
Abstract
The blaNDM-1 gene encodes a carbapenemase, New Delhi metallo-β-lactamase (NDM-1), and the ability to produce NDM-1 is spread among Enterobacteriaceae via horizontal gene transfer of plasmids. It has been widely accepted that blaNDM-1 is regulated by a hybrid promoter (PISAba125) consisting of a –10 box from the original blaNDM-1 and a –35 box from ISAba125. However, the conservation of this promoter and the vertical transmission of blaNDM genes by chromosomal integration have not been comprehensively analyzed. We retrieved the region containing the ORF of blaNDM-1 (>95% translated protein identity) and a region 120 bp upstream of the blaNDM-1 start codon from the complete sequence data of Enterobacteriaceae plasmids (n = 10,914) and chromosomes (n = 4908) deposited in GenBank, and the 310 extracted blaNDM genes were analyzed by an in-silico approach. The results showed that most blaNDM genes (99.0%) utilized the promoter, PISAba125. Interestingly, two blaNDM-1 genes from the genus Citrobacter utilized the ISCR1-derived outward-oriented promoters POUT (PISCR1). Furthermore, the insertion of ISAba125 and ISCR1 occurred upstream of the CCATATTT sequence, which is located upstream of the –10 box. We also confirmed that most of the blaNDM genes were disseminated by horizontal gene transfer of the plasmid, but 10 cases of the blaNDM genes were integrated into the chromosome via mobile genetic elements such as IS26, IS150, ISCR1, ICE, and Tn7-like elements. Thus, plasmid-mediated transmission of the PISAba125-blaNDM genes is predominant in Enterobacteriaceae. However, the spread of blaNDM genes with new promoters and vertical dissemination via chromosomal integrations may pose additional serious clinical problems.
Similar content being viewed by others


Introduction
In 2008, Klebsiella pneumoniae, which produces carbapenemase, was detected in a Swedish patient of Indian origin who suffered a urinary tract infection while traveling in New Delhi, India [1]. The new type of carbapenemase was named New Delhi metallo-β-lactamase (NDM-1), and the blaNDM-1 gene encoding NDM-1 was present on a large plasmid that can be introduced into Escherichia coli J53 [1] by conjugation. Indeed, NDM-1-producing bacteria, mainly Klebsiella pneumoniae and E. coli, are spread throughout the world via horizontal gene transfer of the plasmid as carbapenem-resistant Enterobacteriaceae (CRE) [2]. Plasmids are one of the types of mobile genetic elements (MGEs), and bacteria acquire multiple drug resistance (MDR) by incorporating multi-drug-resistance plasmids from other bacteria via horizontal gene transfer. For example, the blaNDM-1 and tetracycline-resistance genes of Citrobacter freundii ZY198 are located on different plasmids, i.e., pZY-NDM1 and pZY-1, respectively (GenBank: CP055250; CP055248). In addition, the plasmids themselves integrate multidrug-resistance genes by incorporating MGEs such as insertion sequences (ISs) and/or transposons [3].
DNA transposons generally contain accessory genes in the cargo region between the two IS elements that are composed of relatively short DNAs, with a short imperfect terminal inverted repeat sequence (IR) on both sides of the DNA ends, and the tnp gene, which encodes transposase [4]. Transposase is involved in target DNA excision and the strand-transfer reaction. Accessory genes within the transposon are involved in drug resistance, heavy metal resistance, metabolism, efflux functions, etc., and the transposition of the transposon into plasmids or chromosomes confers accessory functions on their hosts [5]. The clinical problem is the acquisition of drug-resistance genes via transposon. When transposon is inserted into a chromosome, it may be accompanied by a gene disruption. However, this is not always disadvantageous; indeed, gene disruption may be advantageous in some cases. For example, the loss of the OmpK36 porin gene reduces the permeability of the outer membrane, and this is known to contribute to drug resistance by reducing the uptake of antimicrobial agents into the bacterial cell [6].
