

Abstract
The origin of insect wings has long been debated. Central to this debate is whether wings are a novel structure on the body wall resulting from gene co-option, or evolved from an exite (outgrowth; for example, a gill) on the leg of an ancestral crustacean. Here, we report the phenotypes for the knockout of five leg patterning genes in the crustacean Parhyale hawaiensis and compare these with their previously published phenotypes in Drosophila and other insects. This leads to an alignment of insect and crustacean legs that suggests that two leg segments that were present in the common ancestor of insects and crustaceans were incorporated into the insect body wall, moving the proximal exite of the leg dorsally, up onto the back, to later form insect wings. Our results suggest that insect wings are not novel structures, but instead evolved from existing, ancestral structures.






Similar content being viewed by others

Data availability
All of the data that support the findings of this study are available in the main text or the Supplementary Information.
References
Lozano-Fernandez, J. et al. Pancrustacean evolution illuminated by taxon-rich genomic-scale data sets with an expanded remipede sampling. Genome Biol. Evol. 11, 2055–2070 (2019).
Averof, M. & Cohen, S. M. Evolutionary origin of insect wings from ancestral gills. Nature 385, 627–630 (1997).
Kukalová-Peck, J. Origin of the insect wing and wing articulation from the arthropodan leg. Can. J. Zool. 61, 1618–1669 (1983).
Snodgrass, R. E. Principles of Insect Morphology (Smithsonian Institution, 1935).
Crampton, G. C. The phylogenetic origin and the nature of the wings of insects according to the paranotal theory. J. N. Y. Entomol. Soc. 23, 1–39 (1916).
Jockusch, E. L. & Nagy, L. M. Insect evolution: how did insect wings originate? Curr. Biol. 7, R358–R361 (1997).
Niwa, N. et al. Evolutionary origin of the insect wing via integration of two developmental modules. Evol. Dev. 12, 168–176 (2010).
Clark-Hachtel, C. M., Linz, D. M. & Tomoyasu, Y. Insights into insect wing origin provided by functional analysis of vestigial in the red flour beetle, Tribolium castaneum. Proc. Natl Acad. Sci. USA 110, 16951–16956 (2013).
Ruiz-Losada, M., Blom-Dahl, D., Córdoba, S. & Estella, C. Specification and patterning of Drosophila appendages. J. Dev. Biol. 6, 17 (2018).
Abzhanov, A. & Kaufman, T. C. Homologs of Drosophila appendage genes in the patterning of arthropod limbs. Dev. Biol. 227, 673–689 (2000).
Cohen, S. M. & Jürgens, G. Proximal–distal pattern formation in Drosophila: cell autonomous requirement for Distal-less gene activity in limb development. EMBO J. 8, 2045–2055 (1989).
Cohen, B., Simcox, A. A. & Cohen, S. M. Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development 117, 597–608 (1993).
Campbell, G. & Tomlinson, A. The roles of the homeobox genes aristaless and Distal-less in patterning the legs and wings of Drosophila. Development 125, 4483–4493 (1998).
Beermann, A. et al. The Short antennae gene of Tribolium is required for limb development and encodes the orthologue of the Drosophila Distal-less protein. Development 128, 287–297 (2001).
Angelini, D. R. & Kaufman, T. C. Functional analyses in the hemipteran Oncopeltus fasciatus reveal conserved and derived aspects of appendage patterning in insects. Dev. Biol. 271, 306–321 (2004).
Liubicich, D. M. et al. Knockdown of Parhyale Ultrabithorax recapitulates evolutionary changes in crustacean appendage morphology. Proc. Natl Acad. Sci. USA 106, 13892–13896 (2009).
Kao, D. et al. The genome of the crustacean Parhyale hawaiensis, a model for animal development, regeneration, immunity and lignocellulose digestion. eLife 5, e20062 (2016).
Mardon, G., Solomon, N. M. & Rubin, G. M. dachshund encodes a nuclear protein required for normal eye and leg development in Drosophila. Development 120, 3473–3486 (1994).
Tavsanli, B. C. et al. Structure–function analysis of the Drosophila retinal determination protein Dachshund. Dev. Biol. 272, 231–247 (2004).
