a,b) ScATAC-seq profiles cluster by cell subset. (a) Fraction of cells (color bar) of each sample (columns) that are members of each scATAC-seq cluster (rows, defined as in Fig. 5b). (b) UMAP embedding of scATAC-seq profiles (as in Fig. 5b) colored by enrichment score of cluster markers (as in Fig. 5c). (c,d) ScATAC-seq profile clusters are enriched for distinct Th17 cell signatures. (c) Enrichment (color bar) of Th17 pathway signatures9 (rows) with genes associated with differentially accessible ATAC peaks in each scATAC-seq cluster (columns). (d) UMAP embedding of scATAC-seq profiles (as in Fig. 5b) colored by enrichment score of Th17 pathway signatures. (e,f) ScRNA-seq of in vitro differentiated npTh17, pTh17, and Th1 cells at 48 h. UMAP embedding of scRNA-seq profiles colored by treatment (e) or cluster assignment (f). (g) Differential peak-to-gene association of key Th17 effector genes in distinct scATAC-seq clusters. Relative signal (z-score, color bar) of peak-to-gene links for selected Th17 effector genes (columns) in each cluster (rows, as in Fig. 5d). (h,i) Variation in chromatin accessibility of npTh17 and pTh17 marker genes across scATAC-seq profiles. UMAP embedding of scATAC-seq profiles (as in Fig. 5b) colored by ATAC-signal as gene score (top) and predicted gene expression as gene integration (bottom) for selected marker genes of npTh17 (h) and pTh17 (i) cells.
Extended Data Fig. 6 ScATAC-seq topic modeling of Th17 and Th1 cells.
(a) Distinct and shared gene programs across Th17 and Th1 cells identified by topic modeling. UMAP embedding of scATAC-seq profiles (dots) colored by topic scores (normalized topic score, colorbar) for 30 of 35 topics (the remaining 5 topics are in Fig. 5f). (b) Chromatin peaks underlying key topics. Regions scores (colorbar) for each of the top 500 accessible chromatin peaks (columns) in five selected topics (rows, same topics as in Fig. 5f). Selected genes associated with peaks in each topic are listed on the right.
Extended Data Fig. 7 TF motif enrichment in scATAC-seq peaks.
(a) TF motifs enriched in accessible peaks in different cell subsets. Significance (-log10(Bonferoni adjusted p-value), hypergeometric test, y axis) and rank (x axis) of enrichment of motifs (dots) in npTh17- (left), pTh17- (middle), and Th1-specific (right) peaks. (b) Comprehensive inferred Th17 and Th1 regulatory network. Pearson correlation coefficients (red/blue color bar) between each TF (row) and each target (column) in the network. Targets were split into two categories based on whether they themselves are (right) or are not (left) TFs regulating other targets in the network. Color bars 1 and 2 indicate the TF and target modules and color bar 3 shows the cell type(s) where the interaction was detected (that is, the target gene was upregulated). (c) Inferred regulatory interactions of ATF3 with its predicted direct targets. Edge color: cell type(s) in which the association was detected; Edge line type: direction of association; Edge width: absolute value of the Pearson correlation coefficient of the expression level between ATF3 and target gene; Node color: TF or non-TF genes. (d) Transcription factor motif enrichment of DACR clusters. Significance (-log10(Bonferroni adjusted p-value), hypergeometric test, y axis), rank (x axis), and fold enrichment compared to background peak set (dot color) of enrichment of motifs (dots) in DACR clusters. (e,f) BATF induces and T-BET suppresses the Th17 program in npTh17 and pTh17 cells. Batf-/- (n = 5 mice) (e) or Tbx21-/- (n = 5 mice) (f) and wild-type (WT) (n = 3 mice) naïve CD4+ T cells were in vitro polarized to npTh17 (left) or pTh17 cells (right). Percent of IL-17A+ (y axis left, mean with ± SEM) and mean fluorescence intensity (MFI) of RORγt (y axis right, mean with ± SEM) of npTh17 and pTh17 cells by flow cytometry. NS= not significant, unpaired two-tailed t-test.
Extended Data Fig. 8 Validation of Bach2 CRISPR/Cas9 editing efficiency.