In addition, IR-insertion into the plasmid or chromosome is sometime involved in the gene activation. As reported for ISCR1 (Insertion Sequence Common Region 1), the IR of the IS element contains an outward-oriented promoter (POUT), and the short transcripts from POUT repress transposition by forming base-pairs with inwardly transcribed tnp mRNAs [7]. It has been reported that ISCR1 POUT contains two promoters, PCR1-1 and PCR1-2, and both are active and involved in the expression of the blaCTX-M-9 gene [8]. It is widely accepted that the blaNDM-1 promoter is a hybrid promoter generated by the insertion of a right-hand inverted repeat (IRR) of ISAba125 upstream of the blaNDM-1 –10 box [9]. However, the detailed mechanism by which blaNDM-1 and its derivatives with this hybrid promoter spread among Enterobacteriaceae have remained unclear. Furthermore, the spread of the blaNDM family with different promoters by different IS insertions in Enterobacteriaceae has not been elucidated in detail. Here, we retrieved the blaNDM genes from the database containing the complete chromosome and plasmid sequences of Enterobacteriaceae deposited in the GenBank database and performed comprehensive analyses of the blaNDM promoter and chromosome insertion by certain ISs.
Materials and methods
Bacterial strains and growth media
Escherichia coli DH10B was used for plasmid preparation. An E. coli NDM-1 strain (ATCC BAA-2469) was used for the blaNDM-1 cloning. E. coli strains were inoculated in lysogeny broth (LB) and incubated at 37 °C for 16 h with shaking.
Collection of the bla NDM gene family from the Enterobacteriaceae database
The bacterial databases listed in Table 1 were used for the initial blaNDM-1 promoter search. The blaNDM-1 and blaNDM-1 variants (NDM-1 protein with >95% identity, 100% query coverage) containing the region 120 bp upstream of the start codon were retrieved by BLAST and SeqKit [10] from the complete plasmid and genome sequences of Enterobacteriaceae deposited in the GenBank database. Under these conditions, we obtained 300 data points for blaNDM-1 and its variants from a database of 10,914 plasmids and 10 data points from a database of 4908 chromosomes. Data retrieval was done on November 19,2021.
Detection of bla NDM genes integrated into a bacterial chromosome
To investigate the integration of blaNDM into the bacterial chromosome, a BLAST search was carried out using the complete genomic database that consisted of 4908 chromosome data of Enterobacteriaceae; the insertion site of the gene cassette containing blaNDM genes was identified by comparison with the genome sequence of the respective reference strains; and the IS element of the flanking regions of the gene cassettes integrated into the chromosome were detected by searching the ISfinder database (www-is.biotoul.fr). If no IS was detected, we performed an ORF analysis using FramePlot [11] to predict the elements involved in the chromosome integration. The reference strains used for the genome comparison were as follows: K. pneumoniae Kp52.145 (GenBank: FO834906), E. coli K-12 substr. W3110 (GenBank: FO834906), and Enterobacter cloacae GGT036 (GenBank: CP009756).
Construction of plasmids for the bla NDM-1 promoter mapping
A 1.0-kbp fragment of the blaNDM-1 gene containing the entire ORF and its 194-bp upstream region was amplified by PCR with the 5IF-FL and 3IF-FL primers using genomic DNA of the E. coli NDM-1 strain (ATCC BAA-2469) as a template, and the resulting DNA fragment was inserted into the BglII-XhoI sites of plasmid pET-28a (+) using an In-Fusion Cloning system (Takara Bio) to obtain pET-NDM-1_FL. Using the same experimental procedure, pET-NDM-1_166, pET-NDM-1_120, pET-NDM-1_96, pET-NDM-1_71, and pET-NDM-1_42 were also constructed using the primer sets, 5IF-166 and 3IF-FL, 5IF-120 and 3IF-FL, 5IF-96 and 3IF-FL, 5IF-71 and 3IF-FL, and 5IF-42 and 3IF-FL, respectively. The primers used in this study are listed in Table 2.