Schaeper, N. D., Prpic, N.-M. & Wimmer, E. A. A clustered set of three Sp-family genes is ancestral in the Metazoa: evidence from sequence analysis, protein domain structure, developmental expression patterns and chromosomal location. BMC Evol. Biol. 10, 88 (2010).
Beermann, A., Aranda, M. & Schröder, R. The Sp8 zinc-finger transcription factor is involved in allometric growth of the limbs in the beetle Tribolium castaneum. Development 131, 733–742 (2004).
Schaeper, N. D., Prpic, N.-M. & Wimmer, E. A. A conserved function of the zinc finger transcription factor Sp8/9 in allometric appendage growth in the milkweed bug Oncopeltus fasciatus. Dev. Genes Evol. 219, 427–435 (2009).
Estella, C. & Mann, R. S. Non-redundant selector and growth-promoting functions of two sister genes, buttonhead and Sp1, in Drosophila leg development. PLoS Genet. 6, e1001001 (2010).
Mito, T. et al. Divergent and conserved roles of extradenticle in body segmentation and appendage formation, respectively, in the cricket Gryllus bimaculatus. Dev. Biol. 313, 67–79 (2008).
Ronco, M. et al. Antenna and all gnathal appendages are similarly transformed by homothorax knock-down in the cricket Gryllus bimaculatus. Dev. Biol. 313, 80–92 (2008).
Rauskolb, C., Smith, K. M., Peifer, M. & Wieschaus, E. extradenticle determines segmental identities throughout Drosophila development. Development 121, 3663–3673 (1995).
Wu, J. & Cohen, S. M. Proximodistal axis formation in the Drosophila leg: subdivision into proximal and distal domains by Homothorax and Distal-less. Development 126, 109–117 (1999).
Calleja, M. et al. Generation of medial and lateral dorsal body domains by the pannier gene of Drosophila. Development 127, 3971–3980 (2000).
Ikmi, A., Netter, S. & Coen, D. Prepatterning the Drosophila notum: the three genes of the iroquois complex play intrinsically distinct roles. Dev. Biol. 317, 634–648 (2008).
Diez del Corral, R., Aroca, P., G mez-Skarmeta, J. L., Cavodeassi, F. & Modolell, J. The Iroquois homeodomain proteins are required to specify body wall identity in Drosophila. Genes Dev. 13, 1754–1761 (1999).
Coulcher, J. F., Edgecombe, G. D. & Telford, M. J. Molecular developmental evidence for a subcoxal origin of pleurites in insects and identity of the subcoxa in the gnathal appendages. Sci. Rep. 5, 15757 (2015).
Kobayashi, Y., Niikura, K., Oosawa, Y. & Takami, Y. Embryonic development of Carabus insulicola (Insecta, Coleoptera, Carabidae) with special reference to external morphology and tangible evidence for the subcoxal theory. J. Morphol. 274, 1323–1352 (2013).
Kobayashi, Y. Formation of Subcoxae-1 and 2 in insect embryos: the subcoxal theory revisited. Proc. Arthropod Embryol. Soc. Jpn 48, 33–38 (2017).
Ober, K. A. & Jockusch, E. L. The roles of wingless and decapentaplegic in axis and appendage development in the red flour beetle, Tribolium castaneum. Dev. Biol. 294, 391–405 (2006).
Beermann, A., Prühs, R., Lutz, R. & Schröder, R. A context-dependent combination of Wnt receptors controls axis elongation and leg development in a short germ insect. Development 138, 2793–2805 (2011).
Clark-Hachtel, C. M. & Tomoyasu, Y. Two sets of candidate crustacean wing homologues and their implication for the origin of insect wings. Nat. Ecol. Evol. https://doi.org/10.1038/s41559-020-1257-8 (2020).
Shiga, Y. et al. Repeated co-option of a conserved gene regulatory module underpins the evolution of the crustacean carapace, insect wings and other flat outgrowths. Preprint at bioRxiv https://doi.org/10.1101/160010 (2017).
Boxshall, G. A. & Jaume, D. Exopodites, epipodites and gills in crustaceans. Arthropod Syst. Phylogeny 67, 229–254 (2009).
Boxshall, G. A. The evolution of arthropod limbs. Biol. Rev. 79, 253–300 (2004).
Hansen, H. J. Studies on Arthropoda II (Gyldendalske Boghandel, 1925).
Schram, F. R. Crustacea (Oxford Univ. Press, 1986).
