

Abstract
Global warming is markedly changing diverse coral reef ecosystems through an increasing frequency and magnitude of mass bleaching events1,2,3. How local impacts scale up across affected regions depends on numerous factors, including patchiness in coral mortality, metabolic effects of extreme temperatures on populations of reef-dwelling species4 and interactions between taxa. Here we use data from before and after the 2016 mass bleaching event to evaluate ecological changes in corals, algae, fishes and mobile invertebrates at 186 sites along the full latitudinal span of the Great Barrier Reef and western Coral Sea. One year after the bleaching event, reductions in live coral cover of up to 51% were observed on surveyed reefs that experienced extreme temperatures; however, regional patterns of coral mortality were patchy. Consistent declines in coral-feeding fishes were evident at the most heavily affected reefs, whereas few other short-term responses of reef fishes and invertebrates could be attributed directly to changes in coral cover. Nevertheless, substantial region-wide ecological changes occurred that were mostly independent of coral loss, and instead appeared to be linked directly to sea temperatures. Community-wide trophic restructuring was evident, with weakening of strong pre-existing latitudinal gradients in the diversity of fishes, invertebrates and their functional groups. In particular, fishes that scrape algae from reef surfaces, which are considered to be important for recovery after bleaching2, declined on northern reefs, whereas other herbivorous groups increased on southern reefs. The full impact of the 2016 bleaching event may not be realized until dead corals erode during the next decade5,6. However, our short-term observations suggest that the recovery processes, and the ultimate scale of impact, are affected by functional changes in communities, which in turn depend on the thermal affinities of local reef-associated fauna. Such changes will vary geographically, and may be particularly acute at locations where many fishes and invertebrates are close to their thermal distribution limits7.
Similar content being viewed by others


Main
The 2016 mass bleaching event affected coral reefs world-wide, with catastrophic impacts reported in the Red Sea, central Indian Ocean, across the Pacific Ocean and in the Caribbean3,8,9. The Australian Great Barrier Reef (GBR), the largest coral reef system in the world, experienced the warmest temperatures on record for the region. An estimated 91.1% of reefs along the GBR experienced some bleaching3, resulting in an estimated loss of approximately 30% of live coral cover over the following six months10. The event was thus comparable to the 1998 mass bleaching event in the Indian Ocean in terms of reported impacts on corals2,11. We surveyed 186 reef sites along the GBR and at less-studied isolated reefs in the Coral Sea before and after the 2016 bleaching event, and here we report reef- and regional-scale effects of the extreme thermal anomaly and loss of coral cover on the rich reef-associated fish and mobile invertebrate fauna. At each site, globally standardized Reef Life Survey census methods12 were used to quantify changes to coral cover, reef fishes and mobile macroinvertebrates at multiple depths (overall mean, 6.7 m; range, 0.8–17.0 m). ‘Before’ data were obtained between 2010 and 2015, and ‘after’ data were obtained 8–12 months after bleaching.
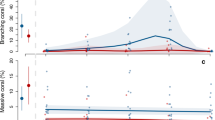
As reported elsewhere10, decreases in live hard coral cover were widespread (Fig. 1), although we found that the regional pattern was more spatially heterogeneous than previously described, when field surveys were standardized amongst shallow reef crest habitat10. Forty-four of the 186 surveyed sites experienced absolute declines in live coral cover that exceeded 10% (up to 51% loss for one site at Osprey Reef), with the northern Coral Sea reefs suffering the most consistent losses (Fig. 1a, b). The magnitude of coral-cover change was related to the local sea temperature anomalies (Fig. 1d and Extended Data Fig. 1), but coral loss varied considerably, and not all reefs in regions that experienced the greatest temperature anomalies experienced losses in live coral cover. In some cases, such as the central Coral Sea reefs, a history of cyclone damage meant that there was relatively little coral to lose. Thus, geographical patterns in pre-bleaching cover had a critical role in the realized effects of bleaching on corals (Fig. 1d). Coral-cover losses of the greatest magnitude occurred in disparate locations, including in the northern Coral Sea (Boot and Osprey Reefs; mean, 15% absolute cover loss, or approximately 40% of the pre-bleaching live coral cover), and the southern GBR (most southerly Swain Reefs; 28% loss, or 100% of pre-bleaching cover). The northern reefs in the GBR experienced the most extensive bleaching of those surveyed during the 2016 event3, but not all of the reefs in that area suffered the extreme rates of live coral-cover loss that were observed more generally10 (Fig. 1a, b). The fate of bleached corals can vary considerably13,14, and a reasonable proportion of corals on some of these reefs must have regained their zooxanthellae and survived the bleaching event. Algal cover substantially increased across the majority of reefs that experienced coral declines (Fig. 1c and Extended Data Fig. 2).

a–c, Reefs in the Coral Sea showed relatively consistent losses of live corals (a, b) and gains in algal cover (c) in the north, whereas changes along the GBR were highly patchy. Absolute changes in live coral cover are mapped for individual sites (n = 186), with aggregation of sites at the reef scale shown as plusses (n = 53). d, Coral-cover loss was related to the local heat anomaly from January to March 2016 regardless of depth, an effect that increased in strength according to pre-bleaching cover of live corals. Average pre-bleaching cover for the region was 26% (middle), whereas low and high (left and right) are shown for ±1 s.d. (19%) from average pre-bleaching live coral cover. Effects in d are from Bayesian mixed-effects models, with shading representing 95% credible intervals.
Not all coral declines that were observed during the study could be assumed to be solely due to the bleaching event (other disturbances, such as cyclones, may have also had impacts on corals at particular locations; see Methods). To investigate the effects on reef fauna that could be most clearly attributable to the bleaching event, we quantified changes on a subset of reefs that experienced extreme heating and substantial live coral-cover loss (see Methods for criteria). On these reefs, the abundance of coral-eating fishes (corallivores) consistently declined, and declines in local fish species richness were also common (Extended Data Fig. 3). Such changes have previously been observed as rapid responses to coral bleaching events5,15,16, and are clearly a concerning form of reef-scale biodiversity loss. These changes were not observed on a subset of comparison reefs that also experienced extreme heating, but that did not experience an observable loss of live coral cover (Extended Data Fig. 3). Other previously reported short-term effects of bleaching, such as increased herbivore abundance15 in response to a boom in algal resources5,16, occurred on some study reefs, but were not consistent features of those reefs with the clearest impacts on coral cover attributable to bleaching (Extended Data Fig. 3).