Antimicrobial susceptibility testing
Minimum inhibitory concentrations (MICs) of meropenem were determined by the microdilution method according to the CLSI guidelines [12] using the Dry Plate Eiken. In brief, bacteria (5 × 104) were inoculated in 100 µl of Mueller–Hinton broth (Becton, Dickinson and Company, Franklin Lakes, NJ) containing meropenem. The tested plates were then incubated at 35 °C for 18–20 h. The MIC value was determined as the lowest concentration of meropenem where no bacterial growth was visibly observed.
Results
Characterization of the minimal promoter region of bla NDM-1
To investigate the blaNDM-1 promoter, we first performed a BLAST search of the 500 bp upstream region of the blaNDM-1 start codon using seven NDM-1-producing strains of E. coli and K. pneumoniae of the GenBank database (Table 1). The results showed that the 194 bp region upstream of the start codon is highly conserved (Fig. 1b), suggesting that the promoter is located within 194 bp upstream of the blaNDM-1 start codon. To analyze the blaNDM-1 promoter, we generated four deletion derivatives using pET-NDM-1_FL, which has a 194 bp upstream region of the blaNDM-1 start codon (Fig. 1a). These plasmids were introduced into E. coli DH10B and grown on LB agar plates containing 100 µg ml−1 of ampicillin. E. coli harboring pET-NDM-1_42 and pET-NDM-1_71 could not grow on the LB agar plates (100 μg ml−1 ampicillin), while E. coli harboring pET-NDM-1_FL, pET-NDM-1_166, pET-NDM-1_120, and pET-NDM-1_96 showed ampicillin-resistant colonies. In order to further validate the promoter activity, the MIC of the plasmid-transfected E. coli DH10B against meropenem was evaluated according to the CLSI guidelines [12]. The MIC of the bacteria harboring pET-NDM-1_FL, pET-NDM-1_166, and pET-NDM-1_120 was 64 μg ml−1. In contrast, the MIC of E. coli harboring pET-NDM-1_96 was 1 μg/ml (Fig. 1). These results suggest that the blaNDM-1 promoter with sufficient activity is located within a 120 bp region upstream of the blaNDM-1 start codon. Furthermore, E. coli harboring pET-NDM-1_96 lacking the –35 box showed reduced resistance to meropenem (Fig. 1). It has been reported that blaNDM-1 is a hybrid promoter consisting of a –35 box derived from the IRR of ISAba125 (Fig. 1b, green highlighted line) and a –10 box derived from the original blaNDM-1 (Fig. 1b, c) [9]. Collectively, our deletion analyses demonstrated that the ISAba125-derived –35 box is indeed essential for full activity of the blaNDM-1 gene.

The ISAba125-derived –35 box in blaNDM-1 is required for sufficient promoter activity. a Domain mapping of the blaNDM-1 promoter. Orange lines indicate an 813 bp ORF encoding NDM-1 carbapenemase. Blue lines indicate the upstream region of the blaNDM-1 start codon. The blaNDM-1 promoter activity was confirmed by growth on LB agar plates containing ampicillin (100 µg ml−1) and MIC against meropenem after introduction of the pET-NDM-1 derivatives into E. coli DH10B. b 194 bp upstream region of the blaNDM-1 start codon. The sequence highlighted in green indicates the IRR of ISAba125. pET-NDM1_96 lacking the –35 box showed significantly reduced meropenem resistance. c Schematic model of the blaNDM-1 promoter. The blaNDM-1 promoter is thought to be a hybrid promoter consisting of a –10 box from the original blaNDM-1 and a –35 box from ISAba125. Red and blue arrows indicate the blaNDM-1 gene and truncated ISAba125, respectively. IRL left inverted repeat, IRR right inverted repeat
The promoter of the bla NDM gene family was highly conserved among Enterobacteriaceae
In the above experiment, we showed that the promoter located within 120 bp is necessary and sufficient for the blaNDM-1 expression. We next analyzed whether this promoter (hereafter called PISAba125) is highly conserved among Enterobacteriaceae of NDM-1 and NDM-1 variants using an in-silico approach as described in the Materials and Methods. We retrieved the 310 full-length blaNDM genes from the Enterobacteriaceae plasmid (n = 10,914) and chromosome (n = 4908) databases. NDM-1 and NDM-1 variants with amino acid substitutions were included, of which NDM-1 and NDM-5 were the most common (Table 3). To investigate the conservation of the blaNDM-1 promoter, we performed multiple sequence alignment by MAFFT (https://mafft.cbrc.jp/alignment/server/) using 310 retrieved blaNDM sequences including the region 120 bp upstream of the start codon. Among them, the promoter PISAba125 (Fig. 1c) was highly conserved in 307 (99.0%) of the 310 blaNDM gene family members (Fig. 2). Although we found one blaNDM-5 with a single base substitution (A to G) within PISAba125, the –35 and –10 boxes were conserved. Therefore, this promoter was also categorized as PISAba125. In contrast, two blaNDM-1 genes were under the control of a different promoter that was generated by insertion of ISCR1 oriIS, as described later. A plasmid found in NDM-5 E. coli (GenBank: CP083875) had two blaNDM-5 genes, and the promoter types were PISAba125 and PNF, respectively. PNF does not have a conserved –35 box. Therefore, the phenotype of NDM-5 may be due to blaNDM-5 being under the control of PISAba125.

