

Abstract
Planktonic calcifying organisms play a key role in regulating ocean carbonate chemistry and atmospheric CO2. Surprisingly, references to the absolute and relative contribution of these organisms to calcium carbonate production are lacking. Here we report quantification of pelagic calcium carbonate production in the North Pacific, providing new insights on the contribution of the three main planktonic calcifying groups. Our results show that coccolithophores dominate the living calcium carbonate (CaCO3) standing stock, with coccolithophore calcite comprising ~90% of total CaCO3 production, and pteropods and foraminifera playing a secondary role. We show that pelagic CaCO3 production is higher than the sinking flux of CaCO3 at 150 and 200 m at ocean stations ALOHA and PAPA, implying that a large portion of pelagic calcium carbonate is remineralised within the photic zone; this extensive shallow dissolution explains the apparent discrepancy between previous estimates of CaCO3 production derived from satellite observations/biogeochemical modeling versus estimates from shallow sediment traps. We suggest future changes in the CaCO3 cycle and its impact on atmospheric CO2 will largely depend on how the poorly-understood processes that determine whether CaCO3 is remineralised in the photic zone or exported to depth respond to anthropogenic warming and acidification.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
The marine calcium carbonate (CaCO3) cycle is a key component of the global carbon cycle, and is intimately related to atmospheric CO2 (ref. 1). The formation of CaCO3 in the ocean is a process largely controlled by the biological calcification of marine organisms2. Planktonic calcifying organisms at the base of the food web (from primary producers to zooplankton) have played a key role since the Mesozoic3, via processes including regulation of surface water alkalinity, ballasting of organic matter and alkalinity export, and establishment of a pelagic carbonate buffer capable of influencing major CO2 change4,5,6.
Since the seminal work of Milliman7,8,9 on the production and accumulation of CaCO3 in the ocean, several studies have aimed to quantify total CaCO3 pelagic production10,11,12 and the contribution of specific calcifying plankton groups. However, the relative contribution of the main calcifying taxa to total CaCO3 pelagic production has not yet been directly quantified.
There is large uncertainty in total pelagic CaCO3 production, with current estimates varying between 0.7–4.7 Pg C yr−1 (0.6−3.9×1014 mol CaCO3 yr−1)7,10,11,12,13. In general, estimates based on satellite observations or modeling of ecosystems/carbonate chemistry in the surface ocean suggest higher CaCO3 production10,12,13 whereas estimates based on export from the production layer typically report lower values7,8,11.
There is also uncertainty on the make-up of pelagic CaCO3 production by different groups. Mainly based on sediment trap export fluxes and sediment data, there is a general understanding that coccolithophores (single-celled haptophytes inhabiting the photic zone, performing photosynthesis and producing calcite) and planktonic foraminifera (single-celled marine eukaryotes producing calcite) each contribute ~50% to the global pelagic CaCO3 production and sedimentation8,14,15,16. However, more recent papers have highlighted the potential of shelled pteropods (specialized free-swimming pelagic sea snails producing aragonite) as an important component of pelagic CaCO3 production13,17,18. Other taxa such as heteropods (holoplanktonic gastropods with aragonite shells, Pterotracheoidea) may also contribute to a lesser degree.
These planktonic calcifying taxa have specific mechanisms of biogenic calcification, and associated differences in vulnerability to ocean acidification19,20 with their shell solubility depending on their specific polymorph mineralogy and Mg content21,22,23. In addition, planktonic calcifiers exhibit a large range of particulate inorganic carbon to particulate organic carbon ratios (PIC/POC)18,24,25, which influences the integrated carbon export rain ratio, an important term for carbon cycling in the oceans and atmospheric CO21,26. Furthermore, and as we discuss later, the association of PIC and POC together within calcifying organisms may play a critical role in driving CaCO3 dissolution above the saturation horizon27. However, despite the importance of pelagic calcification to the marine carbon cycle, key questions remain about pelagic CaCO3 production rates, standing stocks, and export fluxes. Most importantly, it is critical to determine the contributions of different planktonic calcifying groups to pelagic calcification, the proportion of aragonite versus calcite, and the magnitude of CaCO3 production compared to export.
The North Pacific Ocean is a key region for understanding the role of pelagic calcifiers in the global CaCO3 budget, due to its large volume and the wide range of biogeochemical conditions from the subtropical to subpolar gyres. In addition, the waters of the North Pacific are some of the most undersaturated in the global ocean with respect to calcite and aragonite, and thus calcifying organisms in the region are most at risk of future ocean acidification driven by anthropogenic CO2 emissions28,29. Although there are studies of the relative distribution of pelagic calcifiers in the North Pacific30,31, estimates of their relative contribution to CaCO3 standing stock and production rates are severely lacking. The discovery of excess alkalinity above the saturation horizon in the North Pacific has sparked debate about the role of different pelagic calcifiers and their contribution to the alkalinity budget above the thermodynamic saturation horizon23,32,33.
We conducted a research cruise from subtropical to subpolar North Pacific waters in which we assessed the pelagic living CaCO3 standing stock. We targeted the main planktonic calcifiers at five survey stations, from Honolulu, Hawaii, to Seward, Alaska, (Fig. 1, Tables S1 and S2). We deployed plankton nets to sample calcifying zooplankton and rosettes of Niskin bottles to target calcifying phytoplankton. In addition, four intermediate planktonic towing stations were sampled and integrated into the overall data set (Fig. 1, Tables S1, S2). Coccolithophores, foraminifera, pteropods, and heteropods were quantified and the CaCO3 biomass was estimated, providing the first overall picture of the total CaCO3 living standing stock (i.e. inventory), and the relative contribution of the main calcite and aragonite planktonic producers in the productive upper ocean. Using estimates of turnover time for each group we estimate annual production (Methods), and compare this to aragonite and calcite biomineral export out of the surface ocean to 100 and 200 m water depth estimated using floating sediment traps deployed during the time of sampling34 and historical time series in the region.

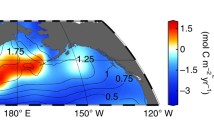
August satellite-derived Particular Inorganic Carbon97 (PIC; mg CaCO3 m-3) climatology (2002-2017) and location of C-DisK-IV stations (black crosses) and long-term sediment trap studies (orange/pink crosses). Large black crosses show the location of Niskin bottle rosette, plankton tow, and floating sediment trap sampling sites at C-DisK-IV stations. Small black crosses show sites with additional plankton tow sampling only at C-DisK-IV stations. Note the logarithmic scale.
Results and discussion
North Pacific CaCO3 standing stocks
The total CaCO3 standing stock is lower in the nutrient-poor and less productive subtropical gyre (~560–900 mg m-2; note here and throughout we refer to mg of CaCO3, unless specified otherwise), and strongly increases into the nutrient-rich and productive subpolar gyre (~1700–4500 mg m−2 total) (Figs. 2 and 3a, Table S3), reflecting the major ecological shift across the North Pacific, from low-CaCO3 production in the oligotrophic subtropics, to high-CaCO3 production in the subpolar region32,35. We find that coccolithophores dominate the CaCO3 producing standing stock at all stations, demonstrating a mean contribution of ~79% (with a range of 62–96% across all sites) to the total CaCO3 standing stocks. Pteropods contribute ~14% (3–29% range), followed by foraminifera (~6% mean, 0.1–22% range), and heteropods (~1%, 0–2%). Calcite from coccolithophores and foraminifera is thus the most abundant mineral, constituting ~86% of the standing stock (71–96%), with aragonite making up ~14% of the standing stock (4–30%).

a chlorophyll fluorescence. Coccosphere, and coccolith CaCO3 from C-DisK-IV stations (b) 1 and 2 and, (c) 3, 4, and 5. Note the different x axis range on panels (b) and (c). d Omega calcite and aragonite at the five stations.

a living CaCO3 standing stock b turnover time of calcifying taxa used to calculate production from standing stock (the range represented by the bar length is applied with a flat probability distribution in our error propagation) c CaCO3 production per day (August 2017) d CaCO3 annual production corrected for seasonal bias using satellite-derived PIC/chlorophyll97,104 and zooplankton seasonality estimates (all data and metadata are publicly available at hahana.soest.hawaii.edu/hot/hot-dogs/interface.html). The total CaCO3 production is shown by the violin plots in panels (c) and (d), where the probability density of the estimate is represented by the thickness of the shaded area and the grey lines show the 68% and 95% confidence interval (CI); note the non-normal distribution with the high-tail on the upper estimate. Error bars for the standing stock (a) and production (c, d) by individual taxon represent the 95% CI (Methods). STG, TZ, and SPG represent subtropical gyre, transition zone, and subpolar gyre, respectively. Purple bands on panels a, c, and d show 68% range of pteropod standing stock and daily/annual production calculated using the MAREDAT database (Methods). The blue stars on panel d show the estimates of total production calculated with in-situ pH and fCO2 measurements at Ocean Station PAPA (ref. 57, light blue), and estimates of production in the subpolar North Pacific calculated using the seasonal cycle of alkalinity and dissolved inorganic carbon (ref. 12, dark blue). STG, TZ, and SPG represent subtropical gyre, transition zone, and subpolar gyre, respectively.