Coherent patterns of ecological change were evident when assessing regional-scale trends between survey periods across the full range of sites surveyed. The latitudinal gradient in local species richness of mobile fauna17 declined in slope through a combination of decreased local fish richness on northern reefs and markedly increased richness of macroinvertebrates and small cryptic fishes on southern reefs (Fig. 2 and Extended Data Fig. 4). The structure of fish communities on southern reefs became more similar to those in the north (Extended Data Fig. 5), a broad-scale homogenization that resulted in a slight decline in the overall number of fish species recorded across all surveys (from 532 to 494). Invertebrate communities also changed considerably between survey periods (Fig. 2 and Extended Data Fig. 4). This was characterized most clearly by sea urchins being found less frequently on northern reefs and in increased abundance on southern reefs after the bleaching event.

a–d, Plots show local richness (log scale). e, Plot shows sea urchin presence (log-odds). f–l, Plots show biomass in g per 500 m2 (log scale). Latitudinal trends are median effect estimates from Bayesian generalized linear mixed-effects models (n = 233 site-by-depth-category combinations). Shaded regions show the marginal 95% credible intervals, and asterisks indicate those metrics for which the term for the change in latitudinal slopes (the interaction between latitudinal and time period effects) has 95% credible intervals that do not overlap zero (model effect sizes with credible intervals for all predictors are shown in Extended Data Fig. 4). y axes are on the link scale (log for Poisson and normal, and logit for sea urchins).
A key outcome of these changes was the regional alteration to the functional structure of reef communities, with potentially important consequences for the recovery of affected reefs. Functional richness (represented by the number of unique functional trait combinations comprised by fishes and invertebrates on each survey) increased on southern reefs, where the potential for local herbivory also increased through herbivorous fish biomass gains (Fig. 2 and Extended Data Figs. 4, 6) and patchy gains in the abundance of sea urchins. By contrast, the frequency of occurrence and biomass of fishes that scrape algae and microscopic autotrophs off coral–rock surfaces (scraping herbivores), and the frequency of sea urchins declined on northern reefs, whereas the biomass of plankton-feeding fishes increased (Figs. 2, 3 and Extended Data Fig. 4).

a, Reefs north of 12° S latitude. Corallivores and scraping herbivores, the coolest-affinity trophic groups (lower STI values, on average), declined in frequency, while excavators (higher STI values, on average) increased in frequency on transects at northern reefs. n = 321 species in GBR, 301 species in Coral Sea. b, Reefs south of 19° S latitude. Excavators became more common on transects at southern reefs in the Coral Sea. n = 320 species in GBR, 305 species in Coral Sea. Points are means of species in each trophic group, shown separately for species recorded in the GBR (blue) and Coral Sea (orange). Data are mean ± s.e.m.
Most of these rapid, regional-scale ecological changes could not be linked to coral loss (Extended Data Fig. 4), and so cannot be assumed to be indirect effects of the bleaching event (or any other causes of coral degradation during the study). Some of these changes could nevertheless result from changes in the local composition and community structure of corals and algae, independently of the total amount of coral loss, but the spatial footprint of changes in the fishes and the invertebrates suggests that at least some of these changes were independent of changes in habitat. The consistency of ecological change along the latitudinal gradient differs from the heterogeneous patterns of the changes in coral and algal cover, particularly along the GBR, whereas the southern Coral Sea reefs showed very clear ecological changes, despite largely escaping bleaching. The loss of large predatory fishes in remote locations, such as in the northern GBR and on some reefs in the southern Coral Sea (Extended Data Fig. 6), could potentially be associated with expansion of the fishing footprint, but this needs further investigation. Changes in fishing pressure are unlikely to have resulted in most of the other coherent regional scale patterns of changes in the communities, because few herbivorous fishes, cryptic fishes and reef-dwelling invertebrates are targeted by fishers in this region.
Another potential explanation for the rapid restructuring of communities relates to more direct effects of region-wide anomalously warm temperatures and altered currents on the local occupancy patterns and abundance of different species18. Marine heat waves and short-term temperature variations have been shown to markedly affect temperate rocky reef communities19, but have not been well-investigated on coral reefs. The sea temperatures that were experienced during the bleaching event (up to 32 °C in the northern GBR18) exceeded those at the warm limits of the distributions for the majority of reef fishes that were recorded in the region7, and many species on northern reefs probably experienced thermal stress.
We used species temperature index (STI) values for fish species recorded during surveys to investigate the possibility that reduced species richness and altered trophic structure on the warmer northern reefs were due to disproportionate effects on species with an affinity for relatively cooler seas. STI values are derived from the realized thermal distributions of species across their entire range7,20, and provide a nuanced and continuous measure of the ocean climate on which the distribution of each species is centred. On average, patterns of change in the frequency of occurrence of species in each trophic group were positively related to their STI values (Fig. 3). Specifically, those species that declined between surveys across northern reefs tended to be corallivores and scraping herbivores with distributions in relatively cooler waters; a pattern that was consistent for fish communities along both the GBR and Coral Sea, which have different biogeographical affinities21 (Extended Data Fig. 5). Fishes that feed by excavating the coral–rock surface tended to have the warmest affinities (that is, higher STI values), and became more common in surveys in both the north and south (Fig. 3), although the increased frequency in the north did not translate to an increase in local biomass (Fig. 2 and Extended Data Fig. 4).