Shultz, J. W. Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications. Zool. J. Linn. Soc. 97, 1–56 (1989).
Snodgrass, R. E. Morphology and mechanism of the insect thorax. Smithson. Misc. Collect. 80, 1–108 (1927).
Ewing, H. E. The Legs and Leg-Bearing Segments of Some Primitive Arthropod Groups: With Notes on Leg-Segmentation in the Arachnida (Smithsonian Institution, 1928).
Imms, A. D. Recent Advances in Entomology (P. Blakiston’s Son & Co. Inc., 1937).
Roonwal, M. L. Studies on the embryology of the African migratory locust, Locusta migratoria migratorioides Reiche and Frm. (Orthoptera, Acrididae). II—Organogeny. Phil. Trans. R. Soc. Lond. B Biol. Sci. 227, 175–244 (1937).
Carpentier, F. & Barlet, J. The first leg segmÿents in the Crustacea Malacostraca and the insects. Smithson. Misc. Collect. 137, 99–115 (1959).
Matsuda, R. Morphology and evolution of the insect thorax. Mem. Entomol. Soc. Can. 102, 5–431 (1970).
Deuve, T. The epipleural field in hexapods. Ann. Soc. Entomol. Fr. 37, 195–231 (2001).
Bäcker, H., Fanenbruck, M. & Wägele, J. W. A forgotten homology supporting the monophyly of Tracheata: the subcoxa of insects and myriapods re-visited. Zool. Anz. 247, 185–207 (2008).
Mashimo, Y. & Machida, R. Embryological evidence substantiates the subcoxal theory on the origin of pleuron in insects. Sci. Rep. 7, 12597 (2017).
Heymons, R. Beiträge zur Morphologie und Entwicklungsgeschichte der Rhynchoten (Druck von Ehrhardt Karras, 1899).
Hansen, H. J. Zur Morphologie der Gliedmassen und Mundtheile bei Crustaceen und Insecten. Zool. Anz. 16, 193–198, 201–212 (1893).
Weber, H. Morphologie, Histologie und Entwicklungsgeschichte Der Articulaten (G. Fischer, 1952).
Sharov, A. G. Basic Arthropodan Stock: With Special Reference to Insects (Pergamon Press, 1966).
Uchifune, T. & Machida, R. Embryonic development of Galloisiana yuasai Asahina, with special reference to external morphology (insecta: Grylloblattodea). J. Morphol. 266, 182–207 (2005).
Kobayashi, Y. The paracoxal suture in insect embryos: its state and importance for understanding the basalmost podomeres. Proc. Arthropod. Embryol. Soc. Jpn 52, 43–44 (2018).
Niwa, N. et al. Correlation of diversity of leg morphology in Gryllus bimaculatus (cricket) with divergence in dpp expression pattern during leg development. Development 127, 4373–4381 (2000).
Requena, D. et al. Origins and specification of the Drosophila wing. Curr. Biol. 27, 3826–3836.e5 (2017).
Wang, D. et al. spalt is functionally conserved in Locusta and Drosophila to promote wing growth. Sci. Rep. 7, 44393 (2017).
Medved, V. et al. Origin and diversification of wings: insights from a neopteran insect. Proc. Natl Acad. Sci. USA 112, 15946–15951 (2015).
Ohde, T., Yaginuma, T. & Niimi, T. Insect morphological diversification through the modification of wing serial homologs. Science 340, 495–498 (2013).
Hu, Y., Linz, D. M. & Moczek, A. P. Beetle horns evolved from wing serial homologs. Science 366, 1004–1007 (2019).
Olesen, J., Haug, J. T., Maas, A. & Waloszek, D. External morphology of Lightiella monniotae (Crustacea, Cephalocarida) in the light of Cambrian ‘Orsten’ crustaceans. Arthropod Struct. Dev. 40, 449–478 (2011).
Alexander, D. E. A century and a half of research on the evolution of insect flight. Arthropod Struct. Dev. 47, 322–327 (2018).
Gruner, H.-E. Über das coxalglied der pereiopoden der isopoden. Zool. Anz. 152, 312–317 (1954).
Dreyer, H. & Wägele, J. W. The Scutocoxifera tax. nov. and the information content of nuclear SSU rDNA sequences for reconstruction of isopod phylogeny (Peracarida: Isopoda). J. Crustac. Biol. 22, 217–234 (2002).