The coccosphere CaCO3 standing stock depth profiles follow the overall chlorophyll fluorescence, albeit with the scaling between fluorescence and coccosphere CaCO3 varying between stations, (Fig. 2, Fig. S1), indicating a substantial contribution by haptophytes/coccolithophores to the total standing stock of photosynthetic algae production36. We observe a shallowing of the chlorophyll maximum depth and coccolithophore CaCO3 standing stock maximum from subtropical to subpolar stations (Fig. 2).
Our estimates of living coccolithophore CaCO3 standing stocks range from 0.13 mg m-3 at 175 m (St. 1) in the subtropical gyre to 110 mg m-3 at 30 m (St. 5) in the subpolar gyre, with depth-integrated estimates from 753 mg m-2 to 3048 mg m-2 at the same stations, respectively (Fig. 2b, c, Tables S2 and S3). Our results support previous work in the North Pacific based on coccolithophore cell concentration, which showed that their biomass is highest at high latitudes, decreasing in temperate and subtropical regions30 (Fig. S8). The subpolar and transitional North Pacific Ocean is also known as a region of sustained seasonal E. huxleyi (the most abundant and cosmopolitan coccolithophore species) blooms37 with an estimated maximum satellite-derived PIC concentration of ~0.8 mmol m−3 (CaCO3 of 80 mg m-3) in August/September (note, satellite PIC is limited to retrievals over the first optical depth of satellite data, ~10 m). There is a remarkable agreement between our estimated values of coccolithophore CaCO3 from the shallowest sampled water depth (~5 m, i.e. the surface CaCO3 concentration) and satellite-derived PIC concentrations (Figs. S2 and S3), which are mostly tuned to capture coccolithophore PIC. This supports the high correlation between satellite-derived PIC and measured PIC surface water concentrations suggested by Balch and others;10,38 however as previously noted38 and discussed later, much of the coccolith PIC production can occur below the depth of the optical retrieval, particularly in the subtropics where the production layer deepens.
We also quantify the CaCO3 contribution of loose coccoliths (calcite plates extruded to the cell surface forming the coccosphere; Fig. 2b; Table S5). Coccoliths are shed into the surrounding waters following death and breakup of the coccosphere, or produced continuously by some species38,39,40. We found that loose coccolith CaCO3 can contribute significantly to the total CaCO3 standing stock in the productive photic layer, with maximum values of 44 and 64 mg m-3 in E. huxleyi blooms at Stations 3 and 5 (Fig. S3). Our results show loose coccolith distribution is tied to the distribution of intact coccospheres and PIC, as observed in previous studies10,38. We note that our coccolithophore living standing stock estimates (used to calculate CaCO3 production rate) only include whole coccosphere cells, and excludes loose coccoliths.
Our pteropod standing stock concentrations in the subpolar gyre range from 109–802 ind. m-3, broadly within the published range of the pteropod standing stocks in the northwestern Pacific (e.g.; 41,42, Fig. 3; Table S3; Fig. S5) and the Gulf of Alaska43,44, although pteropods show a significant seasonal and inter-annual variability in the coastal habitats of the Gulf of Alaska43,44. The recent study of Bednaršek et al.45 found that the abundance of pteropods collected in May 2015 in the subpolar gyre and Gulf of Alaska was (42–423 ind m-2), around two orders of magnitude lower than our abundances observed in August 2017 in the same region (Stations 4–6, Table S3) and previous studies43. This difference in abundance may relate to seasonal and inter-annual variability in this region, and/or the use of larger mesh size in their study (200–335 µm)45, which could have resulted in an underrepresentation of the small size pteropod (juvenile) fraction. Our estimates from the subtropical gyre (22–391 ind. m-3) (Table S3) are similar to previous estimates for this region and are higher than values observed across the Atlantic Ocean46.
Our pteropod CaCO3 concentrations range from 0.2–8.6 mg m-3 (Fig. 3a). We find good agreement between our estimates of pteropod CaCO3 standing and North Pacific sites in the MAREDAT database17 (Methods; Fig. 3), which show a typical concentration of 0.5 (0.2–1.1, 68% CI) mg m-3. Despite our pteropod CaCO3 standing stock results being similar to/higher than previous estimates from the North Pacific/North Atlantic, our values are lower than the proposed 23.17 mg CaCO3 m-3 global-mean value of shelled pteropod CaCO3 standing stock concentration18. However, our analysis shows that this global dataset is heavily skewed (Skewness = 13.3, where a value above one indicates a skewed dataset), with the median pteropod biomass value reported18 being around three orders of magnitude smaller than the reported mean. As such the global-mean value is not a useful descriptor of this data compilation18. Our analysis shows typical pteropod CaCO3 biomass globally is 0.3 (0.08–0.9, 68% CI) mg m-3 in the upper 250 m and 0.2 (0.07–0.8, 68% CI) mg m-3 in the upper 1000 m (Methods; Supplemental Fig. S6), two orders of magnitude lower than the global-mean value reported by ref. 18, and in line with our results from the North Pacific.
Our vertically integrated pteropod CaCO3 standing stocks range between ~64–111 mg m-2 in subtropical gyre and between ~215–1306 mg CaCO3 m-2 in the subpolar gyre (Fig. 3; Table S3; Fig. S5). We find good agreement between our estimates and the vertically integrated pteropod CaCO3 standing stock calculated from North Pacific sites in the MAREDAT database17 (Methods; Fig. 4), which show a typical value of 121 (50–270, 68%) mg m-2, with our estimates thus being slightly higher.

a Pteropod CaCO3 biomass estimated from the MAREDAT database17 and measured in this study; note, maximum values extend above 2 mg m-3. Probability density of pteropod (b) Carbon biomass (c) CaCO3 biomass (d) Integrated CaCO3 standing stock (e) daily CaCO3 production calculated using samples in the upper 250 m of the North Pacific from the comprehensive MAREDAT database17,18 (Methods). Red shading indicates 32–68% confidence interval range. Red values show the 32nd, 50th, 68th percentiles; orange value shows the value with the highest probability (all values given in mg). Note the distributions are highly skewed.
Our heteropod standing stock concentrations range from 5–40 ind. m-3 and 0.01–0.1 mg CaCO3 m-3, and their presence is limited to the subtropics and transition zone. Although previous estimates of heteropod standing stocks are extremely scarce, comparison to previous abundances from a latitudinal Atlantic Ocean transect confirmed that heteropods almost exclusively inhabit warm waters and the recorded maximum of 0.7 ind. m-3 (ref. 46) is lower than our estimates in the North Pacific. Our vertically integrated heteropod CaCO3 standing stocks range from 3-35 mg CaCO3 m-2; heteropods thus contribute between 3–12% of the total aragonite standing stock in the subtropics and transition zone, but are absent from the subpolar region.
Our estimates of integrated foraminiferal CaCO3 standing stock range from 9–37 mg m-2 in the subtropical gyre to 182–404 mg m-2 in the subpolar gyre. Although previous estimates of foraminiferal standing stock in the North Pacific are scarce, our estimates of the integrated vertical standing stock of the number of foraminifera from the subtropical gyre/transition zone sites are similar to, or slightly higher, than previous estimates of the integrated vertical standing stock from the subtropical gyre/transition zone in the western North Pacific31,47 (Figs. S4, 5). Our estimates of the vertically integrated standing stock of the number of foraminifera in the subpolar North Pacific (190,000–250,000 ind. m-2) are generally higher than the estimates of Taylor et al.31, which ranged up to ~80,000 ind. m-2, although such high values are not unprecedented, with previous estimates of the vertically integrated standing stock from the North Atlantic ranging up to ~390,000 ind. m-2 ref. 48.
Pelagic CaCO3 production
We calculate CaCO3 production rate by dividing our measurements of the living CaCO3 standing stock by estimates of the turnover time (i.e. typical life span) of each group (Methods, Fig. 3b). Our approach assumes all of the organisms within the standing stock are living; this is valid for foraminifera, pteropods, and heteropods as individuals sink relatively quickly after death47,49, and the individuals sampled contained cytoplasm/soft tissue (Methods). For coccolithophores this assumption is valid as we only consider intact coccospheres, which disaggregate quickly upon death into the component coccoliths50, and is supported by the fact that the peaks of coccolithophore CaCO3 match the peaks in chlorophyll fluorescence (Fig. 2). We include the caveat that our approach assumes the living standing stock is in approximate steady state.
Given that coccolithophores have a shorter turnover time (1.5–10 days) than the other calcifying groups (Methods, Table 1, Fig. 3b) and dominate the CaCO3 standing stock, they account for ~86% (67–97%) of total CaCO3 annual production across the sites. Pteropods contribute ~10% to total production (2–17%), heteropods ~0.3% (0–1%), and foraminifera contribute ~2% (0.02–9%). As such, 89% of the CaCO3 production is calcite (70–97%), with the remainder being aragonite (Fig. 3c).