A bias in thermal affinities of reef fishes related to their trophic group has not previously been investigated in detail, and the generality of this phenomenon is unknown. In this case, the pattern was characterized by high variability (Fig. 3), and becomes increasingly influenced by excavators at the scale of the full GBR. The opposite situation may occur on temperate reefs, where herbivorous fishes have warmer STIs than other trophic groups22. Further investigation is needed to determine whether biases in STIs of trophic groups are idiosyncratic and location-dependent, or whether coherent geographical patterns emerge for particular trophic groups. The decreased frequency of occurrence of corallivores at northern reefs could also be related to coral mortality, with this effect potentially confounded by inferred effects of thermal stress (or other causes that were not investigated).
Ecological change on southern reefs included an increasing similarity of fish community structure to that on northern reefs (Extended Data Fig. 5), which is consistent with a potential influence of warmer temperatures, but could also result from altered currents and possible enhanced fish recruitment of northern species in the south. No clear signal of an influx of northern species recruits was evident, however, as the local richness of juveniles was no greater after the bleaching event than before (Extended Data Fig. 7). Instead, the majority of positive changes in the south related to taxa of relatively small adult body size—both invertebrates and cryptic fishes. These could be more sensitive to temperature changes and/or capable of increases in local population size more rapidly and/or could experience rapid numerical or behavioural release if predation pressure was reduced. Although less probable, release from predation may have resulted from minor decreases in the frequency of predatory fishes in the Coral Sea (Extended Data Fig. 6) and benthic invertebrate consumers in the GBR (Fig. 3).
Our broad-scale field surveys did not allow a definitive test of causation for the rapid regional ecological changes that were observed. Regardless of the causes, however, a critical feature is that the short-term effects of the bleaching may have been masked in some cases. For example, we observed an increase in fish species richness on a reef in the Swains area, despite concurrent coral devastation (albeit highly localized in a region that was otherwise little affected by bleaching3). Likewise, at the regional scale, local fish species richness increased on 40% of the surveyed reefs, despite mass bleaching, net coral loss and an overall decline in regional species richness. Such trends appear remarkable, given that a reduction in fish species richness has been amongst the most consistently and rapidly observed local ecological responses to coral loss observed in previous studies6,16.
The observed regional-scale reshuffling and trophic reorganization appear to be extremely rapid, observable less than one year after the bleaching event. Rapid changes have previously been noted, such as increasing densities of herbivores15, and have been hypothesized to be due to redistribution on reefs23 rather than to a demographic response5. A substantial proportion of pre-bleaching surveys were undertaken in 2013 (ranging from 2010 to 2015), and many of the observed changes could have resulted from a number of consecutive warm years, rather than the single 2016 bleaching event. In addition to the 2016 event, the study period included two of the next nine warmest years on record for the GBR region (http://www.bom.gov.au/climate/change/; accessed September 2017). The observed patterns may thus in part represent accumulated responses over multiple exceptionally warm years, and could provide valuable signs of the potential trajectory of ecosystem change for a warmer future with increasingly prevalent extreme events24.
Our observations of ecological change over an extreme heating event, with ecosystem consequences that are at least in part independent of coral mortality, may help to explain a lack of consistency among responses to bleaching observed in prior studies. For example, variability has previously been noted in herbivore responses25, despite relatively consistent increases in algal resources following coral death23. This response is of critical importance, as the biomass of herbivorous fishes can be highly influential in determining the recovery trajectories of bleached reefs2. Scraping herbivores are considered to be particularly important for supporting reef recovery26, and this group declined on northern reefs in our study. Whether losses of scraping herbivorous fishes in the northern GBR and Coral Sea will affect recovery of some of the most impacted reefs in the region remains an important question.
Ecosystem impacts of coral loss are likely to increase during the next decade in the GBR and Coral Sea if widespread erosion of dead corals occurs1,5,6,15. The extent to which the 2016 mass bleaching event proves ecologically catastrophic remains uncertain, as does the sum of accumulated effects from multiple bleaching events (as highlighted elsewhere3,24,27). However, rapid local recovery may occur on some reefs28. Either way, the trajectories of bleached reefs will be greatly influenced by the new community structures that we observed during a critical stage of reef recovery, and are thus inextricably linked with warming-related reshuffling of reef communities.
Overall, our results highlight the need for managers and researchers to consider broad spatial and temporal responses to the marine heating events amongst fishes and other biota, beyond the more readily observable impacts on coral habitat29. For example, potential ecological consequences of the changes that were observed in the northern GBR and Coral Sea could be exacerbated if herbivorous fishes were targeted by fisheries in these regions, whereas equivalent herbivore exploitation may not be an urgent management concern in locations where gains in herbivores occur (such as the southern GBR in our study). Likewise, functional changes in fish and invertebrate communities driven by extreme events may either complement or work against efforts to save reefs through restoration and assisted evolution of corals. Geographical location has been recognized as an important input into conservation planning and management from the perspective of considering patterns in ocean thermal regimes30. Our study highlights how location can additionally be important from the perspective of thermal affinities of community members. Accounting for the realized thermal niches of species in key functional groups may allow managers to more explicitly consider the trade-off between managing areas in which more species and functional groups are vulnerable to warming events, versus those in which fewer negative effects are expected. The former could potentially prolong local persistence of species and ecological stability by removing extractive pressures, and the latter may provide important reference areas for determining the importance of novel ecological interactions in shaping future reef ecosystems.
Methods
Data reporting
No statistical methods were used to predetermine sample size. The experiments were not randomized and the investigators were not blinded to allocation during experiments and outcome assessment.
Survey methods
Standardized data were obtained on fishes, mobile invertebrates, coral and algae along 768 50-m underwater transects by trained scientific and recreational divers who participated in the citizen science Reef Life Survey (RLS) program. Full details of census methods are provided elsewhere12,31,32, and an online methods manual (https://reeflifesurvey.com) describes all data-collection methods. Data quality and training of divers have previously been described12,33. All observed fish species were counted in duplicate 5-m-wide transect blocks and aggregated as densities per 500-m2 transect, and cryptic fishes and mobile invertebrates >2.5 cm total length in duplicate 1-m-wide transect blocks (aggregated to 100-m2 transect area) on the same transect lines. Fish length and abundance estimates were converted to biomass using species-specific length–weight coefficients obtained from FishBase (https://fishbase.org), as used in previous studies with the RLS data34,35. Invertebrate classes used for this study were Asteroidea, Cephalopoda, Crinoidea, Echinoidea, Gastropoda, Holothuroidea and Malacostraca. All individuals from these classes exceeding 2.5 cm total length were included in richness estimates for invertebrates, and in functional richness analyses.
