

Abstract
Recent population recovery of many pinniped species (seals, sea lions, walrus) is a conservation success. However, pinniped population recovery combined with increasing global fisheries operations is leading to increased conflicts between pinnipeds and fisheries. This human-wildlife conflict threatens pinniped conservation outcomes and may impose damaging impacts on fisheries, but the economic consequences and extent of these impacts are poorly understood. Here, we provide a global assessment of pinniped and fisheries operational interactions. We show that a third of reported fishing days have interactions with pinnipeds and 13.8% of catch is lost. Our results also reveal high heterogeneity between studies. Small-scale fisheries are three times more likely to interact with pinnipeds and lose four times as much catch as large-scale fisheries. Finally, we develop a spatial index that can predict where conflict is most likely to occur. Our findings reveal a substantial global issue requiring appropriate management as pinniped populations continue to recover.
Similar content being viewed by others

Introduction
Conflict between marine predators and fishers is a long-standing global issue that has widespread impacts on both conservation outcomes and livelihoods1. Fisheries are a dominant global industry, with a spatial extent over four times greater than terrestrial agriculture and are valued at USD 406 billion2,3. However, fishing activity inevitably attracts marine predators, particularly cetaceans and pinnipeds, which target many of the same prey species as fishers4. Pinnipeds, which include seals, sea lions and walruses, are increasingly reported as interacting with fisheries5,6. Historically, interactions with pinnipeds were frequent, which led to systematic hunting or opportunistic or managed culls. The combined impacts of hunting and culls resulted in average pinniped population declines of over 70% relative to historic baselines7. In the 20th century, the exploitation of pinnipeds coupled with widespread overfishing resulted in many species being threatened with extinction or going extinct8,9,10. To date, several pinniped populations remain threatened with extinction like the Hawaiian Monk Seal (Neomonachus schauinslandi)11, while the status of other species, like the Caspian seal (Pusa caspica), remain unknown due to a lack of data12. Following successful conservation management initiatives, such as the landmark US Marine Mammal Protection Act of 1972, many pinniped species have recovered or are recovering12,13. Magera et al.14 estimated that 44–58% of pinniped species have seen significant increasing population trends since the end of the 20th century. Note that for some species, e.g., the harbour seal (Phoca vitulina), there may be considerable heterogeneity in population trends across regions15. Alongside pinniped population recoveries is a resurgence of reports of conflict with fisheries13,14. Current reports of pinniped-fishery conflict most commonly focus on pinniped depredation of fishery catch, damage to fishing gear (i.e., operational interactions, described below)4,16,17, or pinniped bycatch (the incidental catch of non-target species)18,19.
Pinniped population recovery presents managers with novel, and increasingly prevalent, conservation challenges. As conservation outcomes improve, economic outcomes can deteriorate, prompting concerns about social impacts on fishing communities20. Against this backdrop, managers are increasingly expected to demonstrate ecosystem based management21,22 that balances the triple bottom line—simultaneously achieving ecological, economic, and social objectives23,24. Ecosystem based management requires coexistence of pinnipeds and fisheries, meaning continued protection for marine mammal populations while safeguarding fishing activities and livelihoods1,25. Reports of pinniped-fishery conflict describe two main interactions. The first are biological interactions, which generally describe indirect impacts, e.g., pinnipeds or fisheries consuming or catching a fish stock, which affects how much of that stock is available for subsequent consumption or catch. Research into biological interactions is typically motivated by concerns for recovering pinniped populations, i.e., assessing whether sufficient prey is available for pinnipeds to meet their energetic requirements, or regarding pinniped impacts on recovering fish stocks and broader food-web dynamics26,27. Second, operational interactions, which describe direct interactions between pinnipeds and fisheries during fishing operations. Research into operational interactions includes investigation of pinniped-bycatch risk or fisher retaliation—describing the negative impacts of fisheries on pinnipeds and how these impacts will affect vulnerable or recovering pinniped populations28,29,30. The other aspect of operational interactions is pinniped damage to fishing gear and depredation (consumption) of catch from nets—the frequency and severity of these two events provides a tractable metric to describe the economic impact of pinniped-fisheries conflict31,32. This second aspect of operational interactions is what we focus on in the rest of this text, as estimates are quantifiable and comparable, and thus well suited to synthesise this global issue.
Qualitative reports of operational interactions between pinnipeds and fisheries are widespread. However, a systematic quantitative global assessment of operational interactions remains a major barrier to better understanding pinniped-fishery conflict4,16,17. Tixier et al. 16 found that 214 fisheries from 44 countries, and all but two FAO fishing regions, reported depredation from marine predators between 1979 and 2019, of which 30.8% corresponded to pinnipeds. Marine predators were responsible for an 11% reduction in catch—but how much of this decrease was attributable to pinnipeds is unknown. From a mixture of dockside surveys and onboard observations conducted during 1997–1999, Weise and Harvey33 estimated that the annual monetary loss caused by damage to gear and catch by the California sea lion (Zalophus californianus) in Monterey Bay to commercial salmon fisheries ranged between US$22 K and US$60 K (gear) and US$224 K and US$504 K (catch). In Peru and Chile, two studies documenting pinniped-fisheries conflict report that 65%34 and 87%17 of interviewed fishers have interactions with pinnipeds. For artisanal fishers in the Foça Monk Seal Pilot Conservation Area, Turkey, damage to fishing gear by the critically endangered Mediterranean monk seal (Monachus monachus) was reported 90 times (of 142 observations) between 1992 and 200435. Increasing reports of the economic cost of pinnipeds to fisheries highlight the need for more quantitative assessments of operational interactions to inform our understanding of the wider pinniped-fishery conflict.
To fully quantify the global impacts of pinniped-fishery conflict, we need to ascertain whether reports of pinniped-fishery conflict are representative of where conflict is occurring. This is because a lack of publication in a given area can demonstrate an absence of either conflict or reporting. For example, publication/reporting biases against statistically insignificant results—an issue in both ecology and resource economics publications36,37—could skew research towards areas with higher conflict, thus inflating estimates of the economic impacts of pinniped-fisheries interactions. Moreover, a lack of interaction data from areas identified as potential hot-spots may indicate that other management methods such as legal/illegal culls have already been implemented by fisheries38. Conversely, limitations in data collection and fishery management—perhaps due to resource constraints—may result in missing interaction data, deflating estimates. Therefore, exploring spatial patterns in the reporting of operational interactions, and predicting where pinniped conflict may occur, is a vital step in quantifying the global extent and severity of this issue.