Wolff, C. The embryonic development of the malacostracan crustacean Porcellio scaber (Isopoda, Oniscidea). Dev. Genes Evol. 219, 545–564 (2009).
Prpic, N.-M. & Telford, M. J. Expression of homothorax and extradenticle mRNA in the legs of the crustacean Parhyale hawaiensis: evidence for a reversal of gene expression regulation in the pancrustacean lineage. Dev. Genes Evol. 218, 333–339 (2008).
Kerner, P., Ikmi, A., Coen, D. & Vervoort, M. Evolutionary history of the iroquois/Irx genes in metazoans. BMC Evol. Biol. 9, 74 (2009).
Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A. & Patel, N. H. Fixation and dissection of Parhyale hawaiensis embryos. Cold Spring Harb. Protoc. 2009, pdb.prot5127 (2009).
Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A. & Patel, N. H. In situ hybridization of labeled RNA probes to fixed Parhyale hawaiensis embryos. Cold Spring Harb. Protoc. 2009, pdb.prot5130 (2009).
Choi, H. M. T. et al. Third-generation in situ hybridization chain reaction: multiplexed, quantitative, sensitive, versatile, robust. Development 145, dev165753 (2018).
Sander, J. D., Zaback, P., Joung, J. K., Voytas, D. F. & Dobbs, D. Zinc Finger Targeter (ZiFiT): an engineered zinc finger/target site design tool. Nucleic Acids Res. 35, W599–W605 (2007).
Sander, J. D. et al. ZiFiT (Zinc Finger Targeter): an updated zinc finger engineering tool. Nucleic Acids Res. 38, W462–W468 (2010).
Martin, A. et al. CRISPR/Cas9 mutagenesis reveals versatile roles of Hox genes in crustacean limb specification and evolution. Curr. Biol. 26, 14–26 (2016).
Gloor, G. B., Nassif, N. A., Johnson-Schlitz, D. M., Preston, C. R. & Engels, W. R. Targeted gene replacement in Drosophila via P element-induced gap repair. Science 253, 1110–1117 (1991).
Linz, D. M. & Tomoyasu, Y. Dual evolutionary origin of insect wings supported by an investigation of the abdominal wing serial homologs in Tribolium. Proc. Natl Acad. Sci. USA 115, E658–E667 (2018).
Acknowledgements
We thank C. Clark-Hachtel and Y. Tomoyasu for sharing results before publication. We thank E. Jarvis for the Parhyale phalloidin muscle stains and A. Pomerantz for the Oncopeltus image and whole adult Parhyale image. We thank C. Wolff, T. Deuve, Y. Kobayashi, F. Schram and J. W. Shultz for helpful discussion. We thank J. Shen for confirming that spalt RNAi reduces both the wing and lobe in Locusta. We thank G. Mardon for the image of the Drosophila dac knockout adult leg. This work is supported by the National Science Foundation (NSF; IOS-1257379 to N.H.P.) and NSF Graduate Research Fellowship (to H.S.B.).
Author information
Authors and Affiliations
Contributions
H.S.B. and N.H.P. conceived the experiments. H.S.B. performed the experiments, conceived the model and wrote the manuscript. N.H.P. edited and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Expression of leg gap genes in whole embryos and dissected third thoracic legs (T3).
a–d, Dll-e. e–h, Sp6-9. i–l, dac2. m–p, exd. q–t, hth. Embryonic expression data for Dll-e (Browne 2005; Serano 2015), Sp6-9 (Schaeper 2010), and exd and hth (Prpic 2008) have been previously characterized, but not at the level of individual leg segments. (d) Dll-e is expressed in leg segments 1–5; in the interior of the tergal plate (Tp), coxal plate (Cp), and gill (G). (h) Sp6-9 is expressed in leg segments 1–6. (l) dac2 is expressed in leg segments 3–5. Expression in segment 3 may be stronger at other time points. (p) exd is expressed in the body wall through leg segment 3. Exd is expressed only in the base of the gill (distal gill not visible here). (t) hth is expressed in the body wall through leg segment 5. Hth is expressed only in the base of the gill. Note that both insects and Parhyale share a peculiar disparity between hth expression and function, wherein hth knockout deletion extends one more leg segment further than the hth expression domain. Whole embryo scale bars = 100 µm. Dissected leg scale bar = 25 µm. a, n = 5; b, n = 6; c, n = 7; e, n = 6; f, n = 8; g, n = 8; i, n = 4; j, n = 4; k, n = 5; m, n = 3; n, n = 6; o, n = 4; q, n = 3; r, n = 3; s, n = 8.