Given the large seasonality of PIC production51,52 (Fig. S7), we estimate annual CaCO3 production correcting for seasonal bias (Methods, Table S6). Our seasonally corrected annual CaCO3 production estimates range from 0.2–0.4 mol m-2 yr-1 in the subtropical gyre, a similar range or slightly lower than the estimate of the production rate of 0.7 mol m-2 yr-1 in the subtropical/tropical Atlantic53, although in good agreement with the global mean estimate of 0.4 mol m-2 yr-1 (ref. 10, Fig. 3d). Our estimates from the transition zone and the productive subpolar gyre are higher (0.9–1.0 mol m-2 yr-1) than this global average even at the 95% confidence interval (Fig. 3d); however, they agree well with the estimate of production calculated with in-situ pH and fCO2 measurements at Ocean Station PAPA through (1.2 mol m−2 yr-1; ref. 54), and estimates of production in the subpolar North Pacific calculated using the seasonal cycle of alkalinity and dissolved inorganic carbon (0.9 ± 0.1 mol m-2 yr-1; ref. 12).
To explore the implications of our estimates of CaCO3 production for global production, we use a global climatology of satellite-derived PIC (Fig. S7). While there is a high correlation between satellite PIC and our estimates of surface PIC concentration (Fig. S2a; ref. 38), our results indicate depth integrated CaCO3 production is only twice as high in the nutrient-rich subpolar gyre than the nutrient-poor subtropical gyre, smaller than the range expected from the satellite PIC (CaCO3) estimates, which suggest PIC concentrations ~6–7 times higher in the subpolar region compared to the subtropics. This difference likely reflects the deeper (coccolithophore) CaCO3 production in the subtropics, as well as the thickness of the coccolithophore productive layer (Fig. 2) from the upper ~175 m in the subtropics to the upper ~75 m in subpolar region, which will bias the satellite-derived PIC estimates to lower values38. For Stations 1–4 surface PIC is below 10 mg m- 3 (both at the time of sampling and in the annual mean climatology), yet we observe a depth integrated seasonally corrected production of 0.2–1 mol m-2 yr-1 at these sites (Table S6). Note, this surface PIC value is very similar to the threshold proposed by Balch et al.38 (0.13 mmol m-3/ 13 mg m-3) between surface-dominated and depth-dominated CaCO3 production regimes. Similar low surface PIC regimes (with annual surface PIC of <10 mg m-3) represent 87% of the surface of the ocean (Fig. S8); thus, assuming similar rates of CaCO3 production as the seasonally corrected production rates at Stations 1–4 globally puts a minimum estimate (assuming the remaining 13% of the ocean with higher surface PIC will have higher depth integrated production) for total global CaCO3 production of ~2.2×1014 mol yr-1 (2.6 Pg C yr-1), that is 0.8×1014 mol yr-1 using the production rate at Station 1 and 3.6×1014 mol yr-1 using the production rate at Station 3.
To make a first-order approximation of the impact of deepening CaCO3 production on global CaCO3 production we use a simple linear regression of total CaCO3 production at our sites against satellite PIC (CaCO3) (Fig. S2), with the deepening of production primarily manifesting as a non-zero intercept (note the high variability in the intercept coming from the changing production regimes from Stations 1 to 4 despite the low surface PIC at all Stations). We then apply this relationship to global satellite PIC climatology (Fig. S8). We include the caveat that 1) this assumes the bias caused by the deepening of CaCO3 production in the subtropics scales with surface PIC in a similar way globally38, and 2) the regression in Fig. S2 is driven by one station with high surface PIC (Station 5). While crude, this approach allows us to make a first-order approximation of the impact of deepening CaCO3 production on global CaCO3 production. Applying the relationship between total production and satellite PIC (Table S7) to the global mean surface satellite-derived PIC climatology (Fig. S8), and integrating globally (weighting by area) results in a total CaCO3 production of 3.1×1014 mol yr-1 (3.7 Pg C yr-1) globally. This estimate is similar to, although toward the upper end, of previous estimates of total pelagic calcification based on satellites, upper water column measurements, seasonal alkalinity changes, and ecosystem modeling which range from 0.9–3.9×1014 mol CaCO3 yr-1 (1.1–4.7 Pg C yr-1)10,12,13,28,32,55. However, as previously noted by others10,13 this estimate is considerably higher than estimates of global mean CaCO3 export flux from the upper ocean which ranges from 0.5–0.6×1014 mol yr-1 (0.6–0.7 Pg C yr-1; refs. 7,8).
While our estimate of the total amount of CaCO3 produced agrees well with that of Buitenhuis et al.13, there is a large discrepancy between our results and those of Buitenhuis et al.13 in terms of the dominant CaCO3 polymorph produced. We find CaCO3 production is dominated by calcitic coccolithophores, however, their results suggested pelagic CaCO3 production is mainly driven by aragonite pteropods with coccolithophores and foraminifera playing a minor role.
Given the discrepancy with the results of Buitenhuis et al.13 and the limited temporal interval of our sampling and the potential for large temporal variability of pteropod abundances43, we also calculate pteropod CaCO3 biomass and production in the North Pacific using the comprehensive MAREDAT database18, which has excellent spatial and seasonal sampling distribution in the North Pacific (Methods, Fig. 4). This results in a typical pteropod CaCO3 biomass in the upper 250 m of the North Pacific of 0.5 mg m-3 (0.2–1, 32–68% CI; note the dataset is highly skewed, Fig. S6), a vertically integrated pteropod CaCO3 biomass of 122 mg m-2 (50–269, 32–68% CI), and a pteropod CaCO3 production rate of 12 mg m-2 day-1 (5–27, 32–68% CI). The results calculated using the MAREDAT database are thus in good agreement with the estimates calculated using the samples collected during our own cruise (Fig. 3). We propose the discrepancy with the results of Buitenhuis et al.13 instead comes from three other factors: Firstly, their model parametrization uses a fixed PIC/POC ratio of 0.1 ref. 56 for coccolithophores; this value is substantially lower than the published review by Gafar et al.25 which ranged from 0.19 to 2.30 and much lower than the value 0.52 they used for pteropods, which is itself about two times higher than the estimates of Bednaršek et al.18 (Table 1). Secondly, they assume a similar turnover time for (single-celled) coccolithophores and (complex) pteropods, contrary to the available estimates from the literature57,58,59,60 (Table 1). Finally, within their calculation they assume all CaCO3 dissolving above the calcite saturation horizon is aragonite, an assumption which is likely to exaggerate aragonite production; as we discuss below, previous studies52,61,27 and our results indicate substantial dissolution of coccolithophore calcite above the calcite saturation horizon (which we attribute to respiration-driven dissolution and dissolution within the guts of grazers) such that this assumption is likely to be invalid.
CaCO3 sinking and export fluxes versus production
Our estimate of 3.1×1014 mol CaCO3 yr-1 global pelagic CaCO3 production is ~30 – 300 times larger than required to meet the 0.8 1012 - 1.1×1013mol CaCO3 buried in deep sea sediments each year8,11,62,63,64 and balance the riverine of input of alkalinity to maintain steady-state, reaffirming previous findings that most of the CaCO3 produced in the surface ocean is dissolved and recycled within the ocean interior (e.g., ref. 8). More surprisingly, at several stations our estimates of CaCO3 production are larger than the export fluxes at 100 to 200 m water depth in floating sediment trap deployed during the plankton sampling34, and our production estimates at Stations 1 and 5 are higher than the long running shallow sediment traps at Station ALOHA65 and Ocean Station PAPA64,66,67) (Fig. 5). While the discrepancy observed with the floating traps deployed during the sampling interval (in place for ~72 hrs) may be explained by a decoupling of CaCO3 production and natural mortality/sinking of pteropods, and coccolith aggregation, such that there could be a time lag between production at the surface and export through the water column, this time lag cannot explain the discrepancy observed with the long running sediment traps at Ocean Station PAPA and Station ALOHA. Our results show an annual production of 0.4 (0.2-2.1, 95% CI) and 0.9 (0.5-3.8) mol CaCO3 m-2 yr-1 at Stations 1 and 5, which is ~5 times higher than the annual export of 0.08 and 0.16 mol CaCO3 m-2 yr-1 at 150 m depth at ALOHA and 200 m at PAPA (Fig. 5c, d). We reiterate that previous estimates of annual CaCO3 production at PAPA station based on seasonal cycle of seawater carbonate chemistry support our production value12,54,57.

a total CaCO3 production versus sinking flux in the floating traps deployed at 100 m and 200 m during the plankton sampling at all stations (PIC concentrations not available at 200 m depth for stations 1 and 3)34 b Station 1/ALOHA 65 c Station 5/PAPA;64 turquoise star represents production estimate at PAPA from ref. 54 based on the seasonally cycle of in-situ pH and fCO2 d fraction aragonite in production and sinking flux in the floating traps deployed during the plankton sampling as a function of latitude; red dashed line shows the depth of aragonite saturation horizon (calculated from GLODAPv2105 and orange dotted line shows depth of deepest floating trap. Production in all panels is produced during the time of sampling (August 2017) i.e. it is not corrected for seasonal bias. Error bars for the total production (a, b, c) and fraction aragonite of production (d) represent the 95% CI (Methods) See legend in panel a for square symbols in panels (c, d, e). STG, TZ, and SPG represent subtropical gyre, transition zone, and subpolar gyre, respectively.