Here, we provide a quantitative global assessment of the impacts of operational interactions between pinnipeds and fisheries. First, we conducted a systematic literature review to synthesise comparable quantitative assessments of operational interactions in articles from the Scopus and Web of Science databases. We used two negative operational interactions as dependent variables: 1) the proportion of fishing days in which interactions between fisheries and pinnipeds were observed, and 2) the proportion of catch lost to pinnipeds. Interactions included any temporally explicit observation of pinniped activity in proximity to, or interference with, fisheries. We included only temporally explicit operational interactions, thus allowing fishing effort to be normalised by fishing days so that observations were comparable, and those primarily assessed by independent observers. Note that we excluded estimates from recreational fishing and only included estimates from aquaculture that could be temporally standardised. Next, we used mixed-effects meta-analyses to estimate pooled impacts of interactions and catch loss on fisheries. We also explored how the study-specific effects of fishery scale (e.g., whether large-scale industrial fisheries or small-scale/artisanal fisheries, herein large- or small-scale) and fishing gear type (net, line, trawl, or other) influenced operational interactions using meta-regression analyses. Finally, by linking recently published global fishing data from Global Fishing Watch3, extant pinniped occurrence data12, and assessing proximity of marine areas to the nearest coast39, we predicted the spatial extent of pinniped-fishery interactions and investigated the spatial biases and global context of reported operational interaction data and their association with global fishing effort3. Our quantitative focus on operational interactions provides a global assessment of the heterogeneity of this conflict. Evidence from our assessment can help inform conservation and economic policies at appropriate scales, particularly those centred on compensation or conflict resolution schemes, whilst achieving conservation goals and protecting human livelihoods.
Results
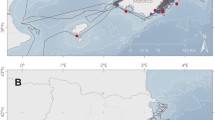
There are a lack of articles presenting quantifiable estimates of the impact of pinniped-fishery operational interactions. Of the 1179 articles we screened from Scopus and Web of Science, 375 articles focused on coexistence and interactions between pinnipeds and fisheries, highlighting that operational interactions are a global issue (Figure S1). Of these 375 articles, several investigated bycatch (n = 50), biological interactions (n = 48), or social impacts (n = 26). Of the articles focussed on operational interactions (n = 58), 22 had insufficient data for extraction (Figure S1). Thus, we retained 36 articles that included quantitative and temporally explicit measures of operational interactions between pinnipeds and fisheries. There were 34 observations of interactions from 24 articles, and 33 observations of the proportion of catch lost to pinnipeds from 22 articles (Table S1; Fig. 1; Supplementary Data 1). Of the pinniped populations assessed in retained articles, 9 were described as having an increasing population, 4 a decreasing trend, and 1 with a stable population (Table S1). There are insufficient articles that standardise interactions by some unit of effort, but a greater number that standardise observations temporally, and therefore we used the number of fishing days represented in reported estimates to account for differences in sampling effort and variance between studies. While there is variation in fishing time based on gear type, vessel size, and other fishery characteristics, we captured information on fishery size and gear type for meta-regressions. Retained articles report observations of interactions with total observation periods of 25,867 and 6,929 days for interaction and catch loss data respectively (Fig. 1). An additional 13 articles present non-standardised measures of interactions and damage (Fig. 1; Supplementary Data 2). These observations were excluded from the meta-analyses due to the omission of temporal information, which was required to standardise observations, or the use of qualitative versus quantitative estimates (e.g., qualitative categories such as ‘frequent’ or ‘infrequent’ used to describe frequency of interactions). Non-standardised data were typically gathered through questionnaires or interviews. This non-standardised data indicates large proportions of fishers report regular interactions (mean n=11 = 61.8 %) and catch damage due to pinnipeds (mean n=8 = 46.6 %) (Figure S2). The retained quantitative articles, while relatively sparse, are distributed across the globe, but there are general biases to South America and Northern Europe (Fig. 1).

The size of the points indicates the number of fishing days in each article. The colour of the points denotes the type of operational interaction data available for each article: Light blue points = interaction data, dark blue = catch loss data, purple points = both, and grey triangles indicate non-standardised data that were excluded from meta-analyses. Illustrative species examples are, from left to right, the Californian sea lion (Zalophus californianus), South American fur seal (Arctocephalus australis), harbour seal (Phoca vitulina - north), South American sea lion (Otaria flavescens - south), grey seal (Halichoerus grypus - north), Brown fur seal (Arctocephalus pusillus pusillus - south), Mediterranean monk seal (Monachus monachus), Caspian seal (Pusa caspica), Antarctic fur seal (Arctocephalus gazella), and the Australian fur seal (Arctocephalus pusillus doriferus).
There are extensive operational impacts of pinnipeds on fisheries (Fig. 2). On average, 33.4% [95% confidence limits: 22.9−45.8] of fishing days are affected by interactions with pinnipeds (Fig. 2a). Furthermore, 13.8% [8.08-22.5] of catch is lost to pinnipeds (Fig. 2b). However, there is a high degree of heterogeneity in meta-analysis outcomes (Fig. 2). For both negative interactions and damage, Cochran’s \(Q\) tests of similarity in binomial responses indicate highly significant differences in responses across studies (\(Q\) = 2141, p < 0.001 and \(Q\) = 997, p < 0.001, respectively), as well as substantial heterogeneity indices of \({I}^{2}\), \({\tau }^{2}\), and \(H\) (Fig. 2). The results of these tests provide clear evidence that global operational impacts vary substantially across fisheries and indicate that other contextual fishery characteristics may be crucial in driving differences in operational interactions.

Meta-analysis results of pooled estimates (middle panels) with respect to observations from each article (top panels), and heterogeneity measures (bottom panels), for articles quantifying the proportion of fishing days with interactions (a; n = 24) and the proportion of catch lost in those days (b; n = 22). In the top panels, each point is an observation from a retained article with estimated 95% confidence limits (Clopper-Pearson interval; Balduzzi et al. 70), with the size of the point giving the number of fishing days, and multiple points indicating repeated observations. The middle panel gives the pooled estimates for both the random effects model and fixed effects only model, but we present random effects results, which accounts for repeated observations across studies, and gives more conservative estimates of uncertainty. Thus, the 95% confidence limits (shaded area) and pooled estimates (vertical line) are presented for the random effects model. The tables in the bottom panels are the heterogeneity measures, with 95% confidence limits.