Extended Data Fig. 2 T7 endonuclease assay to confirm CRISPR-Cas9 mutagenesis.
For each gene, one or two wild type (WT) hatchlings were assayed, and one, two, or three KO hatchlings were assayed. T7 endonuclease was either added (+) or not added (–) to the heteroduplex mixture. In brief, a ~1 kb region flanking the CRISPR-Cas9 target site by at least 300 bp to either side was amplified by PCR from either WT or KO hatchlings. The purified PCR products were denatured, then slowly cooled to allow WT DNA and mutant DNA with indels to anneal, resulting in a ‘bubble’ of unpaired DNA (heteroduplex) at the target site. T7 endonuclease was added to the (+) samples, incubated, and run on a 1.5% agarose gel. KO animals are mosaic, so if the target site was cut, the indels will cause heteroduplexes when annealed with either a WT strand, or a different indel. When a single deletion is present, each half of the cut heteroduplex adds up to approximately 1 kb (see Sp6-9 KO 1 and 2). Some deletions are large enough to be seen without the T7 endonuclease assay (see Dll-e KO), and some hatchlings had multiple deletions which produced multiple bands when cut with T7 (see exd KO 1, hth KO 2, dac2 KO). In hatchlings that have a phenotype but only the WT band (Exd KO 2 and Hth KO 1), the deletion may be so large that one or both of the primer sites may have been deleted.
Extended Data Fig. 3 CRISPR-Cas9 knockout phenotypes in thoracic legs 4 and 5 (T4, T5).
Note that T4 and T5 are morphologically and molecularly indistinguishable, and are treated as interchangeable here. a, Wild type T4 leg. b, Sp6-9 KO T4/T5 leg. c, dac KO T4/T5 leg. The fused and nearly deleted remnant of leg segments 3–5 are in gray. Gill is unaffected, but became oriented upward during specimen mounting. d, Dll KO T4/T5 leg. Scale bar = 50 µm.
Extended Data Fig. 4 Exd and Hth phenotypes.
Body segment fusions are due to the interaction of exd and hth with engrailed during segmentation (Kobayashi 2003). a, Body segment fusions/deletions in Exd knockout whole hatchling. Confocal of unilaterally affected hatchling, dorsal view, anterior at bottom, posterior at left. Left side of animal (L) appears WT. The foreshortening of only the right (R) half twists the body laterally into a nearly spiral shape. The tissue where the eye (E) would have been located is deleted, leaving a recess. Left first antenna (An1), left and right telson (tL, tR). White brackets compare the length of the body segments in right fused and left unfused segments. b, WT T4 leg. c, WT T3 leg, same as Fig. 2a. d, Exd KO T4 leg. Loss of exd deletes/fuses leg segments 1–4 and proximal 5, while the distal half of leg segment 5, and all of leg segments 6 and 7 are WT, because the joint between leg segments 5 and 6 is normal, but there is no joint on the proximal side of leg segment 5, indicated by the proximally faded cyan shading. exd KO causes transformation of remaining T3 leg segments towards a T4/5 identity: compare blue leg segment 6 of WT T3 (Fig. 2) and T4 legs to that of exd KO T3 (Fig. 2) and T4 legs. e,f, Exd and hth KOs produce similar body segment fusions/deletions and proximal thoracic leg segment fusions/deletions. Colours in leg segments are as in Fig. 2.
Extended Data Fig. 5 Drosophila dachshund KO leg.
dac4/dac4 homozygote from dac mutant lines created by Professor Iain Dawson (Mardon 1994). The trochanter through proximal tarsus (leg segments 3–5, and proximal tarsus) are affected, forming a single, fused tissue. Figure adapted with permission from Graeme Mardon.
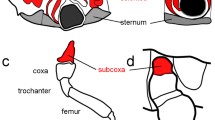
Extended Data Fig. 6 Parhyale precoxa forms a true, muscled joint and extends musculature to another leg segment.