Multiple sequence alignment of the blaNDM promoter region. 310 data points (see Table 3) of the 120 bp region upstream of the blaNDM start codon were aligned by MAFFT. Green shaded boxes indicate conserved regions. PISAba125, hybrid promoter derived from ISAba125 insertion; PISCR1, POUT promoter derived from ISCR1 insertion; PNF, non-functional promoter
PISAba125-dependent bla NDM family members are expanded among Enterobacteriaceae
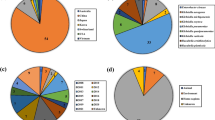
The above analysis revealed that the blaNDM gene family utilizes the highly conserved promoter, PISAba125. Using the 297 data points for the PISAba125-dependent blaNDM genes from a database of 10,914 plasmids, we investigated how the PISAba125-blaNDM spread to Enterobacteriaceae. As shown in Fig. 3, PISAba125-blaNDM-1 and its variant were mostly found in E. coli and Klebsiella spp., but were also detected in the genera Enterobacter, Raoulltera, Salmonella, and Citrobacter. Thus, we confirmed that the PISAba125-blaNDM genes spread to various Enterobacteriaceae via horizontal gene transfer with a plasmid and had the ability to confer carbapenem resistance.

Distribution of the PISAba125-blaNDM family members in Enterobacteriaceae. The horizontal gene transfer of blaNDM-containing plasmids was investigated using 297 data points of PISAba125-blaNDM retrieved from the Enterobacteriaceae plasmid database (n = 10,914). Although some plasmids contain two blaNDM genes or multiple plasmids each contain a blaNDM gene, the bar graph represents the distribution of 297 data points of PISAba125-blaNDM genes
Emergence of the promoter PISCR1 by insertion of ISCR1
Although most of the CRE utilize the PISAba125-blaNDM genes, blaNDM-1 utilizing the promoter POUT derived from the ISCR1 element was found in plasmids possessed by Citrobacter sedlakii [13] and Citrobacter portucalensis (Fig. 4). Interestingly, the ISCR1 element was inserted at the same site in both plasmids, upstream of the –10 box of blaNDM-1 via the conserved sequence CTTCCTATACCC located at the 3′ end of the oriIS of ISCR1 (Fig. 4b: the 232 bp oriIS is highlighted in green). The insertion of the oriIS upstream of the –10 box of the blaNDM-1 promoter generated two oriIS-derived promoters called PCR1-1 (−35 box: TAACG; −10 box: TAAGAT) and PCR1-2 (–35 box: TTCGGA; −10 box: TTTATA) [8]. Thus, two POUT promoters were involved in the expression of blaNDM-1, and we designated the oriIS-derived Pout as PISCR1.