This disparity between the amount of CaCO3 produced, and the amount of CaCO3 that is exported out of the photic zone, suggests that a large portion (~80%) of the total CaCO3 produced in the photic zone is never exported, and is instead remineralised in situ; that is, only ~20% of the total CaCO3 produced is exported from the photic zone. Bishop & Wood52 suggested up to 92% of the total CaCO3 produced dissolved within the upper 500 m in the subpolar North Pacific. In situ remineralisation of such a high fraction of the CaCO3 that is produced within the photic zone explains the previous discrepancy between higher estimates of global CaCO3 production based on satellites, upper water column measurements, seasonal alkalinity changes, and ecosystem modeling (which all estimate the total amount of CaCO3 produced)10,13,28 and lower estimates of CaCO3 based on export production (such as sediment traps below the photic zone)7,8.
Shallow remineralisation of pteropods is suggested by the fraction of aragonite present in the shallow traps versus the production ratio, with the fraction of aragonite in the shallow traps decreasing northwards as the aragonite saturation horizon shoals, while the fraction produced increases (Fig. 4e); the highest fraction aragonite produced is at Station 5, where we also observe the lowest fraction in the shallow traps, and the aragonite saturation horizon shoals to above the depth of the shallow trap (<200 m). This dissolution pattern in living pteropod communities in the Gulf of Alaska has been observed recently by Bednaršek et al.45 However, our results indicate that not only aragonitic pteropod and heteropods dissolve13, but also a large amount of coccolithophore calcite. Our findings thus require CaCO3 dissolution above the calcite/aragonite saturation horizon throughout the North Pacific27. Tracers of excess alkalinity support widespread shallow dissolution, far above the calcite and aragonite saturation horizons, throughout the global ocean23,27.
The large amount of CaCO3 dissolution above the calcite/aragonite saturation horizon may be driven by multiple mechanisms, including localized undersaturation within the microenvironment driven by the remineralisation of organic matter, as well as dissolution within the guts of grazers and predators23,33,45,68,69. Both pteropods and coccolithophores contain a large fraction of organic carbon (Table 1), the respiration of which after their death can promote dissolution from the inside out (as well as making them a more attractive food source to grazers). Although the degree to which this could drive total dissolution of whole pteropod shells is still uncertain70, widespread dissolution of coccolithophore CaCO3 within the upper water column has been observed during bloom events (e.g., ref. 71). Furthermore, coccolithophores (which constitute by far the largest fraction of CaCO3 produced) disintegrate into individual coccoliths after death (this can be seen in the coccolith standing stocks in Fig. 2), and must sink by forming aggregates (e.g. marine snow). In situ micro-electrode measurements demonstrate a large pH drop with marine-snow during respiration72, which would drive further micro-environmental undersaturation and CaCO3 dissolution27. In addition, shallow sediment traps indicate coupling of PIC and POC remineralisation from 100 m to 200 m depth in the North Pacific34.
Although the processes by which coccolithophore CaCO3 dissolves in supersaturated waters remain uncertain, some insight may be gained from the residence time of loose coccolith CaCO3 within the production layer; dividing the loose coccolith CaCO3 standing stocks (Fig. 2) by the estimated whole coccosphere CaCO3 production rate at each station gives an approximate coccolith CaCO3 residence time on the order of several days to a couple of weeks. Given the requirement of coccoliths to sink by aggregation and fecal pellets73, the export and dissolution of coccolithophore calcite may occur in discrete events, possibly associated with episodes of high organic carbon production and grazing, rather than sinking as a steady rain, in agreement with previous suggestions based on seasonal export production in sediment trap studies (e.g., refs. 4,50). While our results suggest that a large portion of coccolithophore CaCO3 is remineralised in the photic zone, the very high rate of production means a substantial amount is still exported4,5, and incorporated within sediments14.
Foraminifera comprise ~20% of the total CaCO3 flux in a sediment trap at 3800 m at Station PAPA;74 however, our results show they consist of only ~4% of the total annual CaCO3 standing stock, and ~1% of the total annual CaCO3 production at Station 5. Due to their mass and low organic carbon content24,75 (Table 1), foraminifera sink quickly after death50, have a low self-dissolution potential, and are thus efficient exporters of CaCO3 out of the photic zone. Thunell & Honjo74 found the August flux of foraminiferal calcite in a sediment trap at 3800 m at Station PAPA ranged from 2−6 mg m-2 day-1. Our foraminiferal calcite production estimate at Station 5 is 9 mg m-2 day-1, suggesting foraminiferal calcite is efficiently exported into the deep ocean. Foraminifera thus play an important role in CaCO3 export and sedimentation, despite their low contribution to total production; estimates of CaCO3 in sediments suggest a ~50/50 ratio of coccolithophores/foraminifera13 versus the ~90/2 production ratio we observe.
While much attention has been given to decreasing calcification under ocean acidification19,76 our results indicate a decoupling of CaCO3 production and export (Fig. 5). As such, future changes in the processes driving shallow dissolution, and the ability to export CaCO3 out of the photic zone may play an equally important role in determining the future response of the CaCO3 cycle and its effects on the marine carbon cycle under anthropogenic climate change. Changes in grazing, particle aggregation, the PIC/POC ratio of the aggregates, or the relative abundance of foraminifera to coccolithophores/pteropods, could lead to large changes in the amount of CaCO3 exported from the surface ocean and thus the cycle of alkalinity. The PIC/POC ratio of coccolithophores has been demonstrated to decrease with increasing CO2 (ref. 25); if the dissolution of coccolithophore CaCO3 within the photic zone is in part related to the degree of calcification and/or the remineralisation of organic carbon contained within the soft tissue of the calcifying organisms, this decrease in the PIC/POC of coccolithophores may lead to a negative feedback with CO2, with increased dissolution (and thus reduced export of alkalinity) out of the surface ocean acting to buffer rising atmospheric CO2. Given the potential importance of CaCO3 export in driving changes in alkalinity and atmospheric CO2, and the large uncertainties in our current understanding, future work should focus on understanding the processes by which CaCO3 is either dissolved within the photic zone or exported to depth.
Methods
Sample collection
Samples were collected along a transect from Hawaii to Alaska during August 2017 as part of the CDisK-IV (KM1712) cruise on R/V Kilo Moana (Fig. 1). The five stations along the transect were designed to sample subtropical, transition zone, and subpolar waters. A rosette of Niskin bottles equipped with CTD (conductivity, temperature, depth) and other sensors for coccolithophore and biogeochemical parameters and a vertically integrated plankton tow were collected at each station. Further plankton tows were conducted at four additional intermediate stations (Supplementary material).
A 0.5 m diameter net with 90 µm mesh size was used throughout; based on previous work this mesh size should provide a good estimate of both pteropod18,77 and foraminiferal78 biomass. The sampling strategy was designed to capture an integrated sample of all foraminifera, pteropods, and heteropods from juveniles to adults living throughout the upper water column. The net was towed from the surface down to a specified maximum depth within the water column, and then back to the surface in a continuous manner following an oblique trajectory through the water column. The maximum depth was determined from the fluorescence profile of the preceding CTD cast, and was selected to ensure the net sampling captured well below the base of the chlorophyll maximum and ranged from 150 m in the most northerly subpolar sites to 300 m in the subtropical region (Tables S1, S2). The volume of water represented by each net tow sample was calculated by multiplying the net area by the distance traveled as determined by a flowmeter. For the vertically integrated values, the integration is carried out from the surface to the maximum depth of the tow.
After collection, samples were preserved in a 4% formalin seawater solution, buffered to a pH of ~8.1 with hexamethylenetetramine73. Samples were split with a Folsom splitter or a McLane rotary splitter (splitting error <4%). Large pteropods and heteropods (>1 mm) were picked and quantified before splitting. Half of the split sample was transferred into ethanol solution in the laboratory for the analysis of pteropods and heteropods.
Water samples from rosettes of Niskin bottles equipped with CTD (Sea-Bird SBE 9) were collected at different depths throughout the photic zone and including the chlorophyll maximum depth.
Quantification of calcifying plankton community standing stock and biomass
All foraminifera were wet picked from the sample splits, divided into groups greater and less than 125 µm, counted, and weighed with a high precision microbalance. We assume the mass of organic matter is negligible since dry cytoplasm has no statistically significant effect on the weight of tests > 150 μm24,75. Empty tests made up a negligible component of the standing stock, typically comprising <2% of the total standing stock. Replicate picking and weighing of splits gave a typical reproducibility of ±4% (1σ). Foraminiferal assemblages from these samples were previously reported31.
Pteropods and heteropods were quantified and shell diameter was measured using a Leica Z16 AP0 binocular light microscope at 20−100×. Pteropods and heteropods were identified and grouped respectively in three (Cavoliniidae, Cymbuliidae, Limacinidae) and two (Atlantidae, Carinidae) families.