Photoquadrats were taken vertically downward of the substrate every 2.5 m along each of the same transect lines, and later scored using a grid overlay of 5 points per image, 100 points per transect. Categories of benthic cover scored were from a set of 50 morphological and functional groups of algae and corals (Extended Data Table 1), as previously described32 and aligning with the standard Australian hierarchical benthic classification scheme36. Analyses undertaken for this study were based on the sum of all live hard coral categories (that is, percentage of live hard coral per transect), and the sum of all algal categories (percentage of algal cover per transect), with categories listed in Extended Data Table 1.
Survey design
Matching before–after bleaching surveys were undertaken at 186 GPS-referenced sites at 53 reefs (see Fig. 1 for distribution of reefs; mean = 3.5 sites per reef) along the full length of the GBR and western Coral Sea region within the Australian Exclusive Economic Zone. At each site, multiple surveys (mean = 2.1 transects per site) were undertaken at different depths, with transects laid along a depth contour. Depths were binned (see ‘Covariates’), such that the site-by-depth bin was the level of replication, making 233 matching site-by-depth replicates surveyed both before and after the bleaching event.
Different divers often surveyed the fishes and the invertebrates along the same transect line. Pre-bleaching surveys were mostly undertaken from a survey cruise along the entire GBR and Coral Sea in 2015 (42% of pre-bleaching surveys) and a previous survey cruise through the GBR and Coral Sea in 2013 (39% of pre-bleaching surveys). Additional ‘before data’ (19%) were collected at Lizard Island, Great Keppel Island and the Whitsundays in 2010, and some sites in the central Coral Sea and GBR in 2012. All post-bleaching data were collected during a survey cruise through the entire region from November 2016 to March 2017. No strong biases were apparent in the interval between pre- and post-bleaching surveys along the latitudinal gradient or locations experiencing different heating anomalies (Extended Data Fig. 8).
Seven of the 10 divers who undertook pre-bleaching surveys also undertook post-bleaching surveys, and G.J.E. and R.D.S.-S. together undertook 45% of all fish surveys (and led 85% of survey voyages before and 92% after the bleaching event). There was thus a substantial element of consistency in divers during the study. To explore the effect of different divers undertaking surveys at different times, however, we reran the models for Fig. 2 and Extended Data Fig. 4 with ‘diver’ included as a random effect. This resulted in no changes in the effect sizes or conclusions. Therefore, results are presented for models without the diver effect, so that marginal uncertainty intervals include site-to-site variation but not observer variation.
Species traits
All fishes and invertebrates were allocated into one of the following trophic groups: corallivores, scraping herbivores, benthic invertivores, algal farmers, browsing herbivores, omnivores, planktivores, higher carnivores, excavators, detritivores, suspension feeders and cleaners. Additional traits used for calculation of functional richness were: maximum body size (included as 10-cm bins up to 50 cm, and all species which grow to >50 cm binned together), and water column position (benthic, demersal, pelagic site-attached and pelagic non-site-attached). All traits were taken from a previously published dataset37. Functional richness was calculated as the richness of functional entities per 50-m transect, in which all species with the same combination of trait levels for those three traits were considered functionally equivalent.
STI values were taken for each species from a previously published dataset20, and represent the midpoint between the 5th and 95th percentile of local mean sea-surface temperature values from all occurrence locations of the species. It thus represents the centre of each species’ range when expressed as a range of sea temperatures experienced across its distribution, and provides a nuanced means of ordering species by their preferences for warmer or cooler environments. Full details, including discussion of strengths and weaknesses, are provided in previous publications7,20.
Covariates
The mean depth contour of each reef transect was recorded by divers during surveys, with surveys then allocated into three depth bins (<4 m, 4–10 m and >10 m). For any before–after comparisons, we first obtained the mean values of univariate responses taken from among all transects within each depth bin at a given site (that is, site-by-depth bin combinations). This gave 233 site-by-depth combinations, with a mean of 76 sites and 35.3 reefs per depth class. For each site, we also applied a four-level categorical measure for wave exposure: (1) sheltered, with only wind waves from non-prevailing direction; (2) wind-generated waves from the prevailing direction; (3) exposed to ocean swells, either indirectly with exposure to prevailing winds, or directly but sheltered from prevailing winds; or (4) exposed to open ocean swell from prevailing direction. There was a mean of 62 sites and 24 reefs per exposure category. Reef habitat categories are often used for ecological studies of coral reefs (for example, slope, crest, flat and lagoon), but delineation between similar or adjacent habitats can sometimes be difficult. Instead of making these delineations for our survey sites, we considered that these two environmental axes of wave exposure and depth together appropriately capture the important variation between such reef habitat classifications with respect to their importance in describing potential for bleaching2.
Sea-surface temperature anomalies used in analyses relating coral-cover change to degree heating days (DHD) was obtained from the ReefTemp Next Generation38. Fine scale anomalies for the period of January–March 2016 were matched to survey sites.
Analysis of coral- and algal-cover change
We modelled the response of change in coral and algal cover as a function of DHD using a Bayesian mixed-effects model (n = 211 site-depth combinations where benthic-cover data were available). Additional fixed covariates included the depth of survey, the four wave exposure categories, a factor for whether the survey was in the GBR or the Coral Sea, the initial cover of corals or algae, an interaction between DHD and depth and an interaction between DHD and initial cover. We included a random effect for reef. We did not include a random effect for sites nested within reefs because only 36 had measurements at more than one depth across both time periods (before and after bleaching). Change in coral and algal cover was modelled with Gaussian errors and standard model checks confirmed that this assumption was appropriate. We scaled the variance of the model by the number of years between before and after surveys (maximum = 7 years, mean = 3.3 years), because we expect greater variance in the measured change in coral cover when those measurements were taken a longer time apart. We compared models with and without the variance scaling using the widely applicable information criteria (WAIC)39,40. The WAIC indicated that for the coral-cover model with variance scaling provided an enhanced fit to the data (1,658 versus 1,721), whereas for the algae-cover model the unscaled model had an enhanced fit (1,801 versus 1,806), so we present results from these best models. However, the estimated effects of the covariates were nearly identical regardless of model used in both cases.
