
Abstract
Cancer-associated fibroblasts (CAFs) are activated fibroblasts that constitute the major components of tumor microenvironment (TME) and play crucial roles in tumor development and metastasis. Here, we generated fibroblast-specific inducible focal adhesion kinase (FAK) knockout (cKO) mice in a breast cancer model to study potential role and mechanisms of FAK signaling in CAF to promote breast cancer metastasis in vivo. While not affecting primary tumor development and growth, FAK deletion significantly suppressed breast cancer metastasis in vivo. Analyses of CAFs derived from cKO mice as well as human CAFs showed that FAK is required for their activity to promote mammary tumor cell migration. We further showed that FAK ablation in CAFs decreased exosome functions to promote tumor cell migration and other activities, which could contribute to the reduced metastasis observed in cKO mice. Lastly, profiling of miRs from CAF exosomes showed alterations of several exosomal miRs in FAK-null CAFs, and further analysis suggested that miR-16 and miR-148a enriched in exosomes from FAK-null CAFs contribute to the reduced tumor cell activities and metastasis. Together, these results identify a new role for FAK signaling in CAFs that regulate their intercellular communication with tumor cells to promote breast cancer metastasis.
Similar content being viewed by others
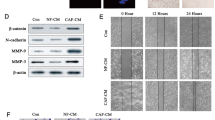
Introduction
Cancer metastasis is an inefficient multistep process requiring the disseminated tumor cells to adapt and survive at a foreign microenvironment in distant tissues. During this process, intercellular communication is critical, especially between tumor cells and the surrounding tumor microenvironment (TME), to create a tumor-favoring niche that allows tumor growth and colonization. Cancer-associated fibroblasts (CAFs) are activated fibroblasts that constitute the major components of TME and play crucial roles in tumor development and metastasis [1, 2]. In human breast cancer, increased CAFs have been shown to be associated with aggressive disease, recurrence, and drug resistance [3]. However, the role and mechanisms of CAF contribution to breast cancer metastasis is still poorly understood. It remains as a significant challenge to elucidate the intracellular signaling pathways in CAFs that regulate intercellular communication between CAF and tumor cells critical for different stages of cancer metastasis.
Exosomes are a class of extracellular vesicles generated in multivesicular bodies and released from different cells [4, 5]. They range from 50 to 150 nm in size and contain bioactive molecules such as proteins, lipids, and nucleic acids, including microRNAs (miRNA). Previous studies showed that cancer cells secrete exosomes to regulate recipient cell functions in TME to facilitate cancer metastasis and progression [6, 7]. Exosomal miRNAs derived from cancer cells contribute to increased tumor angiogenesis and metastasis by affecting both local TME and distal organs via circulation [8]. A recent study suggested that CAF-derived exosomes can promote stemness properties, EMT phenotype, and anchorage-independent growth of tumor cells [9]. However, in contrast to extensive studies of exosomes and miRNAs from cancer cells, less is known about the functions of CAF-derived exosomes through delivering miRNA contents to recipient cancer cells to promote cancer metastasis.
Focal adhesion kinase (FAK) is a non-receptor tyrosine kinase that mediates signal transduction by integrins and other cell surface receptors to regulate cell adhesion, migration, survival, proliferation and differentiation in a variety of cells [10,11,12,13,14]. FAK overexpression in breast cancer correlates with poor prognosis. Previous studies have strongly implicated FAK in the development and progression of breast and other cancers [15, 16]. Several groups, including us, have shown that the deletion of FAK in mammary epithelial cells suppresses the tumor formation and progression in mouse models of breast cancer [17,18,19,20]. We and others have also shown that FAK signaling play important roles in endothelial cells to promote embryonic and cancer angiogenesis and drug resistance [21,22,23]. However, in contrast to the wealth of knowledge on FAK in tumor cells, much less is known about its potential role in tumor stroma, other than a role for endothelial FAK in tumor angiogenesis.
In this study, we generated and analyzed fibroblast-specific inducible FAK knockout mice in a breast cancer model to investigate the potential role and mechanisms of FAK in CAF to promote breast cancer metastasis in vivo. Our studies showed that FAK ablation in fibroblasts significantly reduced breast cancer metastasis by impairing the abilities of CAF-derived exosomes to promote cancer cell migration and other activities at least in part due to altered exosomal miR-148a and miR-16. These results provide significant insights into the multiple mechanisms of FAK signaling in TME besides their essential role in mammary tumor cells, which may contribute to the future development of novel therapies for breast cancer targeting FAK.