CaCO3 biomass (B) was estimated as follows: CaCO3 (mg) = PIC (mg) ×8.33 (assuming that all inorganic carbon is in the form of CaCO3), where the constant 8.33 represents the molecular mass ratio of carbon to CaCO3 and PIC is the Particulate Inorganic Carbon79. To estimate PIC we used the PIC/POC ratio of 0.27:0.73 calculated on pteropods by Bednaršek et al.18, where POC is the Particulate Organic Carbon (POC) representing the soft tissue of the organisms. POC was estimated by converting Wet Weight (WW, mg) to Dry Weight (DW, mg) using80 equation DW = WW × 0.28. DW was subsequently transformed to POC using the conversion factor POC = DW × 0.25 following81. WW and/or DW were calculated from the shell length (L, mm) using equations based on the different shell shape: ref. 80,82,83.
For Cymbulidae, we generated the following equation DW = (0.0392 × L)−0.003 from the measurement of shell (mm) and DW of 67 individuals.
For Atlantidae we generated the following equation CaCO3 = 0.769e0,0023L (R2 = 0.885) from the measurement of shell length (mm) and CaCO3 biomass (mg) of 85 individuals. For the measurement of CaCO3 biomass heteropods were heated to 550 °C for 5 h to eliminate organic matter content and the ashes (representing the remains of the shells) weighed using a Toledo microbalance. The ash weight can be considered an indirect estimate of CaCO3 content84.
Between 2.1 and 6.0 liters of seawater were filtered onto Millipore cellulose acetate membranes, with 0.45 µm of pore size and 47 mm of diameter, for coccolithophore analysis. Filters were oven dried at 40 °C and stored in plastic petri dishes. A portion of each filter was mounted on a glass slide and analyzed by a polarized LEICA DM6000 light microscope at 1000× magnification along radial transects whose area was between 0.35 and 3.14 mm2. Cell concentrations per liter of seawater were estimated as follows:
where F is the effective filtration area (mm2), C is the number of coccospheres, V is the filtered seawater volume (L) and A is the investigated filter area (mm2).
Taxa were identified following taxonomic concepts for living coccolithophores by ref. 85,86. To estimate the CaCO3 contribution by coccolithophore assemblages in each sample, we carried out the transformation of coccospheres in number of coccoliths, following the estimates by ref. 87, and then we adopted the coccolith mass estimates by88. The coccolith mass of Noëlaerhabdaceae also took into account the estimates by ref. 89 considering different mass for E. huxleyi calcification degree.
The estimate of coccosphere calcite was further complemented by the individual coccolith (detached from the coccosphere) calcite concentration quantified by microscopy along radial transects of 0.32 mm2, and subsequent mass estimates as described above.
The integrated total living coccolithophore calcite standing stock was obtained considering the first shallow sampling depth to a depth equal to 1% of the fluorescence peak. In St1 it ranges from 6 m to 180 m, Station 2 5–215 m, Staction 3 5–135 m; Staction 4 5–130 m, Staction 5: 5–130 m. To estimate the total annual coccolithophore calcite production we consider only the coccosphere calcite (number of calcifying cells). Uncertainty in the coccolith CaCO3 standing stock estimates is typically ±9% (1σ).
From concentration to annual production
We converted the measured CaCO3 concentrations (i.e. CaCO3 standing stock, CaCO3 biomass) into production rate, using estimates of the turnover time for each group (that is, the typical lifespan of an individual; Table 1): for foraminifera we used a range of 10–30 days90,91,92,93, noting that more slowly reproducing deep-dwelling species make up only a very small fraction of the assemblages in our tows31. For pteropods and heteropods we used a range of 5–16 days (although we note their lifespan may be much longer than this94). Pteropods and heteropod turnover time was calculated as turnover time (days) =1/G, where G is the average instantaneous growth rates expressed as mg Ca deposited (on mg Ca shell)−1 day−1 ref. 57,58. We assume that growth rates do not vary with shell size; this approximation is supported by a previous study77, who found no significant difference in the shell growth rates of small and large sizes of any of the four pteropod species the author examined.
For coccolithophores we used a range of 0.1–1.5 cell division day−1 (1.5–10 days) (Table 1). This range is derived from laboratory field estimates and simulated by a generalized coccolithophore model for equatorial to North Pacific Ocean59. We are aware that cell growth phase differs for small cells with few coccoliths produced during exponential growth phase (normal, rapid division) and larger cells with more coccoliths produced during early stationary phase (slowed cell division).
Given the large range in the turnover rate of coccolithophores, foraminifera, pteropoda, and heteropoda, we apply a probabilistic approach to determine the production rate and propagate the uncertainties in turnover time through to our estimates of total production using a flat probability distribution i.e. for foraminifera there is equal chance of the average lifespan being 10 days as it is 30 days (this highly conservative approach thus results in larger total uncertainties in production rate). The production (mg m-2 day-1) is then given as the CaCO3 standing stock (in mg m-2) divided by the turnover time (days),
Our approach assumes that all of the organisms we sampled are living. This assumption is valid for foraminifera and pteropods as they sink individually, and relatively quickly upon death. For coccolithophores this assumption is valid as we only consider intact coccospheres, which mostly disaggregate quickly upon death. Annual estimates were then calculated by multiplying the daily estimates by 365 accounting for the seasonal bias at the time of sampling using PIC/chlorophyll_a/zooplankton time series (see below).
The data and R code to perform the calculation of CaCO3 production including error propagation and seasonal bias correction (see below) is available at https://doi.org/10.5281/zenodo.7458132.
Correction of production for seasonality and interannual variability
To account for seasonal/inter-annual bias (or specifically, the bias at the time of sampling compared to mean annual production) we use satellite-derived PIC (CaCO3) to correct the coccolithophore production estimates (Fig. S7). The rationale behind this is that although satellites only capture coccolithophore PIC concentrations in the upper few meters of the water column, the relative seasonal/inter-annual changes at the surface should broadly reflect the relative depth integrated seasonal/inter-annual changes in production at depth10,38. We assess the validity of using satellite-derived PIC by regressing satellite-derived PIC (CaCO3) estimates during August 2017 against the surface values of coccolithophore CaCO3 (Fig. S1). Our surface (~5 m depth) estimates of coccolithophore CaCO3 standing stock show a strong correlation with satellite PIC (CaCO3) during August 2017. For each site the seasonal bias factor is calculated as satellite PIC during August 2017)/satellite mean annual PIC (2009–2019). Annual mean coccolithophore production is then given as depth integrated coccolithophore CaCO3 production during August 2017 × 1/seasonal bias factor. As a sensitivity experiment, we repeated this exercise using satellite-derived chlorophyll (see below) instead of PIC, which results in larger estimates of annual CaCO3 production than using PIC.
Unlike coccolithophores, we have no way to directly measure changes in foraminiferal CaCO3 production through time. Instead, we use satellite-derived Chlorophyll A (chlor_a) to correct the foraminiferal production estimates for seasonal/interannual changes (Fig. S9). The rationale here is that the seasonal flux of foraminifera in the North Pacific has been shown to follow primary production31,74, such that we can use relative changes in chlorophyll through time at each site to correct the foraminiferal production estimates. For each site the seasonal bias factor is calculated as chlor_a during August 2017)/mean annual chlor_a (2002–2019). Annual mean foraminiferal production is then given as foraminiferal CaCO3 production during August 2017 × 1/seasonal bias factor.
Again, as we have no way to directly measure changes in pteropod/heteropod CaCO3 production through time, for heteropods and pteropods we refer to the long-term zooplankton data set from ocean stations ALOHA and PAPA to correct pteropod and heteropod CaCO3 production for seasonality. The rational here is that the seasonal changes in pteropod/heteropod abundance should broadly follow the seasonal changes in zooplankton abundance57. We note, that unlike the satellite PIC and chlorophyll estimates used for coccolithophores and foraminifera, this method is not able to account for interannual variability, and only adjusts for the seasonal trend. Based on the multidecadal data set of total zooplankton biomass at St. PAPA66,67 and St. ALOHA (all data and metadata are publicly available at hahana.soest.hawaii.edu/hot/hot-dogs/interface.html) the mean zooplankton biomass in the summer is respectively 2 and 1.2 times greater than the mean annual zooplankton biomass. We extrapolate these values of seasonal bias to each of our sites using latitude. We assume undetectable seasonal variation in pteropod growth rates. If growth rates of pteropods could slightly decrease with temperature, the annual production of aragonite would be less. Given the large assumptions within our method of correcting the pteropod and heteropod production data for seasonal variability, and the possibility of large temporal variability in pteropod abundances43, we also calculate annual pteropod CaCO3 production using the comprehensive pteropod biomass compilation of Bednaršek et al.18 (see below), which has excellent spatial and temporal sampling in the North Pacific (below).