Characterization of the POUT promoter of ISCR1 in Citrobacter spp. a blaNDM-1 using the POUT promoter (PISCR1) derived from the ISCR1 insertion was found in Citrobacter sedlakii and Citrobacter portucalensis plasmids. Red and blue arrows indicate blaNDM-1 and rcr1, respectively. Gray arrows indicate neighboring genes surrounding rcr1 and blaNDM-1 genes. sul1, sulfonamide resistance gene; rcr1, transposase gene; trpF, N-(5’-phosphoribosyl) anthranilate isomerase gene; ble, bleomycin resistance gene; qacΔE, semi-functional derivative of quaternary ammonium compound resistance gene. b The 232 bp region of oriIS is highlighted in green. Two promoters (PCR1-1, PCR1-2) were generated by the ISCR1 oriIS insertion
The bla NDM genes are integrated into a bacterial chromosome
To investigate the blaNDM integration into the bacteria chromosome, a BLAST search was carried out using the complete genomic database that consisted of 4908 chromosome data of Enterobacteriaceae, as described in the Materials and Methods. The chromosome integration of blaNDM genes was detected in E. coli, K. pneumoniae, and Enterobacter cloacae. Although the inserted gene cassettes contained various drug-resistance genes as well as the blaNDM gene, only the blaNDM and the element involved in the genome integration are illustrated here (Figs. 5, 6). Interestingly, the gene cassette integrated into the chromosome contains two blaNDM genes in the E. coli Y5 [14] and K. pneumoniae QD23 [15] strains (Figs. 5, 6). Furthermore, all of the chromosome-integrated blaNDM genes were under the control of PISAba125. In E. coli and Enterobacter cloacae, gene cassettes ranging from 17.5 kbp to 99.4 kbp were surrounded by IS150 or IS26 (Fig. 5). In E. coli ecoli019 and Enterobacter cloacae 174, the blaNDM-1 gene was inserted at the same site on the chromosome, between ompF and asnS.

Transposons are involved in the chromosome-integration of blaNDM in Escherichia coli and Enterobacter cloacae. Red and blue arrows indicate the blaNDM and transposase genes (tnp), respectively. Brown arrows indicate the blaNDM-1-ble fusion gene and the truncated derivative of blaNDM-1, respectively, and both genes appear to be non-functional. Orange boxes indicate inverted repeats (IRL, IRR). Gray arrows indicate adjacent genes of chromosomes located in the transposon insertion. Genes within brackets represent chromosomal genes disrupted by the transposon insertion

blaNDM integrations into chromosomes are diverse in Klebsiella pneumoniae. Red arrows indicate blaNDM genes. Blue arrows indicate related genes involved in the chromosome integration of the blaNDM gene: the transposase gene (tnp), ICE and Tn7-like transposon. Orange boxes indicate insertion sequences involved in the chromosomal integrations. Gray arrows indicate adjacent chromosomal genes located in the transposon insertion. Genes within brackets represent chromosomal genes disrupted by the transposon insertion
In contrast, the manner of the chromosome integration was somewhat complicated in Klebsiella species. In the case of K. pneumoniae 33476891I, the gene cassette containing the blaNDM-1 gene was surrounded by different IS elements, IS26 and ISCR1 (Fig. 6). Interestingly, the integrating and conjugative element (ICE) and Tn7-like transposon were also involved in chromosome integration of blaNDM genes (Fig. 6). Two copies of blaNDM-3 were integrated in a chromosome in K. pneumoniae QD23 via ICE. It has been reported that ICE is inserted via a specific sequence in prfC on the chromosome [16]. Indeed, we found that the chromosome integration of ICE occurred in prfC and was mediated by the typical attR and attL sequences (Fig. 6). The blaNDM genes of K. pneumoniae KP64 and KP67, which are similar in genome composition, were reported to be inserted into the chromosome via IS5 [17]. We found that the site of IS5-insertion into the chromosome was within the Tn7-like transposon (Fig. 6). Thus, the blaNDM genes were distributed not only by horizontal gene transfer via plasmids but also by vertical gene transfer via the integration of various MGEs into the chromosome.