Fishery characteristics significantly affect the frequency and impact of operational interactions with pinnipeds (Fig. 3). From a total of 15 fishery and study characteristics that we initially identified as potential drivers of operational interactions (Table S2), we retained three characteristics that were reported in sufficient detail across studies: fishery scale, gear type and reported population trend of pinnipeds. Fishery scale (small vs. large) is typically reported in retained studies, although definitions can vary (Table S340). We find small-scale fisheries using nets are significantly more likely to have interactions with pinnipeds. Note that because fishery scale and location were highly correlated—large-scale fisheries are much more likely to operate at greater distances from shore (Figure S3)—we excluded location from meta-regressions. For both proportion of fishing days with interactions and proportion of catch lost, fishery scale has a significant impact on operational interactions (\(\beta\) = 1.37 ± 0.44 SEM, \(z\) = 3.08, p < 0.01 and \(\beta\) = 1.66 ± 0.66 SEM, \(z\) = 2.53, p < 0.05, for interactions and damage, respectively; Fig. 3a, c; Tables S4 & S5). The proportion of fishing days with pinniped interactions for large-scale fisheries is 16.9% [9.10–29.2], in comparison to 44.5% [25.1–65.7] for small-scale fisheries (i.e., small-scale fisheries were over twice as likely to interact with pinnipeds). The proportion of catch damage is four times greater in small-scale fisheries, at 19.8% [6.39–47.2] of catch compared to 4.48% [1.50−12.5] in large-scale fisheries (Fig. 3a, c).

The relationship between fishery scale (a, c) and the gear type used (b, d), and the proportion of fishing days with interactions (a, b) and the proportion of catch lost (c, d). Coloured points are observations from retained articles, with shaded areas indicating the approximate symmetrical data density. Black points and lines are the predicted mean operational interactions with 95% confidence limits. * and ** indicates classification is significantly different to reference group (a, c, ‘Large’, b, d, ‘Line’) at the 95% and 99% levels respectively.
Fishing gear, namely net, line, trawl or other operations, predict operational interactions. Fisheries using nets have a greater proportion of interactions and a greater proportion of catch lost (Fig. 3b, d; Tables S4 & S5). Net fisheries are predicted to interact with pinnipeds on 41.2% [18.0-69.3] of fishing days and lose 17.0% [5.04–44.0] of their catch, in contrast to 15.1% [6.58;−30.8] of fishing days and 3.44% [1.11–10.2] of catch lost in line fisheries (Fig. 3b, d). Fisheries using other gear types and trawls also generally report a greater number of operational interactions (Fig. 3b, d), but reduced sample sizes in these groups prevents reliable inference. The reported local population trend of pinnipeds was available in a smaller subset of studies and is not significantly associated with either the proportion of days with interactions or the proportion of catch lost (Tables S4 & S5).
We provide a global prediction of spatial patterns in pinniped-fishery interactions and explore potential spatial biases in retained articles. Using this spatial tool, we aimed to predict where pinniped-fishery interactions are likely to occur, regardless of reports in the literature, using independent, globally available data. To achieve this aim, we constructed a global composite spatial index of the potential for pinniped-fishery interactions at a resolution of 0.5o. This index incorporates fishing effort (log10-transformed)3, occurrence data for pinniped populations listed on the IUCN Red List of Threatened Species12, and the distance of each 0.5o pixel centroid from shore39, with areas closer to shore likely to have higher pinniped density due to haul out and breeding locations (Fig. 4 & S4). This global index highlights nine regions that are expected to have the highest occurrence of pinniped-fishery interactions: the Bering Sea, the West-Coast of North America, the temperate and cape region of South America, the North-East of North America, Iceland, temperate and arctic regions of Northern Europe, the Cape of Southern Africa, southern Oceania, and the Seas of Japan and East China (Fig. 4). We found that this spatial index of conflict potential is largely robust to changes in the spatial layers describing fishing effort and proximity to shore (Fig. S6).

Pixels represent the composite index of the potential for pinniped-fishery interactions at a spatial resolution of 0.5°, for which high values (purple) indicate high fishing effort, presence of pinnipeds and proximity to coast. Black circles and triangles describe study locations: circles indicate quantitative articles (n = 36) retained in meta-analyses; open triangles indicate non-standardised articles (qualitative, n = 13) that were excluded from our meta-analyses to ensure comparability.
Discussion
Humans have exploited marine resources for at least 40,000 years, and conflict with marine predators such as pinnipeds is just as old41,42. Current conflict between recovering pinniped populations and fisheries often revolves around operational interactions. We find that in areas where operational interactions are quantified and reported, fishers interact with pinnipeds on 33.4% of fishing days and report damages of 13.8% to total catch. The resulting situation is a multifaceted human-wildlife conflict likely to increase globally (Fig. 4). Conflict between pinnipeds and fisheries is often exacerbated by overfishing16, which can lead to increased interactions as fishing effort increases and pinnipeds and fishers coincide more as fish abundance decreases. Bycatch risk to pinnipeds during operational interactions18,19 also leads to worse outcomes for both fisheries and pinniped populations. Bycatch is costly for fisheries through damage to gear and delays to operations, but is deadly for pinniped individuals with potentially severe consequences for recovery or persistence of vulnerable pinniped populations43. Population modelling can be used to assess the sensitivity of pinniped populations to bycatch risks or other fisheries-induced threats44. Assessments focused on modelling the future economic impacts of pinniped-operational interactions must also recognise that pinniped populations are vulnerable to disease risk and climate change impacts when recommending management responses45. Solutions to manage pinniped-fisheries interactions necessitate fishers and other marine industries operating alongside large populations of pinnipeds. Fisheries managers and conservation scientists and practitioners must therefore find new non-lethal ways of managing conflict, which also avoid retaliatory actions by fishers17. Policies to achieve this outcome have to-date been stymied by a lack of quantitative data and analysis4,16. We fill this gap by quantifying the extent and severity of operational interactions between pinnipeds and fisheries. Our results highlight pinniped-fishery conflict as a global economic issue that requires continued and extended management as pinniped populations continue to recover.