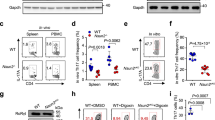
(a) Editing efficiency at the Bach2 locus. Editing efficiency (y axis; % loci with indels by Sanger sequencing) for a Bach2-targeting sgRNA (measured at the Bach2 locus) and for two sgRNAs targeting the GFP transgene locus (x axis). (b) Bulk RNA-seq of in vitro differentiated npTh17 and pTh17 Bach2 KO cells. RNA-seq normalized counts (z-score) for 863 genes (rows) that are differentially expressed (FDR < 0.05) between Bach2 KO (green) and control (grey) for pTh17 or npTh17 cells, and clustered by k-means clustering (k = 4). Relevant genes are listed on the left. (c) Differentially accessible loci in Bach2 KO are enriched for pathogenic Th17 signatures. Enrichment (x axis) of Th17 pathway signatures9,42,43 (y axis) in genes associated with the differential ATAC-peaks in Bach2 KO vs. control in npTh17 cells (purple, 1,118 peaks) and in pTh17 cells (blue, 627 peaks) (Supplementary Table 13). Signature origins: Stem-like CD101-TIM3− d45 or d1542, stem-like CD8+ T cells43, and others9. (d) Impact of Bach2 KO on chromatin accessibility in Treg and Th17 related loci. Global chromatin accessibility (y axis, average normalized reads) in npTh17 (blue) and pTh17 (purple) cell populations from Bach2 KO (light color) or control (dark color) cells in chromatin regions that are more accessible in Treg-specific clusters (2–4 in Fig. 3e) and in Th17-specific clusters (6,7 in Fig. 3e). (e, f) Mice were adoptively transferred with Bach2 KO (green) or control (grey) 2D2 cells. (e) Frequency (left) and number (right) of viable 2D2 cells in the CNS. (f) Frequency of 2D2 cells in the spleen (left) and CNS (right). Data are mean ± SEM. NS = Not significant, unpaired two-tailed t-test. n = 4 mice.
Extended Data Fig. 9 Bach2 overexpression in Th17 cells.
(a) Bach2 overexpression validation. Bach2 transcript expression (y axis, fold change by qPCR, mean ± SEM) in Bach2 overexpressing (Bach2 OE) and control pTh17 cells (x axis) (n = 4). Unpaired two-tailed t-test. (b,c) Impact of Bach2 OE on cell frequency, number and viability in vivo. Frequency (b, left) number (b, right) of transduced (GFP+) cells in the spleen and (c) frequency of viable GFP+ cells in the spleen (left) and CNS (right) in mice adoptively transferred with Bach2 OE (green) or control (grey) 2D2 cells. Data are mean ± SEM. NS = Not significant. unpaired two-tailed t-test. n = 5 mice. (d) Differentially expressed genes in Bach2 OE pTh17 2D2 cells. Significance (-log10(p-value), Wald’s test, y-axis) and effect size (log2(fold change), x axis) of differential expression of each gene between transduced cells (GFP+) isolated from the CNS of recipients of Bach2 OE or control pTh17 2D2 cells. Blue: FDR < 0.2. (e) BACH2 induces a stem-like Th17 cell program. Enrichment score (x axis) of the in vivo stem-like Th17 cell signature13 with genes associated with differentially accessible ATAC peaks in the Bach2 OE and Bach2 KO cells (y axis). (f) RNA expression level (y axis, normalized RNA counts) of stem-like genes in in vitro differentiated Bach2 OE (n = 4 mice) (left) and KO (n = 3 mice) (right) pTh17 cells (green) and matching controls (grey) (x axis). Unpaired two-tailed t-test. (g, h) Inferred regulatory interactions of BACH2 (g) or BACH2 and RBPJ (h) with their predicted direct targets. Edge color: cell type(s) in which the association was detected; Edge line type: direction of association; Edge width: absolute value of the Pearson correlation coefficient of the expression level between BACH2 and target gene; Node color: TF or non-TF genes. In (g) genes are highlighted in bold if they were validated using bulk Bach2 KO vs. WT RNA-seq data (log2(fold change) >0 for repressed genes and log2(fold change) <0 for activated genes).
Extended Data Fig. 10 Association of rs72928038 variant with increased BACH2 expression.