Discussion
In this study, we retrieved 310 blaNDM genes encoding NDM-1 and its variants from the complete plasmid (10,914 data points) and chromosome (4908 data points) databases of Enterobacteriaceae and performed a BLAST search for the blaNDM promoter analysis. Most of the blaNDM promoters were a hybrid-type PISAba125 consisting of a –35 box from ISAba125 IRR and –10 box from the original blaNDM-1. Furthermore, the deletion analysis of PISAba125 revealed that the –35 box was required for the sufficient expression of blaNDM-1. Although in previous studies the PISAba125-blaNDM genes were found to constitute the majority of genes in various CRE, the PISCR1-type of blaNDM-1 was recently detected in a plasmid in Citrobacter sedlakii 3347689II for the first time [13]. This clinical strain was isolated from a Swiss man who had been transferred from a hospital in Macedonia, where he had been hospitalized as a polytraumatized patient, to a hospital in Switzerland. Using an in-silico approach, we detected PISCR1-blaNDM-1 in a plasmid of Citrobacter portucalensis, which was a clinical isolate from Nepal. As shown in Fig. 3, PISCR1 showed the following characteristic features: 1) ISCR1 was located downstream of the qacΔE-sul1 genes; (2) the rcr1-encoding transposase was surrounded by the 379 bp terIS and 232 bp oriIS; and 3) ISCR1 was inserted upstream of the –10 box of blaNDM-1 via a conserved sequence, CTTCCTATACCC, located at the 3′ end of oriIS. The promoter PISCR1 has been reported to be involved in the expression of genes conferring trimethoprim resistance, aminoglycoside resistance, ciprofloxacin resistance, and the abilities to produce class A and C β-lactamase [7]. Among the 310 blaNDM data points, only two blaNDM-1 genes were found to utilize the promoter PISCR1. However, because the above Citrobacter spp. were isolated from different geographical locations, the clinical spread of PISCR1-blaNDM genes should be carefully monitored. In this study, we retrieved 310 data points for blaNDM-1 and its variants (NDM-1 proteins with >95% identity, 100% query coverage) from both plasmid and chromosome databases for promoter analysis. In the process of data extraction, new promoters with different IS-insertions might be detected by reducing the similarity to the NDM-1 protein.
blaNDM genes were generally spread via the horizontal gene transfer of plasmids. We found that a small number of blaNDM genes were integrated into the bacterial chromosome. In E. coli and Enterobacter cloacae, the gene cassette including blaNDM-1 or blaNDM-5 surrounded by the same IS was integrated into the chromosome as transposons or IS26-mediated pseudo-compound transposons [18] (Fig. 5). In the case of K. pneumoniae strains, the manner of chromosome integration was different. The flanking region of the gene cassette had different ISs, ISCR1 and IS26, in K. pneumoniae 3347689I. On the other hand, ICE was involved in the chromosome integration of two copies of blaNDM-3 in K. pneumoniae QD23 [15]. Like plasmids, ICE is a self-transmissible MGE, but it differs from plasmids in that ICE is inserted into the bacterial chromosomes [19]. In ICE, several proteins are involved in the excision, integration, and self-transfer, and we detected the genes encoding these proteins (Fig. 6). In K. pneumoniae KP64 and KP67, blaNDM-1 was reported to be integrated into the chromosome via IS5 [17]. Interestingly, the gene cassette surrounded by IS5 was located within the Tn7-like transposon. The transposon Tn7 is known to recognize the attT7 sequence at a specific site downstream of the glmS gene in the chromosome [20]. Indeed, the transposition of Tn7 occurred downstream of glmS (Fig. 6). Furthermore, tnsABCDE, which is involved in the transposition of T7, and IR sequences specific to Tn7 (Tn7-IR) were also detected (Fig. 6). Thus, the Tn7-like transposon is also involved in the chromosome integration of blaNDM-1 in K. pneumoniae KP64 and KP67.
In this study, we demonstrated that the blaNDM genes utilized a highly conserved promoter, PISAba125. On the other hand, two cases of blaNDM-1 using PISCR1 were detected in the Citrobacter species. Interestingly, ISAba125 and ISCR1 were inserted upstream of the CCATATTT sequence located upstream of the –10 box of the blaNDM gene, suggesting that they function as hotspot sites for promoter acquisition. Furthermore, we found that the blaNDM genes spread not only through the horizontal transfer of plasmids but also through the vertical transfer of genes integrated into the chromosome, which may be a clinical problem in the future.