While the impacts of operational interactions are ubiquitous, conflict with pinnipeds does not affect all fishers equally, nor do all pinniped-fishery interactions involve conflict. We find that small-scale fishers are almost three times as likely to have interactions with pinnipeds on a given fishing day, and lose four times as much catch, compared to large-scale fisheries. These increases in operational interactions are also observed for fishing operations using nets. Given that small-scale fishers are more likely to operate closer to shore, and that operations involving nets typically provide more opportunity for interactions due to a larger amount of gear being left in the water for longer (i.e., compared to lines)—there is more opportunity for interactions with pinnipeds. On average, our results show that interactions with pinnipeds result in catch losses, however, it is worth noting that several of the studies incorporated into the meta-analysis found negligible or no catch loss resulting from pinniped interactions (e.g., refs. 46,47). Nevertheless, our results highlight the importance of engaging with small-scale fisheries when managing pinniped conflict. The Food and Agriculture Organization of the United Nations declared 2022 the International Year of Artisanal Fisheries and Aquaculture, describing the small-scale fishing industry as small in scale, big in value for the achievement of sustainable development goals48. Ensuring that the livelihoods of this important natural resource sector are protected, perhaps through compensation or other management strategies, will go a long way towards addressing the impacts of pinniped-fishery conflict. Our results further add to an increasingly dominant narrative in human-wildlife conflict research, that the poorest sectors of society experience a disproportionate burden of wildlife-conflict49,50,51,52. A recent global analysis by ref. 52 found that economic vulnerability to conflict with large terrestrial carnivores is between 2 and 8 times greater for communities in developing economies compared to developed economies. Protecting local communities who have the most at stake from conservation management actions is vital to ensure sustainable development and effective environmental protection.
In addition to a quantitative synthesis of the frequency and severity of operational interactions, we take the first steps to predict pinniped-fishery conflict worldwide. To do this, we combine spatial data layers describing the current distribution of pinniped populations12, spatially-explicit fishing effort3, and the proximity of marine areas to the nearest coast39. We find that operational interactions are more likely in areas where pinniped occurrence and high fishing effort overlap. Of the areas we identify as potential hotspots of operational conflicts, the current study has quantitative observations from six of the nine regions—although there is a bias in study localities, particularly towards South America and Northern Europe, which comprise 23 of the 36 retained studies. We lack reports of interactions from the Bering Sea, Iceland, and the Seas of Japan or East China. Although we have some data from southern Oceania, the lack of reports from New Zealand is notable. Nevertheless, the average potential for pinniped-fishery interactions is substantially higher in locations where studies were carried out (Figure S5), suggesting that the measures of global fishing effort, pinniped occurrence, and proximity to coast are useful proxies for predicting areas with operational interactions. Lack of reported quantifiable interactions may occur for a variety of reasons. These reasons include: that fisheries and pinnipeds are targeting different resources in an area so do not interact; insufficient prey abundance to support large pinniped populations due to overexploitation from fisheries53; that alternative conflict management strategies such as culls and hunting have already been implemented (e.g., ref. 54); or a focus on bycatch or other interactions55. Missing quantifiable interactions may also reflect a lack of resources in different regions to conduct research into the topic or that this research is conducted by government agencies and reported in the grey literature. Increased research effort into pinniped-fisheries interactions will help further illuminate the global extent of pinniped-fishery conflict, highlight global reporting biases, and ultimately improve the development of targeted management strategies.
Our spatial predictive tool is targeted at a global scale, but future research could better predict conflict at the regional or local level by including finer resolution spatial data, including local pinniped population density or small-scale fishery activity. Further extensions to this tool include predicting where conflict is likely to occur in the future by modelling how climate change will affect future distributions of pinniped population abundance, using, for example, species distribution or energetics models56, and corresponding shifts in the spatial signature of fishing activity as the distribution of fish stocks change57. Further avenues of research include adding a temporal signature to model predictions, as many pinniped populations will shift their activities throughout the year for breeding and moulting. Similarly, many fisheries change their activities throughout the year as they target different fish stocks. It may be the case that pinnipeds and fisheries do not temporally overlap in some identified hot spots and thus there is no conflict.
Our analysis highlights several information gaps that future research could target to improve quantification of pinniped-fishery conflict. First, despite extensive reporting16, only a small proportion of identified articles report quantifiable, temporally standardised estimates of interactions or damage. Coordinated efforts that increase the independent recording58 of temporally standardised interactions, and ideally extending reporting to be standardised by units of effort (e.g., ref. 50) and thus the relative impacts of conflict, are needed. Second, very few studies extend work on operational interactions to quantify the economic costs of pinniped conflict—either from catch loss or gear damage59, most likely due to difficulty in estimating losses that chiefly occur under the surface. Early efforts from Weise and Harvey33 estimated monetary losses to the commercial salmon fishery by the California sea lion (Zalophus californianus) in Monterey Bay during 1997–1999. They found damage to gear ranged between US$22 K and US$60 K and catch losses ranged from US$224 K to US$504 K. Losses to anglers in the Moray Firth in Scotland from seal damage to nets and catch losses have also been estimated at £16,500 per annum60. Fjälling et al. 61,62 have also collected extensive records of seal damage to salmon-trap nets in the Baltic sea. These studies provide a useful blueprint for future economic assessments. Third, for our spatial predictions we use global fishing data that is biased towards large-scale fisheries with centralised GPS tracking. Given our finding that interactions are disproportionally experienced by small-scale fisheries, future research efforts should prioritise collecting spatial data from this sector. Data from the small-scale fisheries sector is often difficult to source—due to commercial sensitivities regarding profitable fishing grounds. Emphasizing the importance of an automatic identification system (AIS), which transmits a ship’s position, to ensure safety for vessels at sea may incentivise the use of this technology and subsequent data sharing63.
Fisheries conflict with marine predators such as pinnipeds is a global issue, and one that threatens the ideals of ecosystem-based management. To achieve ecological, economic, and social outcomes, managers need information on the economic outcomes of conflict. The interventions needed will certainly vary by location, and our study contributes by illuminating the heterogeneity of the impacts operational interactions can have. We provide this critical missing element through a quantification of the impact on fisheries of operational interactions with pinnipeds. We find that interactions with pinnipeds occur on 33% of fishing days and 14% of catch is lost or damaged by pinnipeds. Equitable management must address the heterogeneous distribution of these costs, which are often disproportionately borne by small-scale fisheries. Using this information, managers can begin to work towards holistic solutions for society, such as fishing gear modifications, deterrence methods, or compensation schemes. However, more work is needed to better understand and map the heterogeneity we unveil, so that interventions can be designed at the appropriate scale with the right incentives. Coexistence with marine predators is the end goal, but knowledge of individual fishery-contexts will be key to achieving working solutions.
Methods
Our approach was to identify relevant articles from the natural and social sciences using SCOPUS and Web of Science. We conducted an initial sorting of relevant articles based on title, abstract, and keywords screening, published in all available years equating to 1st January 1960 through 17th August 2022 for SCOPUS, and 1st January 1945 through 17th August 2022 for Web of Science. The percent overlap of relevant articles between the two search platforms was calculated and the relevant articles were consolidated. We then developed a rubric to guide data extraction and completed a scoping study to determine whether sufficient data existed to calculate our effect sizes of interest. In what follows, we provide an overview of our systematic literature review, the definition of the effect sizes of interest, the final rubric for data extraction, then our analytical methods.