Pteropod biomass and production estimated from MAREDAT
Given the discrepancy observed with previous estimates of pteropod CaCO3 production13 and the sparse spatial and temporal resolution of our net-tow sampling, which unlike the coccolithophore data, cannot be independently verified using satellite data, we also estimate pteropod CaCO3 biomass and production in the North Pacific using the MAREDAT database17. The database has excellent spatial and temporal coverage of pteropod sampling within the North Pacific (Fig. 5). Similar to Bednaršek et al.18, we find no significant trends in biomass by latitude or time of year, so we perform our analysis using all samples spanning the entire North Pacific basin and for all months. We take all estimates of pteropod carbon biomass (reported in the database as mg C m-3) from the upper 250 m of the water column, which we consider as the production layer, with the vast majority of pteropod biomass found within the upper 200m18,95. This results in 1793 discrete observations. We include zero values (120 samples, <7% total) and remove 3σ outliers (16 samples, <1% total). Including these outliers results in unstable Gaussian Kernal densities (see below). We calculate the CaCO3 biomass as the carbon biomass multiplied by the fraction PIC (fPIC), using PIC:POC estimates given in Table 1. We then integrate the standing stock over the production layer, and calculate the production by diving by the turnover time (Table 1) in the same manner as for the C-Disk-IV samples. We calculate the uncertainties via Bootstrapping of the dataset, propagating the uncertainty in PIC:POC and turnover time using Monte-Carlo simulation, using a highly-conservative flat probability distribution for both. Using a different depth range for the production layer (i.e. 0–1000 m) has a negligible effect on our results.
We estimate the probability of pteropod carbon and CaCO3 biomass in the upper 250 m, integrated CaCO3 biomass, and CaCO3 production rate using truncated kernal density:96 this approach truncates the densities below zero, up-weighting the values that are closest to zero, and thus deals better with the highly-skewed dataset containing zero values.
In the North Pacific we find a typical CaCO3 biomass in upper 250 m of 0.5 mg m-3 (0.2–1, 32–68% range), vertically integrated CaCO3 biomass in upper 250 m of 122 mg m-2 (50–269, 32–68% range), daily CaCO3 production of 12 mg m-2 day-1 (5–27, 32-68% range).
Expanding our analyzes the global dataset of ref. 18 (i.e. not only the North Pacific subset described above) we find the global dataset is heavily skewed (Skewness = 13.3; a value above 1 is considered Skewed), such that the mean value reported by ref. 18 is not a useful statistic to describe the dataset. Using the same kernal density method used above for the North Pacific subset we find a typical pteropod CaCO3 biomass of 0.02 mg m-3 globally using data from the upper 250 m, or 0.04 mg m-3, considering all data in the upper 1000 m (Fig. S6). These values are three orders of magnitude smaller than the mean pteropod CaCO3 biomass value reported by ref. 18.
Global satellite-derived PIC
In order to extrapolate the implications of our CaCO3 production estimates globally, we utilize a global climatology of satellite PIC (CaCO3) (Fig. S8;97). Our results indicate satellite PIC (MODIS CaCO3) estimates in the subtropical gyre are biased low compared to vertically integrated CaCO3 production, due to the deeper (mainly coccolithophore) CaCO3 production depth and the thickness of the coccolithophore productive layer (Fig. 2). To account for this bias in the CaCO3 estimates we use a simple regression of total CaCO3 production at our sites against satellite CaCO3,
R2 = 0.71, p < 0.05, standard error = 0.3 mol m2 yr-1 (Fig. S2; Table S8). The deepening of production manifests as the non-zero intercept. Note this relationship is driven by one station with high surface PIC (Station 5). Applying the relationship between total production and satellite PIC (Fig. S2, Table S8) to the global mean surface satellite derived PIC climatology (Fig. S7, Table S8), and integrating globally (weighting by area) results in a total CaCO3 production of 3.1 1014 mol yr-1 (3.7 Pg C yr-1) globally, with the large caveat that this assumes the bias caused by the deepening of production away from the high latitudes scales with surface PIC similarly globally. While we acknowledge this approach is very crude, it nevertheless provides us with a first-order approximation of total global production of CaCO3 implied by our results. The MODIS PIC data are available from NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group97.
Floating sediment trap deployments
The methods and data for the floating sediment traps are given in Dong et al.34 To summarize, at Stations 1–5 (Fig. 1) an array of surface-tethered sediment traps was deployed on a single line; one at 100 m and the other at 200 m depth. Traps were deployed as free-floating arrays for 52 to 78 h. The traps were polycarbonate particle interceptor tubes (PIT) that were 70 cm long, 10 cm diameter (12 tubes per trap) with funnels inserted to guide particles into a Falcon tube attached to the end of the funnel. Falcon tubes were pre-filled with HgCl2 poison in brine solutions to inhibit diffusive loss of poison during deployment. The poison-brine solution was made from seawater collected at 150 m with NaCl added to increase the salinity by ∼5, and sodium borate was added to increase alkalinity by ∼2000 μM (US JGOFS protocol). Samples from six arbitrarily-chosen tubes among the 12 tubes at the same depth were combined and ‘swimmers’ were manually picked out. The samples were then filtered onto a pre-weighed glass fiber filter (Whatman glass microfiber filters, Grade GF/F, 1825–047) and, after being returned to the lab, were reweighed to calculate sinking mass flux. The solid materials on the filters were then collected and analyzed with XRD for mineralogy (aragonite/calcite), and with the Picarro for PIC and total C. We refer the reader to Table 1 in ref. 34 for the values aragonite/calcite ratios and fluxes.
Data availability
The data are given in Tables S1-6 in Supplementary Information and are available on Pangaea (https://doi.org/10.1594/PANGAEA.948508).
Code availability
The data and R code to perform the calculation of CaCO3 production including error propagation and seasonal bias correction is available at https://doi.org/10.5281/zenodo.7458132.
References
Broecker, W. S. & Peng, T.-H. Tracers in the Sea. 690 (Lamont-Doherty Geological Observatory, Columbia University, 1982).
Morse, J. W. & Mackenzie, F. T. Geochemistry of sedimentary carbonates. (Elsevier, 1990).
Ridgwell, A. & Zeebe, R. E. The role of the global carbonate cycle in the regulation and evolution of the Earth system. Earth Planet. Sci. Lett. 234, 299–315 (2005).
Ziveri, P., de Bernardi, B., Baumann, K.-H., Stoll, H. M. & Mortyn, P. G. Sinking of coccolith carbonate and potential contribution to organic carbon ballasting in the deep ocean. Deep Sea Res. Part II: Topical Stud. Oceanogr. 54, 659–675 (2007).
Salter, I. et al. Carbonate counter pump stimulated by natural iron fertilization in the Polar Frontal Zone. Nat. Geosci. 7, 885–889 (2014).
Manno, C. et al. Threatened species drive the strength of the carbonate pump in the northern Scotia Sea. Nat. Commun. 9, 1–7 (2018).
Milliman, J. et al. Biologically mediated dissolution of calcium carbonate above the chemical lysocline? Deep Sea Res. Part I: Oceanographic Res. Pap. 46, 1653–1669 (1999).
Milliman, J. D. Production and accumulation of calcium carbonate in the ocean: budget of a nonsteady state. Glob. Biogeochemical Cycles 7, 927–957 (1993).
Milliman, J. D., Müller, G. & Förstner, U. Recent Sedimentary Carbonates, Part 1 Marine Carbonates. Vol. 19 (Springer‐Verlag, 1974).
Balch, W., Drapeau, D., Bowler, B. & Booth, E. Prediction of pelagic calcification rates using satellite measurements. Deep Sea Res. Part II: Topical Stud. Oceanogr. 54, 478–495 (2007).
Berelson, W. et al. Relating estimates of CaCO3 production, export, and dissolution in the water column to measurements of CaCO3 rain into sediment traps and dissolution on the sea floor: A revised global carbonate budget. Glob. Biogeochem. Cycles 21, GB1024 (2007).
Lee, K. Global net community production estimated from the annual cycle of surface water total dissolved inorganic carbon. Limnol. Oceanogr. 46, 1287–1297 (2001).
Buitenhuis, E. T., Le Quere, C., Bednaršek, N. & Schiebel, R. Large contribution of Pteropods to shallow CaCO3 export. Glob. Biogeochem.Cycles 33, 458–468 (2019).
Broecker, W. & Clark, E. Ratio of coccolith CaCO3 to foraminifera CaCO3 in late Holocene deep sea sediments. Paleoceanogr. 24, PA3205 (2009).
Honjo, S. Fluxes of particles to the interior of the open ocean. Particle flux in the ocean (eds. V. Ittekkot, P. Aschauffer, S. Honjo, & P. Depetris) 91–145 (Wiley, New York, 1996).
Berner, R. Sedimentation and dissolution of pteropods in the ocean. The fate of fossil fuel CO2 in the oceans. Plenum Press, New York, pp 243ą260 (1977)
Buitenhuis, E. et al. MAREDAT: towards a world atlas of MARine Ecosystem DATa. Earth Syst. Sci. Data 5, 227–239 (2013).
Bednaršek, N., Možina, J., Vogt, M., O’brien, C. & Tarling, G. The global distribution of pteropods and their contribution to carbonate and carbon biomass in the modern ocean. Earth Syst. Sci. Data 4, 167–186 (2012).
Kroeker, K. J., Kordas, R. L., Crim, R. N. & Singh, G. G. Meta‐analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 13, 1419–1434 (2010).
Gattuso, J.-P. et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 349, aac4722 (2015).
Subhas, A. V. et al. The dissolution behavior of biogenic calcites in seawater and a possible role for magnesium and organic carbon. Mar. Chem. 205, 100–112 (2018).