References
Yong D, et al. Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Ch. 2009;53:5046–54.
Acman M, et al. Role of mobile genetic elements in the global dissemination of the carbapenem resistance gene blaNDM. Nat Commun. 2022;13:1131.
Razavi M, Kristiansson E, Flach C-F, Larsson DGJ. The association between insertion sequences and antibiotic resistance genes. Msphere. 2020;5:e00418–20.
Bourque G, et al. Ten things you should know about transposable elements. Genome Biol. 2018;19:199.
Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol Rev. 2018;31:e00088–17.
Hernández-Allés S, et al. Development of resistance during antimicrobial therapy caused by insertion sequence interruption of porin genes. Antimicrob Agents Ch. 1999;43:937–9.
Toleman MA, Bennett PM, Walsh TR. ISCR elements: novel gene-capturing systems of the 21st century? Microbiol Mol Biol Rev Mmbr. 2006;70:296–316.
Lallement C, Pasternak C, Ploy M-C, Jové T. The role of ISCR1-borne POUT promoters in the expression of antibiotic resistance genes. Front Microbiol. 2018;9:2579.
Partridge SR, Iredell JR. Genetic contexts of blaNDM-1. Antimicrob Agents Ch. 2012;56:6065–7.
Shen W, Le S, Li Y, Hu F. SeqKit: a cross-platform and ultrafast toolkit for FASTA/Q file manipulation. Plos One. 2016;11:e0163962.
Ishikawa J, Hotta K. FramePlot: a new implementation of the frame analysis for predicting protein-coding regions in bacterial DNA with a high G+C content. Fems Microbiol Lett. 1999;174:251–3.
Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing (30th Edition), Clinical and Laboratory Standards Institute, Wayne, PA. M100-ED30 (2020)
Moser A, et al. A patient with multiple carbapenemase producers including an unusual Citrobacter sedlakii hosting an IncC blaNDM-1- and armA-carrying plasmid. Pathogens. Immun. 2021;6:119–34.
Shen P, et al. Detection of an Escherichia coli sequence type 167 strain with two tandem copies of blaNDM-1 in the chromosome. J Clin Microbiol. 2017;55:199–205.
Luo X, et al. Chromosomal integration of huge and complex blaNDM-carrying genetic elements in Enterobacteriaceae. Front Cell Infect Mi. 2021;11:690799.
Wozniak RAF, et al. Comparative ICE genomics: insights into the evolution of the SXT/R391 family of ICEs. Plos Genet. 2009;5:e1000786.
Sakamoto N, et al. Genomic characterization of carbapenemase-producing Klebsiella pneumoniae with chromosomally carried blaNDM-1. Antimicrob Agents Ch. 2018;62:e01520–18.
Varani A, He S, Siguier P, Ross K, Chandler M. The IS6 family, a clinically important group of insertion sequences including IS26. Mob DNA. 2021;12:11.
Ryan MP, Armshaw P, Pembroke JT. SXT/R391 integrative and conjugative elements (ICEs) encode a novel ‘Trap-Door’ strategy for mobile element escape. Front Microbiol. 2016;7:829.
Kuduvalli PN, Rao JE, Craig NL. Target DNA structure plays a critical role in Tn7 transposition. EMBO J. 2001;20:924–32.
Funding
This work was supported in part by grants from the Ministry of Education, Culture, Sports, Science and Technology and the Japan Society for the Promotion of Science (KAKENHI) nos. 19K07542 (to AA), 20K07485 (to AK) and by AMED under Grant Number JP20nk0101552 (to AA) and JP22nk0101587 (to AA). The funders had no role in the study design, date collection or analysis, the decision to publish, or the preparation of manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kikuchi, Y., Matsui, H., Asami, Y. et al. Landscape of blaNDM genes in Enterobacteriaceae. J Antibiot 75, 559–566 (2022). https://doi.org/10.1038/s41429-022-00553-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41429-022-00553-3
- Springer Japan KK