Literature review
We constructed a two-part search string. The first part focussed on fisheries human-wildlife conflict or variants thereof, including human-wildlife conflict, human wildlife interaction, HWC, operational interaction, conflict management, depredation, overexploitation, animal welfare, conservation conflict, marine mammal-fishery interaction, marine mammal predation, artisanal fishery, coastal fishery, commercial fishery, purse-seine fishery, gillnet fishery, interaction with fisheries, marine mammals and fisheries, seal-induced catch*, seal-induced damage*, poaching, culling, acoustic harassment devices, anti-predator nets. The second part used several terms covering groupings of pinnipeds to refine the breadth of search results. Specifically, we searched for pinnipeds, seal*, fur seal, true seal, sea lion*, walrus, marine mammal*, marine predators as key search terms. Based on our search string, we had 966 hits in SCOPUS and 188 unique hits in Web of Science. We broadened this search through following up hits from reference lists of relevant articles. This search method gave us 28 additional articles, of which 25 were unique. An initial assessment based on title and abstract considered that articles were relevant if pinnipeds and fisheries were both mentioned. This left us with 375 articles that we reviewed in depth.
Operational interactions of interest
We focused on two operational interactions in the current study, in which observations of pinnipeds interacting with/damaging fishery operations were made. These two operational interactions were used as response variables in meta-analyses. We ignored biological interactions for quantitative meta-analyses because biological interactions are both less comparable and harder to observe64. Furthermore, we did not include observations of fisher retaliation against pinnipeds, which is rarely quantified, or bycatch effects, which have been explored in detail28,29,30. Therefore, our two effect sizes of interest centred around the likelihood that fishers will have interactions with pinnipeds, and the impact of these interactions on catch. Both effect sizes of interest compared observed numbers of interactions or catch loss relative to the sampling time (described below). These effects are therefore most appropriately captured by a binomial distribution: (1) number of interactions relative to the total number of interactions possible in the sampling time, and (2) catch lost as a proportion of total catch in the sampling time.
To obtain comparable observations for meta-analyses, we retained only temporally-explicit reports of operational interactions. The most widely reported proxy of effort across studies was fishing time, number of trips, or number of sets. We therefore normalised effort by fishing days to describe the total sampling time, i.e., the amount of effort that occurred in one day. For example, if there were 2 fishing events per day (e.g., trips or sets), then the effort normalisation value would be 0.5. For both effect sizes, the sample size used in meta-analyses estimates was the number of fishing days. The rationale for this measure as opposed to using fishing trips or sets was that trips or sets vary substantially between fisheries and gear type, and more generally the reporting of fishing effort varied between studies. While fishing days may also be biased where fishing activities are not constant throughout each fishing event, they provide a more comparable quantitative measure of the temporal component of fishing activities. Ideally, observations would be standardised using explicit metrics of effort such as catch per unit effort (CPUE), but this was not available for the majority of studies (but see ref. 50). For two studies31,65, we used (spatially) neighbouring studies46,66 with similar fishery scale and practises to normalise fishing trips into fishing days. Furthermore, we primarily used studies in which independent observations were made on board fishing vessels, which reduced potential reporting biases. We assumed that observation effort was consistent across fishing days for each study.
The first effect metric we calculated was the proportion of fishing days that had interactions with a pinniped species. An interaction in this context was broadly defined as the presence or activity of pinnipeds near vessels when fishing operations were taking place. These interactions ranged from observations of pinnipeds swimming near fishing operations67 to direct observations of pinnipeds damaging catch64. Thus, the effect size of interest was the proportion of reported fishing days in which interactions were observed. The second effect metric we calculated was the proportion of catch damaged by pinnipeds for a specific fishery and associated number of fishing days. This effect size provided a more explicit measure of pinniped-fishery impacts than interaction frequency, as observers provided quantitative estimates of the proportion of catch lost to pinnipeds either through direct observations of damage, reporting the amount of catch damaged (e.g., with bite marks65), or through counterfactuals comparing sites with and without pinniped activity.
Covariates
To explore potential drivers of differences in operational interactions between studies, we extracted data for a set of biological, fishery, and study characteristics (see Table S2 and Supplementary Data 1). We identified 15 characteristics as potential drivers of operational interactions. Biological characteristics included the focal pinniped species, the reported local population trend of the focal pinniped species, the target fish species, and a binary indicator of whether a study targeted multiple fish species. The local population trend of the focal pinniped species was obtained from information in the article itself and reported as decreasing, stable, or increasing. We did not use external sources such as IUCN regional assessments for this variable, because we aimed to capture the specific spatio-temporal context of pinniped population change relevant to the article. The categorical variable for fishery scale (industrial [large] or small-scale/artisanal [small]) was chosen to capture broad differences between fishery dimensions such as fishing gear, and fishery characteristics, for which fleet and boat sizes were determined with information from each article’s methodology (Table S3). Despite broad and non-uniformly reported definitions of fishery scale, we captured a wide array of characteristics that effectively distinguished the retained articles (Table S3). Specifically, fishery characteristics recorded included the location of fishing operations (near-shore, off-shore, or both), the type of fishing gear used (net, line, trawl, or other), and binary indicators of reporting of pinniped prevention activities (e.g. use of audio exclusionary devices68), economic compensation for losses due to pinnipeds, and retaliation of fishers towards pinnipeds. Fisheries were classified as near-shore if the authors mentioned ‘coastal’ or ‘near-shore’ in their description of the fishing region, off-shore if they mentioned ‘deep-sea’ or ‘off-shore’, or ‘both’. Other study characteristics included the study location (latitude and longitude) and country, the source of the study (affiliation of the first author: academia, government, non-governmental organisation, fisheries, and unknown), and the method of data collection (survey, on-board observations, cameras, and logbooks). The study location was retrieved from coordinate bounding areas (rectangular) reported in each article. Where maps and areas were not reported, a bounding area was estimated from reported coordinate locations, which were expanded to a bounding box by adding/subtracting 5 degree-seconds to each coordinate. The source of the study was collected from the affiliation of the first author, which indicated the employer of the person carrying out the majority of the work.