(a) Significance (-log10(p-value), hypergeometric test, dot size) of fold enrichment (x axis) over background, of TF motifs (y axis) rank ordered by fold enrichment (x axis) in ATAC-seq peaks differentially accessible between IL-10− IFNγ+ and IL-10+ IFNγ- Th17 cells at 24 h of activation. (b,c) Significance (–log10(p-value)) of association of each variant (dot) with MS risk in GWAS (x axis; from the discovery phase of IMSGC53) and as a BACH2 cis-eQTL (y axis, from the database of immune cell expression study [DICE]56) in CD4+ naïve (b) or CD4+ Treg (c) cells for variants in a region spanning 1MB up and downstream of rs72928038. Rs72928038 post-replication p = 8.38*10−29 and odds ratio for G allele was 0.866. Variants are colored by statistical significance in both MS GWAS and eQTL study (dark blue), only in MS GWAS (light blue), only in eQTL study (light red), or neither (light grey). P-values were determined by GWAS and eQTL analysis with the following statistical significance thresholds: MS GWAS: P < 5*10−8 (genome wide); eQTL: FDR < 10% (locus only test). (d) Tracks from top to bottom display: position in chromosome 6; reference sequence genes (truncated view); alignment of mouse genome (mm9) to human genome (hg19; https://vizhub.wustl.edu/public/hg19/weaver/hg19_mm9_axt.gz); evolutionary conservation in 46 vertebrate species; position of rs72928038; and promoter capture interaction loop between enhancer intronic region, overlapping rs72928038, and BACH2 promoter in CD4+ naïve cells55. All positions are in human genome version 19. (e, f) Distribution of gene expression (y axis, log2(transcripts per million) (log2(TPM)) for each rs72928038 genotype (x axis) in CD4+ naïve cells (e) and CD4+ Treg cells (f). P-values were determined by cis-eQTL analysis of rs72928038 and BACH256. N = replicates as published in Schmiedel et al.56 The box plots display the upper quartile value (upper bound of box), the median value (center line of box), and the lower quartile value (lower bound of box), as well as the upper extreme (top line) and lower extreme (bottom line).
Supplementary information
Supplementary Tables 1–15
Supplementary Table 1. Bulk ATAC-seq data from human IFNγ+IL-10– and IFNγ–IL-10+ TH17 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 2. Bulk ATAC-seq and RNA-seq signals from gene-to-peak correlations in npTH17 versus pTH17 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 3. Bulk ATAC-seq data from CNS-derived versus dLN-derived TH17 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 4. Bulk RNA-seq data from CNS-derived versus dLN-derived TH17 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 5. Gene set enrichment analysis for genes near differentially accessible peaks in cells collected from the CNS versus dLN during EAE (P values were calculated by permutation test followed by Benjamini–Hochberg FDR testing). Supplementary Table 6. Bulk ATAC-seq data from CD4+ T cell subsets. Supplementary Table 7. Bulk RNA-seq data from CD4+ T cell subsets. Supplementary Table 8. Bulk ATAC-seq time course of TH17 differentiation. Supplementary Table 9. Peak-to-gene links of scATAC-seq/RNA-seq of TH17 and TH1 cells (P values were calculated by t-test). Supplementary Table 10. TF regulators from integrated scATAC-seq and scRNA-seq data from TH17 and TH1 cells (P values were calculated by t-test). Supplementary Table 11. Bulk RNA-seq data from Bach2-KO TH17 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 12. Bulk ATAC-seq data from Bach2-KO TH17 and TH1 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 13. Gene set enrichment analysis for genes near differentially accessible peaks in Bach2-KO versus control TH17 cells (P values were calculated by permutation test followed by Benjamini–Hochberg FDR testing). Supplementary Table 14. Bulk ATAC-seq data from Bach2-OE TH17 and TH1 cells (significance was determined by Wald’s test followed by Benjamini–Hochberg FDR testing). Supplementary Table 15. Gene set enrichment analysis of differentially expressed genes in Bach2-OE versus control cells collected from the CNS during EAE (P values were calculated by permutation test followed by Benjamini–Hochberg FDR testing).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Thakore, P.I., Schnell, A., Huang, L. et al. BACH2 regulates diversification of regulatory and proinflammatory chromatin states in TH17 cells. Nat Immunol 25, 1395–1410 (2024). https://doi.org/10.1038/s41590-024-01901-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-024-01901-1
- Springer Nature America, Inc.