Results
FAK ablation in fibroblasts inhibits breast cancer metastasis in vivo
To study the potential role of FAK signaling in CAFs to promote breast cancer development and metastasis in vivo, we crossed MMTV-PyMT;FAKf/f mice [18] with Col1a2-CreER transgenic mice that express tamoxifen (TAM)-activated Cre recombinase in CAFs [24,25,26] to produce MMTV-PyMT;FAKf/f;Cre mice. They (and all strains used in this study) were backcrossed to FVB/N genetic background (syngeneic to the MMTV-PyMT mice used throughout the study) for seven or more times (>99%, or virtually pure FVB/N) to eliminate potential influence of genetic background on phenotypes. Cohorts of female MMTV-PyMT;FAKf/f;Cre mice were treated with vehicle (designated as Ctrl mice) or TAM (designated as cKO mice after induced FAK deletion) at 5 weeks of age every other day for three times [2 mg each time] (Fig. 1a). Unlike previous studies deleting FAK in tumor cells [18], ablation of FAK in CAFs did not affect endogenous mammary tumor development in cKO mice relative to Ctrl mice (Fig. 1b). Further, primary tumor growth was comparable in cKO and Ctrl mice (Fig. 1c). Interestingly, however, we observed that cKO mice developed metastasis at a significantly reduced frequency than Ctrl mice at 4 weeks after TAM treatments (Fig. 1d, e).

a Experimental design scheme for induction of FAK deletion in transgenic mice. b Kaplan–Meier tumor free curve of Ctrl (n = 26) and cKO (n = 30) mice. c Endogenous primary tumor growth curves of Ctrl (n = 9) and cKO (n = 9) mice. d H&E staining of lungs from Ctrl and cKO mice at end point (4 weeks after Tamoxifen induction). Scale bar, 2 mm. e Quantification of metastatic nodules per lung section in Ctrl (n = 9) and cKO mice (n = 9). f Immuno-blots showing levels of FAK, SMA, Pan-CK, and Actin in primary tumor cells and fibroblasts. Quantification of respective protein levels normalized against Actin in g tumor cells and h CAFs described in f. i Immuno-blots showing levels of FAK, SMA, and GAPDH in lung CAFs isolated from Ctrl and cKO mice. Error bars represent mean ± s.e.m. *p ≤ 0.05. j Quantification of respective protein levels normalized against GAPDH in lung CAFs described in i.
To verify FAK deletion in CAFs, tumor cells and CAFs were prepared from mammary tumors in cKO and Ctrl mice, as described previously [9, 18, 27]. Western blot analysis of lysates showed FAK deletion specifically in CAFs, but not mammary tumor cells, from cKO mice (Fig. 1f–h). We next isolated CAFs from lungs of these mice by FACS using PDGFRβ as a marker [28], and showed that FAK was deleted in cKO CAFs compared with CAFs from Ctrl mice (Fig. 1i, j). Lung epithelial cells from cKO and Ctrl mice showed comparable levels of FAK (data not shown). Western blotting analysis using alpha-smooth muscle actin (a-SMA) as a marker for fibroblasts activation (i.e., CAFs) showed comparable levels of a-SMA in fibroblasts from cKO and Ctrl mice, suggesting that FAK deletion did not affect CAF formation in cKO mice. Together, these results suggest that FAK deletion in CAF suppressed breast cancer metastasis, although it did not affect primary mammary tumor development and growth.
FAK signaling in CAFs is required for their promotion of tumor cell migration in vitro
To further evaluate a role of FAK signaling in CAFs to promote human breast cancer metastasis, we prepared human CAFs and examined their effects on human breast cancer cell migration in vitro. First, WI-38 human fibroblasts were incubated with conditioned media (CM) from several human breast cancer cells, and examined by western blotting using antibody for a-SMA (Fig. 2a, b). We found that CM from MCF-7, HCC-1954, and MDA-231 breast cancer cells, but not MCF10A (a normal mammary epithelial cell line), induced WI-38 cells to a CAF-like state with increased expression of a-SMA. FAK phosphorylation was also increased in WI-38 cells treated with CM from these breast cancer cells, supporting a potential role for FAK signaling in CAFs. We next analyzed the ability of the treated WI-38 cells to promote migration of breast cancer cells, which is an important characteristic of metastasis. We found that WI-38 cells treated with HCC-1954 and MDA-231 CM promoted the migration of these two breast cancers relative to WI-38 cells not treated (NT) with the CM, while those treated with MCF-7 CM increased its migration slightly (Fig. 2c, d).