Meta-analysis
We pooled observations of the proportion of fishing days with interactions and the proportion of catch lost across studies using binomial mixed-effects meta-analyses implemented in the metafor and meta R packages69,70. We used R version 4.1.3 for all analyses71. Binomial meta-analyses were constructed using a successes and trials framework, for which trials were used to give the weighted sampling variance for each article. Thus, successes were calculated from the proportion of interactions and catch lost, given the total number of fishing days. Trials were the total number of fishing days. Models were fit using logit transformed observed proportions69. We used both fixed-effects and random effects models to account for repeated observations from the same article (n = 4 and n = 2 studies for interaction and damage proportions, respectively), but reported more conservative uncertainty estimates from random-effects models70. In meta-analyses, we calculated back-transformed confidence limits at the 95% percent level. Thus, using pooled estimates of the proportion of fishing days with interactions and the proportion of catch lost with 95% confidence limits, we were able to assess whether pinniped-fishery interactions were significantly different from 0. To estimate the heterogeneity of operational interaction estimates, and thus how variable reported interactions were, we performed a set of heterogeneity assessments. These included the Cochrans \(Q\) test for heterogeneity, and the \({I}^{2}\), \({\tau }^{2}\) and \(H\) indicators of heterogeneity. Generally, these methods give an estimate of the average distance of observed proportions to the pooled model estimates.
Following meta-analyses models, we performed meta-regressions to explore how biological, fishery, and study characteristics were associated with operational interactions. From the full set of 15 potential drivers of operational interactions, we retained fishery size, gear type, and reported pinniped population trend for meta-regressions (Table S2). Other characteristics were either not reported in sufficient detail, did not vary sufficiently across studies, or were too variable across studies relative to sample sizes (Table S1 & S2). Meta-regressions were fit to the initial meta-analysis models using the metareg function of the meta package. Here, all potential drivers of operational interactions were categorical, such that meta-regressions tested whether there were significant differences between reference levels within each category relative to other levels (Tables S4 & S5). To prevent overfitting due to low sample sizes from retained studies, we included only univariate meta-regression models where a single predictor of interest was included.
Spatial analyses
To explore spatial biases in quantitative reporting of operational interactions and to predict where pinniped-fishery conflict may occur, we developed a composite spatial index of the potential for pinniped conflict. The goal of this predictive tool was to highlight where we expect pinniped-fishery interactions to occur when they are not systematically reported (as in retained articles), using independent, globally available data. We constructed the spatial index from three core components that are likely to be important for pinniped-fishery interactions, i) global fishing pressure, ii) the occurrence of pinniped populations as listed by the IUCN Red List of Threatened Species, and iii) the proximity of each spatial unit of analysis to a shoreline. The spatial layers used were not based on characteristics extracted from retained articles—instead we aimed to use independent, globally available data to identify data gaps and areas with the potential for operational interactions. The overall rationale for this index was to capture areas where fishing activities overlap with pinniped occurrence, whilst accounting for the increased presence of pinnipeds closer to shore. This global index was constructed at a spatial resolution of 0.5° (approximately 55 km grid-squares at the equator) using the rasterize package72. The index was weighted equally with mean global fishing effort from 2012 to 20203, the spatial distribution of pinniped species from the IUCN Red List of Threatened Species12, and the proximity of 0.5° cells to the closest shore39 (Figure S4). For global fishing effort, the data from Global Fishing Watch, sourced from Kroodsma et al. 3 is biased towards large-scale fishing operations and omits many small-scale fishing activities. However, global spatial data on small-scale fishing effort is not currently available. Therefore, we used our global fishing effort as a proxy for both large-scale and small-scale fishing activity, and explored the robustness of the index by assessing outcomes when fishing effort and proximity to shore data were excluded (Figure S6). We calculated fishing effort in each grid square as the sum of fishing hours for all vessels operating in each grid square between 2012–2020, which was scaled to an index between 0 and 1, first standardising fishing hours (with one hour added to avoid observations of 0) on a log10 scale and dividing by the maximum sum of fishing hours across grid squares. Pinniped occurrence data by species group was converted to an index of between 0-1, where occurrence was scored as 1 and non-occurrence as 0.01, to indicate a 1% probability of observing a pinniped species in a given grid square. To explore how well the spatial index predicted observed occurrences of interactions from the literature, we compared the distribution of potential for pinniped conflict values for raster cells in which studies occurred, to the global distribution of index values (Figure S5). Furthermore, we tested how robust the spatial index was to each individual layer. We re-calculated the scaled index but removed either fishing effort data or the proximity to shore data and found that the resulting indices were highly correlated (Figure S6).
Data availability
All data supporting meta-analyses results are from published literature and presented as Supplementary Tables or as Supplementary Data. Additional data used in this study was obtained from publicly available datasets, which are referenced in the text.
Code availability
All analysis, code, and data from the current study are publicly available in a reproducible format, archived on Zenodo (https://doi.org/10.5281/zenodo.12579910), which was created from the following GitHub repository: https://github.com/jjackson-eco/pinniped_meta_analysis.
References
Nyhus, P. J. Human-Wildlife Conflict and Coexistence. Annu. Rev. Environ. Resour. 41, 143–171 (2016).
FAO, World Food and Agriculture – Statistical Yearbook 2021 (FAO, Rome, https://doi.org/10.4060/cb4477en 2021).
Kroodsma, D. A. et al. Tracking the global footprint of fisheries. Science 359, 904–908 (2018).
Guerra, A. S. Wolves of the Sea: Managing human-wildlife conflict in an increasingly tense ocean. Mar. Policy 99, 369–373 (2019).
Chasco, B. E. et al. Competing tradeoffs between increasing marine mammal predation and fisheries harvest of Chinook salmon. Sci. Rep. 7, 15439 (2017).
DeMaster, D. P., Fowler, C. W., Perry, S. L. & Richlen, M. F. Predation And Competition: The Impact Of Fisheries On Marine-Mammal Populations Over The Next One Hundred Years. J. Mammal. 82, 641–651 (2001).
Lotze, H. K. & Worm, B. Historical baselines for large marine animals. Trends Ecol. Evol. 24, 254–262 (2009).
Gerber, L. R. & Hilborn, R. Catastrophic events and recovery from low densities in populations of otariids: implications for risk of extinction. Mammal. Rev. 31, 131–150 (2001).
Jackson, J. B. C. et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 293, 629–637 (2001).
Kovacs, K. M. et al. Global threats to pinnipeds. Mar. Mammal. Sci. 28, 414–436 (2012).
Baker, J. D. et al. Estimating Contact Rates Of Hawaiian Monk Seals (Neomonachus Schauinslandi) Using Social Network Analysis. J. Wildl. Dis. 52, 533–543 (2016).
IUCN, The IUCN Red List of Threatened Species. http://www.iucnredlist.org (2016).
Roman, J. et al. The Marine Mammal Protection Act at 40: Status, recovery, and future of U.S. marine mammals. Ann. N. Y. Acad. Sci. 1286, 29–49 (2013).