Bischoff, W. D., Mackenzie, F. T. & Bishop, F. C. Stabilities of synthetic magnesian calcites in aqueous solution: Comparison with biogenic materials. Geochimica et. Cosmochimica Acta 51, 1413–1423 (1987).
Sulpis, O., Jeansson, E., Dinauer, A., Lauvset, S. K. & Middelburg, J. J. Calcium carbonate dissolution patterns in the ocean. Nat. Geosci. 14, 423–428 (2021).
Schiebel, R., Barker, S., Lendt, R., Thomas, H. & Bollmann, J. Planktic foraminiferal dissolution in the twilight zone. Deep Sea Res. Part II: Topical Stud. Oceanogr. 54, 676–686 (2007).
Gafar, N. A., Eyre, B. D. & Schulz, K. G. A Conceptual Model for Projecting Coccolithophorid Growth, Calcification and Photosynthetic Carbon Fixation Rates in Response to Global Ocean Change. Fronti. Mar. Sci. 4, 433 (2018).
Archer, D. & Maier-Reimer, E. Effect of deep-sea sedimentary calcite preservation on atmospheric CO2 concentration. Nature 367, 260–263 (1994).
Subhas, A. V. et al. Shallow Calcium Carbonate Cycling in the North Pacific Ocean. Global Biogeochemical Cycles 36, e2022GB007388 (2022).
Feely, R. A. et al. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 305, 362–366 (2004).
Orr, J. C. et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437, 681–686 (2005).
Honjo, S. & Okada, H. Community structure of coccolithophores in the photic layer of the mid-Pacific. Micropaleontology 20, 209–230 (1974).
Taylor, B. J. et al. Distribution and ecology of planktic foraminifera in the North Pacific: Implications for paleo-reconstructions. Quat. Sci. Rev. 191, 256–274 (2018).
Feely, R. et al. In situ calcium carbonate dissolution in the Pacific Ocean. Glob. Biogeochem. Cycles 16, 91–12 (2002). 91-91-.
Carter, B. R. et al. Preformed Properties for Marine Organic Matter and Carbonate Mineral Cycling Quantification. Global Biogeochemical Cycles 35, e2020GB00662 (2020).
Dong, S. et al. Aragonite dissolution kinetics and calcite/aragonite ratios in sinking and suspended particles in the North Pacific. Earth Planet. Sci. Lett. 515, 1–12 (2019).
Endo, H. et al. Phytoplankton community responses to iron and CO 2 enrichment in different biogeochemical regions of the Southern Ocean. Polar Biol. 40, 2143–2159 (2017).
Liu, H. et al. Extreme diversity in noncalcifying haptophytes explains a major pigment paradox in open oceans. Proc. Natl Acad. Sci. USA 106, 12803–12808 (2009).
Hopkins, J., Henson, S. A., Painter, S. C., Tyrrell, T. & Poulton, A. J. Phenological characteristics of global coccolithophore blooms. Glob. Biogeochem. Cycles 29, 239–253 (2015).
Balch, W. M., Bowler, B. C., Drapeau, D. T., Lubelczyk, L. C. & Lyczkowski, E. Vertical distributions of coccolithophores, PIC, POC, biogenic Silica, and chlorophyll a throughout the global ocean. Glob. Biogeochem. Cycles 32, 2–17 (2018).
Balch, W. M., Kilpatrick, K., Holligan, P. M. & Cucci, T. Coccolith production and detachment by Emiliania huxleyi (Prymnesiophyceae). J. Phycol. 29, 566–575 (1993).
Daniels, C. J. et al. A global compilation of coccolithophore calcification rates. Earth Syst. Sci. Data 10, 1859–1876 (2018).
Volkov, A. Mean annual characteristics of zooplankton in the sea of Okhotsk, Bering Sea and Northwestern Pacific (annual and seasonal biomass values and predominance). Russian J. Mar. Biol. 34, 437–451 (2008).
Schiel, S. & Cornils, A. Zooplankton abundance measured on Apstein net samples during cruise CISKA2005. Alfred Wegener Institute for Polar and Marine Research, Science for the Protection of Indonesian Coastal Environment (project), Bremerhaven https://doi.org/10.1594/PANGAEA.728970. (2009).
Doubleday, A. J. & Hopcroft, R. R. Interannual patterns during spring and late summer of larvaceans and pteropods in the coastal Gulf of Alaska, and their relationship to pink salmon survival. J. Plankton Res. 37, 134–150 (2014).
Tsurumi, M. et al. Pteropods, eddies, carbon flux, and climate variability in the Alaska Gyre. Deep Sea Res. Part II: Topical Stud. Oceanogr. 52, 1037–1053 (2005).
Bednaršek, N. et al. Integrated Assessment of Ocean Acidification Risks to Pteropods in the Northern High Latitudes: Regional Comparison of Exposure, Sensitivity and Adaptive Capacity. Frontiers in Marine Science 8, https://doi.org/10.3389/fmars.2021.671497 (2021).
Burridge, A. K. et al. Diversity and abundance of pteropods and heteropods along a latitudinal gradient across the Atlantic Ocean. Prog. Oceanogr. 158, 213–223 (2017).
Takahashi, K., Be & Allan, W. H. Planktonic foraminifera: factors controlling sinking speed. Deep Sea Res. Part A 31, 1477–1500 (1984).
Rebotim, A. et al. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosciences 14, 827–859 (2017).
Bruland, K. & Silver, M. Sinking rates of fecal pellets from gelatinous zooplankton (salps, pteropods, doliolids). Mar. Biol. 63, 295–300 (1981).
Ziveri, P., Broerse, A. T., van Hinte, J. E., Westbroek, P. & Honjo, S. The fate of coccoliths at 48 N 21 W, northeastern Atlantic. Deep Sea Res. Part II: Topical Stud. Oceanogr. 47, 1853–1875 (2000).
Guay, C. K. H. & Bishop, J. K. B. A rapid birefringence method for measuring suspended CaCO3 concentrations in seawater. Deep Sea Res. Part I: Oceanographic Res. Pap. 49, 197–210 (2002).
Bishop, J. K. B. & Wood, T. J. Particulate matter chemistry and dynamics in the twilight zone at VERTIGO ALOHA and K2 sites. Deep Sea Res. Part I: Oceanographic Res. Pap. 55, 1684–1706 (2008).
Poulton, A. J. et al. Phytoplankton mineralization in the tropical and subtropical Atlantic Ocean. Glob. Biogeochem. Cycles 20, n/a–n/a (2006).
Emerson, S. et al. Quantifying the flux of CaCO3 and organic carbon from the surface ocean using in situ measurements of O2, N2, pCO2, and pH. Glob. Biogeochem. Cycles 25, GB3008 (2011).
Moore, J. K., Doney, S. C., Glover, D. M. & Fung, I. Y. Iron cycling and nutrient-limitation patterns in surface waters of the World Ocean. Deep Sea Res. Part II: Topical Stud. Oceanogr. 49, 463–507 (2001).
Heinle, M. The effects of light, temperature and nutrients on coccolithophores and implications for biogeochemical models, University of East Anglia, (2013).
Fabry, V. J. Aragonite production by pteropod molluscs in the subarctic Pacific. Deep Sea Res. Part A. Oceanographic Res. Pap. 36, 1735–1751 (1989).
Fabry, V. J. Shell growth rates of pteropod and heteropod molluscs and aragonite production in the open ocean: implications for the marine carbonate system. J. Mar. Res. 48, 209–222 (1990).
Krumhardt, K. M., Lovenduski, N. S., Iglesias-Rodriguez, M. D. & Kleypas, J. A. Coccolithophore growth and calcification in a changing ocean. Prog. Oceanogr. 159, 276–295 (2017).
Poulton, A. J. et al. Coccolithophore dynamics in non-bloom conditions during late summer in the central Iceland Basin (July-August 2007). Limnol. Oceanogr. 55, 1601–1613 (2010).
Buitenhuis, E. T., van Bleijswijk, J., Bakker, D. & Veldhuis, M. Trends in inorganic and organic carbon in a bloom of Emiliania huxleyi in the North Sea. Mar. Ecol. Prog. Ser. 143, 271–282 (1996).
Hayes, C. et al. Global ocean sediment composition and burial flux in the deep sea. Global Biogeochem. Cycles 35, e2020GB006769 (2021).
Cartapanis, O., Galbraith, E. D., Bianchi, D. & Jaccard, S. L. Carbon burial in deep-sea sediment and implications for oceanic inventories of carbon and alkalinity over the last glacial cycle. Clim 14, 1819–1850 (2018).
Timothy, D. et al. Climatology of sediment flux and composition in the subarctic Northeast Pacific Ocean with biogeochemical implications. Prog. Oceanogr. 116, 95–129 (2013).
Karl, D. M. et al. Seasonal-to-decadal scale variability in primary production and particulate matter export at Station ALOHA. Prog. Oceanogr. 195, 102563 (2021).
Frost, B. W. Interannual variation in zooplankton standing stock in the open Gulf of Alaska. From year to year (1983).
Waddell, B. J. & McKinnell, S. Ocean Station” Papa” Detailed Zooplankton Data: 1956-1980. (Science Branch, Department of Fisheries and Oceans, 1995).