Confocal images. a, Phalloidin stain of muscle in right half of Parhyale hatchling. Contrast simple, anterior-posterior body muscles to orthogonal, complexly arranged leg muscles. No muscles cross the coxa-basis joint, as noted by Boxshall 1997. Note that all three plates (tergal, coxal, and basal) form contiguous cuticle with their leg segment, that is there is no distinguishing suture between leg segment and exite. b, Optical section showing superficial muscles of right half. Cuticle in grey, muscle in pink. c, Confocal of dissected left half, medial view. Coxal plate and basis partially cut. The precoxa forms a joint with two articulations with the coxa: an anterior, bifurcated, load-bearing hinge articulation (arrowhead), and a posterior gliding articulation (arrow). Orthogonal muscles visible as striations on T4 precoxa. d, Close-up of left T4, medial-anterior view, showing bifurcated hinge articulation. In a-d, the precoxa forms two articulations with the coxa: an anterior, bifurcated, load-bearing hinge articulation (arrowhead), and a posterior gliding articulation (arrow). Coxa is red (coxal plate not shaded, to focus on joints), basis is orange, precoxa is magenta pink. The proximal-most region of the legs brace against each other at (<) and (*). The length of the protrusion is twice that of the coxa: compare the beginning (< and *) and end of the protrusion where it forms a joint (arrow and arrowhead) with the coxa (most visible in b). Muscles in green insert on the precoxa-coxa joint, indicating that this is a true joint, and not merely a point of flexure in the exoskeleton (annulation; Boxshall 2004, 2013; Shultz 1989). The shorter, anterior muscle originates in the protruding precoxa to insert on the rim of the next leg segment, the coxa. This muscle is therefore an intrinsic muscle, a hallmark of a true leg segment (Boxshall 2004, 2013; Shultz 1989). Panels a,b adapted with permission from Erin Alberstat.
Extended Data Fig. 7 Exites can be split into anterior and posterior lobes in crustaceans and insects.
a–d, Ectopic wing formation on T1 following Scr RNAi in Tribolium. Mildly affected individuals have wing tissue emerging from unconnected anterior and posterior regions of the body wall. In more severe phenotypes, these anterior and posterior tissues are fused into a more completely transformed wing. e–j, Split exites in crustaceans and insects. Anterior lobe (red outline or arrow), posterior lobe (blue outline or arrow). e, Split coxal plates in Parhyale. f, A malacostracan crustacean, Anaspides, with split anterior and posterior exites. g, Decapod with split anterior and posterior exites (arthrobranchs). h, Oncopeltus with split anterior and posterior supracoxal lobes. i,j, Gin traps require the wing genes vg and ap, consistent with an exite identity. Gin traps have split anterior and posterior jaws. Images adapted with permission from ref. 8, PNAS (a–d); from Erin Alberstat (e); from ref. 39, John Wiley and Sons (f); from ref. 38, Arthropod Systematics & Phylogeny (g); from Aaron Pomerantz (h); and from ref. 78, PNAS (i,j).
Extended Data Fig. 8 Proximal gene expression in later Parhyale and Tribolium embryos.
a, pannier is expressed in the dorsal-most tissue in late Parhyale embryos during dorsal closure. b, Tribolium embryo, vestigial marks the future wing region adjacent to the spiracle. The three domains of araucan expression remain distinct even at later stages.
Supplementary information
Supplementary Information
Supplementary Figs. 1–4, Tables 1–12 and Discussion.
Supplementary Video 1
Supracoxal lobe of a cricket nymph.
Rights and permissions
About this article

Cite this article
Bruce, H.S., Patel, N.H. Knockout of crustacean leg patterning genes suggests that insect wings and body walls evolved from ancient leg segments. Nat Ecol Evol 4, 1703–1712 (2020). https://doi.org/10.1038/s41559-020-01349-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41559-020-01349-0
- Springer Nature Limited
This article is cited by
-
New indications for the life habits of long-legged aphidlion-like larvae in about 100-million-year-old amber
PalZ (2024)
-
Evolution of tissue-specific expression of ancestral genes across vertebrates and insects
Nature Ecology & Evolution (2024)
-
A hemimetabolous wing development suggests the wing origin from lateral tergum of a wingless ancestor
Nature Communications (2022)
-
A single WNT enhancer drives specification and regeneration of the Drosophila wing
Nature Communications (2022)
-
The crustacean Parhyale
Nature Methods (2022)