Magera, A. M., Mills Flemming, J. E., Kaschner, K., Christensen, L. B. & Lotze, H. K. Recovery trends in marine mammal populations. PloS one 8, e77908 (2013).
Blanchet, M.-A., Vincent, C., Womble, J. N., Steingass, S. M. & Desportes, G. Harbour Seals: Population Structure, Status, and Threats in a Rapidly Changing Environment. Oceans 2, 41–63 (2021).
Tixier, P. et al. When large marine predators feed on fisheries catches: Global patterns of the depredation conflict and directions for coexistence. Fish. Fish. 22, 31–53 (2021).
Davis, K. J. et al. Local disconnects in global discourses—The unintended consequences of marine mammal protection on small-scale fishers. Conservation Letters 14 (2021).
Reeves, R., McClellan, K. & Werner, T. Marine mammal bycatch in gillnet and other entangling net fisheries, 1990 to 2011. Endang. Species Res. 20, 71–97 (2013).
Punt, A. E. et al. Assessing pinniped bycatch mortality with uncertainty in abundance and post-release mortality: A case study from Chile. Fish. Res. 235, 105816 (2021).
Waldo, Å. et al. Local attitudes towards management measures for the co-existence of seals and coastal fishery - A Swedish case study. Mar. Policy 118, 104018 (2020).
Hall, S. J. & Mainprize, B. Towards ecosystem-based fisheries management. Fish. Fish. 5, 1–20 (2004).
J. S. Link, A. R. Marshak, Ecosystem-Based Fisheries Management: Progress, Importance, and Impacts in the United States. (Oxford University Press., 2022).
Dickman, A. J. Complexities of conflict: The importance of considering social factors for effectively resolving human-wildlife conflict. Anim. Conserv. 13, 458–466 (2010).
Roman, J., Dunphy-Daly, M. M., Johnston, D. W. & Read, A. J. Lifting baselines to address the consequences of conservation success. Trends Ecol. Evolution 30, 299–302 (2015).
R. Woodroofe, S. Thirgood, A. Rabinowitz, People and Wildlife: Conflict or Coexistence? (Cambridge University Press, Cambridge, 2005).
Horimoto, T., Mitani, Y. & Sakurai, Y. Spatial association between northern fur seal (Callorhinus ursinus) and potential prey distribution during the wintering period in the northern Sea of Japan. Fish. Oceanogr. 25, 44–53 (2016).
Hindell, M. A., Bradshaw, C. J. A., Sumner, M. D., Michael, K. J. & Burton, H. R. Dispersal of female southern elephant seals and their prey consumption during the austral summer: relevance to management and oceanographic zones. J. Appl. Ecol. 40, 703–715 (2003).
Hamer, D. J. et al. The endangered Australian sea lion extensively overlaps with and regularly becomes by-catch in demersal shark gill-nets in South Australian shelf waters. Biol. Conserv. 157, 386–400 (2013).
Dmitrieva, L. et al. Assessment of Caspian Seal By-Catch in an Illegal Fishery Using an Interview-Based Approach. PLoS ONE 8, e67074 (2013).
Johnston, D. W. et al. Trends in stranding and by-catch rates of gray and harbor seals along the northeastern coast of the United States: Evidence of divergence in the abundance of two sympatric phocid species? PLoS ONE 10, e0131660 (2015).
Sepúlveda, M. et al. Operational interaction between South American sea lions Otaria flavescens and artisanal (small-scale) fishing in Chile: Results from interview surveys and on-board observations. Fish. Res. 83, 332–340 (2007).
Bell, C., Shaughnessy, P., Morrice, M. & Stanley, B. Marine mammals and Japanese long-line fishing vessels in Australian waters: operational interactions and sightings. Pac. Conserv. Biol. 12, 31–39 (2006).
Weise, M. & Harvey, J. Impact of the California sea lion (Zalophus californianus) on salmon fisheries in Monterey Bay, California. Fish. Bull. 103, 685–696 (2005).
Goetz, S., Wolff, M., Stotz, W., Villegas, M. J. & Goetz, S. Interactions between the South American sea lion (Otaria flavescens) and the artisanal fishery off Coquimbo, northern Chile. ICES J. Mar. Sci. 65, 1739–1746 (2008).
Güçlüsoy, H. Damage by monk seals to gear of the artisanal fishery in the Foça Monk Seal Pilot Conservation Area, Turkey. Fish. Res. 90, 70–77 (2008).
Ferraro, P. J. & Shukla, P. Feature—Is a Replicability Crisis on the Horizon for Environmental and Resource Economics? Rev. Environ. Econ. Policy 14, 339–351 (2020).
Parker, T. H. & Yang, Y. Exaggerated effects in ecology. Nat. Ecol. Evol. 7, 1356–1357 (2023).
Iacarella, J. C., Clyde, G., Bergseth, B. J. & Ban, N. C. A synthesis of the prevalence and drivers of non-compliance in marine protected areas. Biol. Conserv. 255, 108992 (2021).
NASA Ocean Biology Processing Group, Distance to the Nearest Coast. https://oceancolor.gsfc.nasa.gov/docs/distfromcoast/ (2009).
Smith, H. & Basurto, X. Defining Small-Scale Fisheries and Examining the Role of Science in Shaping Perceptions of Who and What Counts: A Systematic Review. Front. Mar. Sci. 6, 236 (2019).
O’Connor, S., Ono, R. & Clarkson, C. Pelagic Fishing at 42,000 Years Before the Present and the Maritime Skills of Modern Humans. Science 334, 1117–1121 (2011).
T. C. Rick, J. Erlandson, Eds., Historical ecology and human impacts on coastal ecosystems of the Santa Barbara Channel region, California. In Human Impacts on Ancient Marine Ecosystems: A Global Perspective (University of California Press, Berkeley, 2008).
Goldsworthy, S. D. & Page, B. A risk-assessment approach to evaluating the significance of seal bycatch in two Australian fisheries. Biol. Conserv. 139, 269–285 (2007).
Karamanlidis, A. et al. Assessing accidental entanglement as a threat to the Mediterranean monk seal Monachus monachus. Endang. Species Res. 5, 205–213 (2008).
Silva, W. T. A. F. et al. Risk for overexploiting a seemingly stable seal population: influence of multiple stressors and hunting. Ecosphere 12, e03343 (2021).
Szteren, D. & Páez, E. Predation by southern sea lions (Otaria flavescens) on artisanal fishing catches in Uruguay. Mar. Freshw. Res. 53, 1161 (2002).