We present the median estimated effects of DHD on coral and algal cover in Extended Data Fig. 1, and credible intervals are 95% quantiles. We also predict median change and the 95% marginal credible intervals for change in coral and algal cover across the range of DHD for each depth category values for low wave exposure reefs in the GBR (Fig. 1d). Credible intervals for predictions were integrated across all random effects, so they should be interpreted as effect sizes relative to variation across reefs. For the coral model, there was a strong interaction effect of initial coral cover with DHD, so we separately plotted predicted effects for the mean initial coral cover and ±1 s.d. in initial coral cover.
We fitted the Bayesian mixed-effects models using the INLA framework41 implemented in the R programming language42 using the INLA R package (version 17.06.20; http://r-inla.org, accessed 4 October 2017). The prior for the precision on the random effect used the log-gamma prior with shape = 1 and rate = 1 × 10−5, although use of other standard priors did not change the results. Priors for fixed effects had mean = 0 and precision = 0.001.
Mapped coral change values in Fig. 1a represent absolute change in live hard coral cover at each site, with the change values interpolated using an inverse-distance weighting and a buffer of 50 km applied from around each reef surveyed (implemented with the gstat package in the R program43). Symbols on the map thus represent the locations of the reefs, although coral change values come from the aggregation of smaller scale data at individual sites within reefs.
Comparison of bleaching-impacted and unaffected reefs
To isolate ecological impacts most likely arising from bleaching-associated coral loss, we used the following criteria to define ‘bleaching-impacted’ reefs: (1) pre-bleaching live hard coral cover >20% on average (across all transects at the reef). This meant that the starting community was more likely one to be comprised of coral-associated fish and invertebrate species; (2) loss of live coral cover >40% of pre-bleaching values, on average; and (3) experienced more than 40 DHD. These criteria were collectively used as a means to show the maximum likely impact of the loss of coral from bleaching, by ensuring there was adequate coral cover to start with (criterion 1), and that coral losses were at least typical of the mortalities observed in other studies11 (criterion 2), while providing some confidence that observed coral loss was most likely attributable to bleaching (criterion 3). We cannot be certain about the latter (see comments below about other potential impacts on coral during the study period), but 40 DHDs well exceeds the threshold for bleaching as identified previously3 for this same bleaching event. Reefs defined as ‘bleaching-impacted’ were widely dispersed along the GBR and Coral Sea (Extended Data Fig. 3).
To provide an objective contrast with these reefs, we also selected reefs that were clearly unaffected by bleaching. We used the same criteria as above, but instead of losing at least 40% of live coral cover, we selected only those that experienced >1% mean gain in live hard corals on average (using the mean percentage of coral-cover change, rather than mean pre-bleaching minus mean post-bleaching cover). For all bleached and unaffected reefs, we examined responses in key metrics of the coral, fish and invertebrate communities, as shown in Extended Data Fig. 3.
Regional community-structure change
Non-metric multidimensional scaling was undertaken separately on reef fish and invertebrate community data to show broad regional change in community structure and visualize consistencies in the direction of community change among regions between before and after surveys (Extended Data Fig. 5). Mean biomass per fish species (kg per 500 m2) and mean abundance per mobile invertebrate species (individuals per 100 m2) were calculated separately across all surveys within each 2° latitudinal band for the GBR and Coral Sea. Biomass and abundance data were log-transformed and Bray–Curtis dissimilarity matrices used for ordination of each. The analysis was undertaken in PRIMER44, with symbols subsequently colour-coded for data collected before and after the bleaching event, and labels to indicate GBR and Coral Sea regions.
Analysis of regional-scale ecological changes
We analysed the response of nine fish and invertebrate metrics to bleaching using Bayesian generalized linear mixed-effects models (GLMMs). Each metric value on each survey was modelled with covariates for latitude, depth, coral cover, protection status (no-take ‘green’ zone versus all other zone types), GBR or coral sea, wave exposure, time (before or after the bleaching event) and an interaction between time and latitude. The interaction was included to allow for the possibility that latitudinal gradients in each metric changed from before to after the bleaching event. We included random effects for reefs and sites within reefs.
We chose error distributions appropriate for each metric. These were: Poisson with a log-link for the richness metrics, log-normal with an identity link for the biomass metrics, and binomial with a logit link for urchin presence. We added 0.5 to the logged biomass data so that zero values were not excluded. Checks of residuals confirmed that a log-normal distribution was appropriate for the biomass data. Rootograms45 and Dunn–Smyth residuals46 were used to confirm the count models were fitted appropriately.
We used the INLA framework to fit the Bayesian GLMMs, using the same settings as for the coral change model. We give effect sizes as median effects of each covariate in Fig. 2 and Extended Data Fig. 4 with 95% credible intervals. Credible intervals that did not overlap zero in Extended Data Fig. 4 are identified by asterisks. We also predict metrics across the latitudinal gradient before and after the bleaching event with marginal 95% credible intervals. Predictions across latitude were made for a reef of <4-m depth, with the mean level of hard coral cover, inside a protected area with low wave exposure and for the GBR. Thus positive and negative effects in Extended Data Fig. 4 can be interpreted in relation to these levels of the relevant covariates. Choosing other covariate values for the baseline would affect the magnitude of the patterns but not the overall trend.