Lalli, C. M. & Gilmer, R. W. Pelagic snails: the biology of holoplanktonic gastropod mollusks. (Stanford University Press, 1989).
Bishop, J. K. B., Stepien, J. C. & Wiebe, P. H. Particulate matter distributions, chemistry and flux in the panama basin: response to environment forcing. Prog. Oceanogr. 17, 1–59 (1986).
Oakes, R., Peck, V., Manno, C. & Bralower, T. Degradation of internal organic matter is the main control on pteropod shell dissolution after death. Glob. Biogeochem. Cycles 33, 749–760 (2019).
Young, J. R. Variation in Emiliania huxleyi coccolith morphology in samples from the Norwegian EHUX experiment, 1992. Sarsia 79, 417–425 (1994).
Alldredge, A. L. & Cohen, Y. Can microscale chemical patches persist in the sea? Microelectrode study of marine snow, fecal pellets. Science 235, 689–691 (1987).
Honjo, S. Coccoliths: production, transportation and sedimentation. Mar. Micropaleontol. 1, 65–79 (1976).
Thunell, R. C. & Honjo, S. Seasonal and interannual changes in planktonic foraminiferal production in the North Pacific. Nature 328, 335–337 (1987).
Mallo, M., Ziveri, P., Mortyn, P. G., Schiebel, R. & Grelaud, M. Low planktic foraminiferal diversity and abundance observed in a spring 2013 west-east Mediterranean Sea plankton tow transect. Biogeosciences 14, 2245–2266 (2017).
Fox, L., Stukins, S., Hill, T. & Miller, C. G. Quantifying the Effect of Anthropogenic Climate Change on Calcifying Plankton. Sci. Rep. 10, 1620 (2020).
Wells, F. Seasonal patterns of abundance and reproduction of euthecosomatous pteropods off Barbados, West Indies. Veliger 18, 241–246 (1976).
Schiebel, R. & Hemleben, C. Planktic foraminifers in the modern ocean. (Springer, 2017).
Accornero, A., Manno, C., Esposito, F. & Gambi, M. The vertical flux of particulate matter in the polynya of Terra Nova Bay. Part II. Biological components. Antarct. Sci. 15, 175 (2003).
Davis, C. S. & Wiebe, P. H. Macrozooplankton biomass in a warm‐core Gulf Stream ring: Time series changes in size structure, taxonomic composition, and vertical distribution. J. Geophys. Res.: Oceans 90, 8871–8884 (1985).
Larson, R. Water content, organic content, and carbon and nitrogen composition of medusae from the northeast Pacific. J. Exp. Mar. Biol. Ecol. 99, 107–120 (1986).
Little, W. S. & Copley, N. J. WHOI Silhouette DIGITIZER Version 1.0 Users Guide. (Woods Hole Oceanographic Institution, 2003).
Bednaršek, N., Tarling, G., Fielding, S. & Bakker, D. Population dynamics and biogeochemical significance of Limacina helicina antarctica in the Scotia Sea (Southern Ocean). Deep Sea Res. Part II: Topical Stud. Oceanogr. 59, 105–116 (2012).
Manno, C. et al. Condition of pteropod shells near a volcanic CO2 vent region. Mar. Environ. Res. 143, 39–48 (2019).
Young, J. et al. A guide to extant calcareous nannoplankton taxonomy. J. Nannoplankton Res. 1, 1–125 (2003).
Jordan, R. W., Cros, L. & Young, J. R. A revised classification scheme for living haptophytes. Micropaleontology 50, 55–79 (2004).
Yang, T.-N. & Wei, K.-Y. How many coccoliths are there in a coccosphere of the extant coccolithophorids? A compilation. Br. Phycol. J. 26, 67–80 (2003).
Young, J. R. & Ziveri, P. Calculation of coccolith volume and it use in calibration of carbonate flux estimates. Deep sea Res. Part II: Topical Stud. Oceanogr. 47, 1679–1700 (2000).
Beuvier, T. et al. X-ray nanotomography of coccolithophores reveals that coccolith mass and segment number correlate with grid size. Nat. Commun. 10, 1–8 (2019).
Schiebel, R., Bijma, J. & Hemleben, C. Population dynamics of the planktic foraminifer Globigerina bulloides from the eastern North Atlantic. Deep-Sea. Res.(Part I, Oceanographic Res. Pap.) 44, 1701–1713 (1997).
Berger, W. H. & Soutar, A. Planktonic foraminifera: field experiment on production rate. Science 156, 1495–1497 (1967).
Spindler, M., Hemleben, C., Bayer, U., Bé, A. & Anderson, O. Lunar periodicity of reproduction in the planktonic foraminifer Hastigerina pelagica. Mar. Eco. Prog. Ser. 1, 61–64 (1979).
Jonkers, L., Reynolds, C. E., Richey, J. & Hall, I. R. Lunar periodicity in the shell flux of planktonic foraminifera in the Gulf of Mexico. Biogeosciences 12, 3061–3070 (2015).
Wang, K., Hunt, B., Liang, C., Pauly, D. & Pakhomov, E. Reassessment of the life cycle of the pteropod Limacina helicina from a high resolution interannual time series in the temperate North Pacific. ICES J. Mar. Sci. 74, 1906–1920 (2017).
Mekkes, L. et al. Pteropods make thinner shells in the upwelling region of the California Current Ecosystem. Sci. Rep. 11, 1731 (2021).
Bebu, I. & Mathew, T. Confidence intervals for limited moments and truncated moments in normal and lognormal models. Stat. Probab. Lett. 79, 375–380 (2009).
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group, Moderate-resolution Imaging Spectroradiometer (MODIS) Aqua Particulate Inorganic Carbon Data, (Greenbelt, MD, USA, 2022).
Gafar, N. A., Eyre, B. D. & Schulz, K. G. Particulate inorganic to organic carbon production as a predictor for coccolithophorid sensitivity to ongoing ocean acidification. Limnol. Oceanogr. Lett. 4, 62–70 (2019).
Gannefors, C. et al. The Arctic sea butterfly Limacina helicina: lipids and life strategy. Mar. Biol. 147, 169–177 (2005).
van der Spoel, S. Euthecosomata, a group with remarkable developmental stages (Gastropoda, Pteropoda). (Brill, 1967).
Davis, C. V. et al. Ocean acidification compromises a planktic calcifier with implications for global carbon cycling. Sci. Rep. 7, 2225 (2017).
Kendall, S., Gradstein, F., Jones, C., Lord, O. & Schmidt, D. Ontogenetic disparity in early planktic foraminifers. J. Micropalaeontol. 39, 27–39 (2020).
Schiebel, R. Planktic foraminiferal sedimentation and the marine calcite budget. Glob. Biogeochem. Cycles 16, 3–21 (2002).
NASA Goddard Space Flight Center, O. E. L., Ocean Biology Processing Group. (2018).
Lauvset, S. K. et al. A new global interior ocean mapped climatology: the 1°×1° GLODAP version 2. Earth Syst. Sci. Data 8, 325–340 (2016).
Acknowledgements
We thank the captain and crew of cruise KM1712 on R/V Kilo Moana.Funding was provided by NSF Grants OCE1220600 and OCE1220302 awarded to JA and WB, respectively, MINECO PID2020-113526RB-I00, the Generalitat de Catalunya MERS (#2017 SGR-1588) awarded to PZ and NERC grant NE/N011716/1 awarded to JR. This work is contributing to the ICTA-UAB “Unit of Excellence” (MINECO (CEX2019-000940-M)). We thank Heidi Block, Alasdair Murphy, Rory Abernathy, Joshua Cook, Jurema Domingos and Laura Simon for assistance generating the foraminiferal standing stock data. We acknowledge the Hawaii Ocean Time-series and Ocean Station PAPA programs for data collected at their respective time-series stations.
Author information
Authors and Affiliations
Contributions
P.Z., W.G., J.R., A.S., S.P., J.A., A.W., and W.B. designed the study and sampling strategy, and collected the samples. P.Z., G.A-.O., and C.M. generated the pteropod/heteropod standing stock data. P.Z., M.G., and A.I. generated the coccolithophore standing stock data. W.G., G.A-.O., and J.R. generated the foraminiferal standing stock data. P.Z. and W.G. wrote the paper, with input from all co-authors. All authors contributed to the interpretation and preparation of the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks James Bishop, Kristen Krumhardt and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ziveri, P., Gray, W.R., Anglada-Ortiz, G. et al. Pelagic calcium carbonate production and shallow dissolution in the North Pacific Ocean. Nat Commun 14, 805 (2023). https://doi.org/10.1038/s41467-023-36177-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-023-36177-w
- Springer Nature Limited
This article is cited by
-
Coccolith-calcite Sr/Ca as a proxy for transient export production related to Saharan dust deposition in the tropical North Atlantic
Scientific Reports (2024)
-
Cellular morphological trait dataset for extant coccolithophores from the Atlantic Ocean
Scientific Data (2024)
-
The Contribution of the South Asian Seas (SAS) Program to SDG 14: A Preliminary Assessment of the Targets (1–3)
Thalassas: An International Journal of Marine Sciences (2024)