Oliveira, L. R. D. et al. Assessing the economic impact caused by South American sea lions based on onboard check versus fishermen’s perception: The two sides of the same coin. Mar. Policy 121, 104193 (2020).
FAO, International year of artisanal fisheries and aquaculture: 2022 Global Action Plan. (United Nations, Geneva, https://www.fao.org/artisanal-fisheries-aquaculture-2022/home/en/ (2022).
Bond, J. & Mkutu, K. Exploring the Hidden Costs of Human–Wildlife Conflict in Northern Kenya. Afr. Stud. Rev. 61, 33–54 (2018).
Distefano, E., Human-Wildlife Conflict worldwide: collection of case studies, analysis of management strategies and good practices. (Food and Agricultural Organization of the United Nations (FAO), Sustainable Agriculture and Rural Development Initiative (SARDI), Rome, Italy, 2005).
Fentaw, T. & Duba, J. Human–Wildlife Conflict among the Pastoral Communities of Southern Rangelands of Ethiopia: The Case of Yabello Protected Area. J. Int. Wildl. Law Policy 20, 198–206 (2017).
Braczkowski, A. R. et al. The unequal burden of human-wildlife conflict. Commun. Biol. 6, 182 (2023).
Teh, L. S. L., Witter, A., Cheung, W. W. L., Sumaila, U. R. & Yin, X. What is at stake? Status and threats to South China Sea marine fisheries. Ambio 46, 57–72 (2017).
IUCN, Zalophus japonicus: Lowry, L. The IUCN Red List of Threatened Species 2017: e.T41667A113089431, International Union for Conservation of Nature; https://doi.org/10.2305/IUCN.UK.2017-1.RLTS.T41667A113089431.en (2014).
Hamilton, S. & Baker, G. B. Review of research and assessments on the efficacy of sea lion exclusion devices in reducing the incidental mortality of New Zealand sea lions Phocarctos hookeri in the Auckland Islands squid trawl fishery. Fish. Res. 161, 200–206 (2015).
Gende, S. M. & Sigler, M. F. Persistence of forage fish ‘hot spots’ and its association with foraging Steller sea lions (Eumetopias jubatus) in southeast Alaska. Deep Sea Res. Part II: Topical Stud. Oceanogr. 53, 432–441 (2006).
Palacios‐Abrantes, J. et al. Timing and magnitude of climate‐driven range shifts in transboundary fish stocks challenge their management. Glob. Change Biol. 28, 2312–2326 (2022).
Suuronen, P. & Gilman, E. Monitoring and managing fisheries discards: New technologies and approaches. Mar. Policy 116, 103554 (2020).
Ríos, N., Drakulic, M., Paradinas, I., Milliou, A. & Cox, R. Occurrence and impact of interactions between small-scale fisheries and predators, with focus on Mediterranean monk seals (Monachus monachus Hermann 1779), around Lipsi Island complex, Aegean Sea, Greece. Fish. Res. 187, 1–10 (2017).
Butler, J. R. A., Middlemas, S. J., Graham, I. M. & Harris, R. N. Perceptions and costs of seal impacts on Atlantic salmon fisheries in the Moray Firth, Scotland: Implications for the adaptive co-management of seal-fishery conflict. Mar. Policy 35, 317–323 (2011).
Fjälling, A. The estimation of hidden seal-inflicted losses in the Baltic Sea set-trap salmon fisheries. ICES J. Mar. Sci. 62, 1630–1635 (2005).
Fjälling, A., Wahlberg, M. & Westerberg, H. Acoustic harassment devices reduce seal interaction in the Baltic salmon-trap, net fishery. ICES J. Mar. Sci. 63, 1751–1758 (2006).
Ou, Z. & Zhu, J. AIS Database Powered by GIS Technology for Maritime Safety and Security. J. Navigation 61, 655–665 (2008).
Sepúlveda, M. et al. Factors affecting the operational interaction between the South American sea lions and the artisan gillnet fishery in Chile. Fish. Res. 201, 147–152 (2018).
Söffker, M. et al. The Impact of Predation by Marine Mammals on Patagonian Toothfish Longline Fisheries. PLoS ONE 10, e0118113 (2015).
Roche, C. & Guinet, C. Marine Mammals And Demersal Longline Fishery Interactions In Crozet And Kerguelen Exclusive Economic Zones: An Assessment Of Depredation Levels. CCAMLR Sci. 14, 67–82 (2007).
Königson, S., Lunneryd, S.-G., Stridh, H. & Sundqvist, F. Grey Seal Predation in Cod Gillnet Fisheries. Cent. Balt. Sea. J. North. Atl. Fish. Sci. 42, 41–47 (2010).
Bordino, P. et al. Reducing Incidental Mortality Of Franciscana Dolphin Pontoporia Blainvillei With Acoustic Warning Devices Attached To Fishing Nets. Mar. Mammal. Sci. 18, 833–842 (2002).
Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 36, 1–48 (2010).
Balduzzi, S., Rücker, G. & Schwarzer, G. How to perform a meta-analysis with R: a practical tutorial. Evid.-Based Ment. Health 22, 153–160 (2019).
R Core Team, R: A language and environment for statistical computing., R Foundation for Statistical Computing; http://www.r-project.org/ (2022).
P. Murrell, rasterize: Rasterize Graphical Output, version 0.1 (2019).
Acknowledgements
We thank all authors of the primary literature used in the current meta-analysis, whose tireless work on this issue is improving our understanding of pinniped-fishery interactions. This publication arises from research funded by the John Fell Oxford University Press Research Fund (0010971, K.J.D.). We also acknowledge support from the Marie Skłodowska-Curie Actions (Grant agreement ID: 101067850, J.J.), and the Alexander von Humboldt Foundation (NZL-1218398-HFST-P, W.N.S.A.). Illustrations by Allysa Adams (@allysaadamsillustration).
Author information
Authors and Affiliations
Contributions
K.J.D. conceived the study, with contributions from W.N.S.A., R.O., and later contributions in study design by J.J. K.J.D., W.N.S.A. and R.O. performed initial literature search and screening. All authors contributed to final literature screening. J.J. analysed the data. J.J. and K.J.D. wrote the manuscript, with critical contributions from W.N.S.A. and R.O. All authors approved the final manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Maritza Sepúlveda and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jackson, J., Arlidge, W.N.S., Oyanedel, R. et al. The global extent and severity of operational interactions between conflicting pinnipeds and fisheries. Nat Commun 15, 7449 (2024). https://doi.org/10.1038/s41467-024-51298-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-51298-6
- Springer Nature Limited