Possible recruitment events
We tested whether patterns in richness before and after bleaching could be related to a coincident fish recruitment event. We analysed the mean richness of juvenile fishes per reef (29 reefs in total from extreme north and south) as a function of three binary covariates: before versus after bleaching, Coral Sea versus GBR and north versus south, using a linear model, implemented in the INLA framework41 from the R programming language42. Juveniles were defined as any individuals that were 10 cm or less, for species that exceed 12.5 cm in maximum size. No significant change in the richness of juveniles was evident before and after bleaching (mean difference = 1.70 with lower and upper 95% credible intervals of –0.6 and 4.0), with the distribution of data shown in Extended Data Fig. 7.
Other potential effects on results
Few trends in fishes and invertebrates were related to changes in coral cover when considered at the scale of the whole study region, and primary study conclusions do not rest on the assumption that all observed coral mortality was driven by the 2016 bleaching event. Cyclones, crown-of-thorns starfish, and pollution and sediment from riverine outputs are other potential impacts on corals across the region. We checked the database of past tropical cyclone tracks on the Bureau of Meteorology website (http://www.bom.gov.au/cyclone/history/index.shtml, accessed 7 April 2018) for intersection of cyclone tracks with our survey sites. Surveys were completed before cyclone Debbie (2017), and the only surveys done before cyclone Yasi (2011) (Lizard Island, Port Douglas, Whitsundays, Keppel) were in areas outside of the destructive path of this cyclone. However, cyclone Ita was reported to have impacts on corals in the Lizard Island area during the study period47, and there is a possibility that other smaller cyclones caused highly localized impacts. Thus, caution is required in ruling out cyclone damage as contributing to coral-cover changes observed in some locations.
We cannot be certain that crown-of-thorns starfish did not affect coral cover at our sites in between surveys, but these are also recorded on the surveys of mobile invertebrates and were found in extremely low densities (mean = 1.4 individuals per 50 m2 when found, only at 15 sites). It is not impossible that a wave of crown-of-thorns starfish came through and reduced live coral cover at a small number of sites, but such effects at this very small number of sites would unlikely have a detectable impact on results or conclusions of the study. Likewise, pollution and sediment from riverine sources could not have been responsible for any changes in the Coral Sea (>250 km offshore), and would be unlikely to have impacted any sites other than a small number of inshore locations. No substantial pollution events (for example, oil spills) were noted near survey locations in the period. Regardless, care is required in inferring causality for observed coral-cover change in this study, and no assumption should be made that all coral loss was attributable to bleaching.
Reporting summary
Further information on experimental design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
Raw reef fish and invertebrate abundance data and photoquadrats of coral cover are available online through the Reef Life Survey website: https://reeflifesurvey.com.
References
Graham, N. A. J. et al. Lag effects in the impacts of mass coral bleaching on coral reef fish, fisheries, and ecosystems. Conserv. Biol. 21, 1291–1300 (2007).
Graham, N. A. J., Jennings, S., MacNeil, M. A., Mouillot, D. & Wilson, S. K. Predicting climate-driven regime shifts versus rebound potential in coral reefs. Nature 518, 94–97 (2015).
Hughes, T. P. et al. Global warming and recurrent mass bleaching of corals. Nature 543, 373–377 (2017).
Poloczanska, E. S. et al. Responses of marine organisms to climate change across oceans. Front. Mar. Sci. 3, 62 (2016).
Pratchett, M. S. et al. Effects of climate-induced coral bleaching on coral-reef fishes—ecological and economic consequences. Oceanogr. Mar. Biol. Annu. Rev. 46, 251–296 (2008).
Graham, N. A. J. et al. Dynamic fragility of oceanic coral reef ecosystems. Proc. Natl Acad. Sci. USA 103, 8425–8429 (2006).
Stuart-Smith, R. D., Edgar, G. J. & Bates, A. E. Thermal limits to the geographic distributions of shallow-water marine species. Nat. Ecol. Evol. 1, 1846–1852 (2017).
Perry, C. T. & Morgan, K. M. Bleaching drives collapse in reef carbonate budgets and reef growth potential on southern Maldives reefs. Sci. Rep. 7, 40581 (2017).
Morgan, K. M., Perry, C. T., Johnson, J. A. & Smithers, S. G. Nearshore turbid-zone corals exhibit high bleaching tolerance on the Great Barrier Reef following the 2016 ocean warming event. Front. Mar. Sci. 4, 224 (2017).
Hughes, T. P. et al. Global warming transforms coral reef assemblages. Nature 556, 492–496 (2018).
Goreau, T., McClanahan, T., Hayes, R. & Strong, A. Conservation of coral reefs after the 1998 global bleaching event. Conserv. Biol. 14, 5–15 (2000).
Edgar, G. J. & Stuart-Smith, R. D. Systematic global assessment of reef fish communities by the Reef Life Survey program. Sci. Data 1, 140007 (2014).
Hoegh-Guldberg, O. & Salvat, B. Periodic mass-bleaching and elevated sea temperatures: bleaching of outer reef slope communities in Moorea, French Polynesia. Mar. Ecol. Prog. Ser. 121, 181–190 (1995).
Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 50, 839–866 (1999).
Garpe, K. C., Yahya, S. A. S. & Lindahl, U. & Öhman, M. C. Long-term effects of the 1998 coral bleaching event on reef fish assemblages. Mar. Ecol. Prog. Ser. 315, 237–247 (2006).
Wilson, S. K., Graham, N. A. J., Pratchett, M. S., Jones, G. P. & Polunin, N. V. C. Multiple disturbances and the global degradation of coral reefs: are reef fishes at risk or resilient? Glob. Change Biol. 12, 2220–2234 (2006).
Edgar, G. J. et al. Abundance and local-scale processes contribute to multi-phyla gradients in global marine diversity. Sci. Adv. 3, e1700419 (2017).
Wolanski, E., Andutta, F., Deleersnijder, E., Li, Y. & Thomas, C. J. The Gulf of Carpentaria heated Torres Strait and the Northern Great Barrier Reef during the 2016 mass coral bleaching event. Estuar. Coast. Shelf Sci. 194, 172–181 (2017).
Bates, A. E., Stuart-Smith, R. D., Barrett, N. S. & Edgar, G. J. Biological interactions both facilitate and resist climate-related functional change in temperate reef communities. Proc. R. Soc. B 284, 20170484 (2017).
Stuart-Smith, R. D., Edgar, G. J., Barrett, N. S., Kininmonth, S. J. & Bates, A. E. Thermal biases and vulnerability to warming in the world’s marine fauna. Nature 528, 88–92 (2015).
Edgar, G. J., Ceccarelli, D. M. & Stuart-Smith, R. D. Assessment of Coral Reef Biodiversity in the Coral Sea. Report for the Department of the Environment (Reef Life Survey Foundation, 2015).
Vergés, A. et al. Long-term empirical evidence of ocean warming leading to tropicalization of fish communities, increased herbivory, and loss of kelp. Proc. Natl Acad. Sci. USA 113, 13791–13796 (2016).
Diaz-Pulido, G. & McCook, L. J. The fate of bleached corals: patterns and dynamics of algal recruitment. Mar. Ecol. Prog. Ser. 232, 115–128 (2002).
Hughes, T. P. et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 359, 80–83 (2018).
McClanahan, T., Maina, J. & Pet-Soede, L. Effects of the 1998 coral morality event on Kenyan coral reefs and fisheries. Ambio 31, 543–550 (2002).
Mumby, P. J. The impact of exploiting grazers (Scaridae) on the dynamics of Caribbean coral reefs. Ecol. Appl. 16, 747–769 (2006).
Hughes, T. P. et al. Coral reefs in the Anthropocene. Nature 546, 82–90 (2017).
Ferrari, R. et al. Quantifying the response of structural complexity and community composition to environmental change in marine communities. Glob. Change Biol. 22, 1965–1975 (2016).
Hoegh-Guldberg, O. & Bruno, J. F. The impact of climate change on the world’s marine ecosystems. Science 328, 1523–1528 (2010).
Chollett, I., Enríquez, S. & Mumby, P. J. Redefining thermal regimes to design reserves for coral reefs in the face of climate change. PLoS ONE 9, e110634 (2014).
Edgar, G. J., Stuart-Smith, R. D., Cooper, A., Jacques, M. & Valentine, J. New opportunities for conservation of handfishes (Family Brachionichthyidae) and other inconspicuous and threatened marine species through citizen science. Biol. Conserv. 208, 174–182 (2017).
Cresswell, A. K. et al. Translating local benthic community structure to national biogenic reef habitat types. Glob. Ecol. Biogeogr. 26, 1112–1125 (2017).
Edgar, G. J. & Stuart-Smith, R. D. Ecological effects of marine protected areas on rocky reef communities—a continental-scale analysis. Mar. Ecol. Prog. Ser. 388, 51–62 (2009).
Edgar, G. J. et al. Global conservation outcomes depend on marine protected areas with five key features. Nature 506, 216–220 (2014).
Duffy, J. E., Lefcheck, J. S., Stuart-Smith, R. D., Navarrete, S. A. & Edgar, G. J. Biodiversity enhances reef fish biomass and resistance to climate change. Proc. Natl Acad. Sci. USA 113, 6230–6235 (2016).
Althaus, F. et al. A standardised vocabulary for identifying benthic biota and substrata from underwater imagery: the CATAMI classification scheme. PLoS ONE 10, e0141039 (2015).
Stuart-Smith, R. D. et al. Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 501, 539–542 (2013).
Garde, L., Spillman, C. M., Heron, S. & Beeden, R. ReefTemp next generation: a new operational system for monitoring reef thermal stress. J. Oper. Oceanogr. 7, 21–33 (2014).
Gelman, A., Hwang, J. & Vehtari, A. Understanding predictive information criteria for Bayesian models. Stat. Comput. 24, 997–1016 (2014).
Watanabe, S. Asymptotic equivalence of Bayes cross validation and widely applicable information criterion in singular learning theory. J. Mach. Learn. Res. 11, 3571–3594 (2010).
Rue, H., Martino, S. & Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. Roy. Stat. Soc. B 71, 319–392 (2009).
R Core Team. R: A Language and Environment for Statistical Computing http://www.R-project.org/ (R Foundation for Statistical Computing, Vienna, 2013).
Pebesma, E. J. Multivariable geostatistics in S: the gstat package. Comput. Geosci. 30, 683–691 (2004).
Anderson, M. J., Gorley, R. N. & Clarke, K. R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods (PRIMER-E, Plymouth, 2008).
Kleiber, C. & Zeileis, A. Visualizing count data regressions using rootograms. Am. Stat. 70, 296–303 (2016).
Dunn, P. K. & Smyth, G. K. Randomized quantile residuals. J. Comput. Graph. Stat. 5, 236–244 (1996).
Cheal, A. J., MacNeil, M. A., Emslie, M. J. & Sweatman, H. The threat to coral reefs from more intense cyclones under climate change. Glob. Change Biol. 23, 1511–1524 (2017).
Acknowledgements
We thank the Reef Life Survey (RLS) divers and boat skippers who assisted with field surveys, including D. and J. Shields, I. Donaldson and S. Griffiths, and A. Cooper, J. Berkhout and E. Clausius at the University of Tasmania for logistics and data management; J. Stuart-Smith, S. Baker, A. Bates and N. Barrett for further support in the development of RLS, fieldwork and concepts explored in the paper. Development of RLS was supported by the former Commonwealth Environment Research Facilities Program, and analyses were supported by the Marine Biodiversity Hub, a collaborative partnership supported through the Australian Government’s National Environmental Science Programme (NESP), and by the Australian Research Council. Funding and support for the GBR and Coral Sea RLS field surveys was provided by The Ian Potter Foundation and Parks Australia. Permits were provided by Parks Australia and the Great Barrier Reef Marine Park Authority. C.J.B. was supported by a Discovery Early Career Researcher Award (DE160101207) from the Australian Research Council.
Reviewer information
Nature thanks J. Bruno, R. Ferrari and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
G.J.E. and R.D.S.-S. collected the data with the assistance of other Reef Life Survey divers; C.J.B. undertook the data analysis and preparation of figures with assistance from R.D.S.-S.; D.M.C. analysed the photoquadrats for benthic cover data; R.D.S.-S. drafted the paper with input from all other authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Results of GLMMs for changes in coral and algal cover during the 2016 bleaching event.
a, Changes in coral cover. b, Changes in algal cover. Change in cover is modelled as a function of the influences of starting cover (of corals and algae, respectively), wave exposure, thermal anomaly (DHD), the interaction between DHD and starting cover, depth category (depths between 4 and 10 m and >10 m modelled in comparison to <4-m depth), and the interaction between depth category and DHD (n = 211 site–depth combinations). All continuous predictors were normalized to mean = 0 and s.d. = 1 for comparative purposes.
Extended Data Fig. 2 Changes in algal and coral cover spanning the 2016 bleaching event.
a, Coral- and algal-cover change were negatively correlated (ρ = −0.56). b, The greatest algal-cover increases occurred at sites with the lowest coral cover after the bleaching event (ρ = −0.28). n = 211 site–depth combinations.
Extended Data Fig. 3 Ecological changes on surveyed reefs most clearly affected by coral bleaching (red) versus un-impacted reefs (blue).
Reefs categorized as bleached were those with >20% pre-bleaching live coral cover, that experienced >40 DHD and that lost >40% of pre-bleaching coral cover (see Methods for rationale). The un-impacted reefs were those that had >20% pre-bleaching live coral cover and experienced >40 DHD, but did not show a reduction in coral cover. The vertical axis is the percentage change of each metric across the reefs in each category (n = 6 bleached, n = 5 unbleached reefs), and horizontal lines on box plots show median, first and third quartiles, with the range indicated by the error bars. Crosses indicate means and circles indicate individual reefs within quartiles. Values for corallivores, browsing herbivores and scraping herbivores describe change in densities of species in these groups. Densities and species richness are means per 500 m2 (fishes) or 100 m2 (invertebrates). Bleached and unbleached reefs each include reefs from both northern and southern regions. Only coral cover differed noticably between these two groups of reefs (mean difference = −72%, with 95% credible intervals of 25–107%), although there was a small decline in corallivore densities post-bleaching (mean difference = 42% with 95% credible intervals from −0.24 to 78.0).
Extended Data Fig. 4 Effect sizes from GLMMs of regional change for each ecological metric.
Median additive effects of each covariate on the linear expectation for each metric (with 95% credible intervals as error bars) (n = 233 site-by-depth-category combinations). Effect sizes are on a log scale for all metrics, except for sea urchin presence, which gives the effect on the log-odds of presence versus absence. The influences of latitude, and its change from before to after the bleaching event (the interaction between latitude and bleaching (Latitude * bleaching)), are modelled in relation to differences between the GBR and Coral Sea reefs (GBR), wave exposure (Exposure), depth of the survey (depths between 4 and 10 m and >10 m modelled in comparison to <4-m depth), the percentage cover of live hard corals in the survey (Coral cover) and before versus after the bleaching event (After bleaching). Effects for which credible intervals do not overlap zero are indicated with black, rather than grey, points and error bars.
Extended Data Fig. 5 Non-metric multidimensional scaling plots for reef fish and mobile invertebrate communities along the GBR and Coral Sea.
Fish biomass data (top) and invertebrate abundance data (bottom) were averaged across surveys within 2° latitudinal bands, with number labels representing the northern latitude (that is, 21 represents the 2° band from 21° to 23° south). Coral Sea reefs are distinguished from those in the GBR by a ‘C’ in the label. Symbols have been colour-coded for data collected before and after the bleaching event (n = 13 latitudinal bands each before and after).
Extended Data Fig. 6 Changes in the trophic structure of reef fishes following the 2016 mass bleaching event on the GBR and Coral Sea.
Bars represent the proportion of total biomass made up by each trophic group, averaged across surveys on each reef, and reefs ordered by latitude. Cleaners and algal farmers were removed owing to their small contributions to biomass.
Extended Data Fig. 7 Local species richness of juvenile fishes (per 500 m2) before and after the 2016 mass bleaching event on the GBR and Coral Sea.
Species richness is shown before (blue) and after (red) the 2016 mass bleaching event on the GBR (left) and Coral Sea (right). ‘North’ reefs were north of 12° S (n = 10 reefs), and ‘south’ reefs were south of 19° S (n = 19 reefs). Juveniles were classified as any individuals 10 cm or less, for species that exceed 12.5 cm in maximum size. A Bayesian linear model indicated juvenile richness differed between the GBR and Coral Sea, but not between north and south or before and after the bleaching event (mean difference = 1.70 with lower and upper 95% credible intervals of −0.6 to 4.0). The distribution of raw data is shown in box plots, with crosses indicating means and circles indicate individual reefs within quartiles.
Extended Data Fig. 8 The distribution of sampling effort through space and time.
The temporal gap between pre- and post-bleaching surveys (n = 768 surveys total) between GBR and Coral Sea, along the latitudinal gradient, and locations experiencing different heating anomalies. For the box plot, the box shows the interquartile range and whiskers are 1.5× interquartile range.
Supplementary information
Rights and permissions
About this article

Cite this article
Stuart-Smith, R.D., Brown, C.J., Ceccarelli, D.M. et al. Ecosystem restructuring along the Great Barrier Reef following mass coral bleaching. Nature 560, 92–96 (2018). https://doi.org/10.1038/s41586-018-0359-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0359-9
- Springer Nature Limited
This article is cited by
-
Global impacts of marine heatwaves on coastal foundation species
Nature Communications (2024)
-
The impacts of climate change on coastal groundwater
Nature Reviews Earth & Environment (2024)
-
Sustainable blue economy: Opportunities and challenges
Journal of Biosciences (2024)
-
Does climate change increase the risk of marine toxins? Insights from changing seawater conditions
Archives of Toxicology (2024)
-
Distribution shifts in Indo-Pacific humpback dolphins and the co-occurrence of marine heatwaves
Reviews in Fish Biology and Fisheries (2024)