a Immuno-blots showing levels of SMA, FAK-pY397, FAK, and Actin in WI-38 human fibroblasts that are not treated (NT) or treated with conditioned media from respective human breast cancer cells. b Quantification of respective protein levels normalized against total FAK in WI-38 fibroblasts described in a. c Illustration of coculture transwell migration assay. d Quantification of migrated cells in transwell migration assays of human breast cancer cells that were educated by fibroblasts (not treated [NT] or treated with respective conditioned media from tumor cells). e Immuno-blots showing levels of FAK, SMA, and GAPDH in WI-38 cells treated with shCtrl or shFAK to knockdown FAK. f Quantification of transwell migration assays of MDA-MB-231 tumor cells to WI-38 cells treated with shFAK and/or MDA-231 conditioned medium for 72 h. Levels of FAK knockdown in WI-38 cells are indicated by immune-blots. Representative images (g) and quantification (h) of migrated cells in transwell migration assays of PyMT tumor cells to primary fibroblasts purified from lungs of WT, Ctrl, or cKO mice. Scale bar, 50 μm. Error bars indicate mean ± SEM. *p < 0.05, **p < 0.01.
To directly examine the role of FAK signaling in CAF, we generated WI-38 cells with stable FAK knockdown using lentiviral vectors for FAK shRNA and examined their ability to promote the migration of MDA-231 cells following treatment with MDA-231 CM. We found that FAK knockdown did not change the activation of WI-38 cells by MDA-231 CM, as measured by a-SMA expression (Fig. 2e). However, FAK knockdown abolished the ability of MDA-231 CM treated WI-38 cells to promote the migration of breast cancer cells (Fig. 2f). Consistent with these results using WI-38 cells, CAFs from Ctrl mice, but not those from cKO mice, showed greater ability to promote PyMT tumor cell migration, relative to lung fibroblasts from WT mice (Fig. 2g, h). Collectively, these results suggest that FAK signaling in CAFs is important for their ability to stimulate tumor cell migration, which were consistent with the decreased metastasis in cKO mice in vivo.
FAK deletion in CAFs results in defective exosomes for promoting tumor cell migration and other functions
Exosomes are small micro vesicles containing functional biomolecules like proteins and RNAs, which can transfer between the donor cells and the recipient cells [4, 5]. To explore whether FAK signaling in CAFs regulates tumor cells through exosomes, we isolated and examined exosomes from Ctrl and cKO CAFs by ultracentrifugation of the CM, as described previously [29, 30]. We found a comparable number of exosomes in CAFs from Ctrl and cKO mice (Fig. 3a). However, both protein and RNA contents were decreased in exosomes from cKO CAFs relative to those in Ctrl CAFs (Fig. 3b, c), suggesting that FAK deletion in CAFs reduced the amount of protein or RNA in exosomes. On the other hand, size distributions of exosomes isolated from CAFs of cKO and Ctrl mice were similar (Fig. 3d). Analyses of the exosome preparations by western blotting showed that both Ctrl and cKO exosomes are enriched in various exosome proteins; including CD81 and Alix at comparable levels in both exosomes, and CD63 and integrin β4 at reduced levels in exosomes from cKO CAFs relative to exosomes from Ctrl CAFs (Fig. 3e, f). FACS analysis using CD63 as a marker verified purities of >99% in both exosomes from Ctrl and cKO CAFs (Fig. 3g). Electron microscopy further confirmed purity of exosome preparations, showing exosomes ranging between 70 and 170 nm (Fig. 3h), which is comparable to estimates by Nanosight (see Fig. 3d).

Quantification of a numbers of exosome generated per million of cells (volume of preparation), b protein content (mg/ml) per volume of preparation, and c RNA content per volume of preparation of exosomes from purified primary lung fibroblasts. d Quantification of exosome size distribution by nanosight. e Immuno-blots showing levels of exosome markers Integrinβ4, CD63, CD81, and Alix using purified exosomes from primary lung fibroblasts of Ctrl or cKO mice. f Quantification of respective protein levels from purified exosomes normalized against GAPDH as described in e. g CD63 FACS analysis of purified exosomes. Exosomes without CD63 primary antibody incubation were utilized as negative control. h Representative images of electron microscopic analysis of exosomes derived from the primary lung fibroblasts. Scale bar, 100 nm. Error bars indicate mean ± SEM. **p < 0.01.
We next examined the effect of CM from Ctrl CAFs with or without exosomes on PyMT tumor cell migration using wound closure assay. We found that while the Ctrl CM significantly increased tumor cell migration, this activity was much lower after the removal of exosomes (Fig. 4a, b). CM from cKO CAFs also promoted tumor cell migration but to a less extent, and removal of exosomes from the media did not affect this activity (Fig. 4b). Interestingly, CM from cKO CAFs with exosome depletion showed greater activity to promote tumor cell migration than CM from Ctrl CAFs without exosomes (Fig. 4b), suggesting Ctrl exosomes are primarily responsible for the activity in CM from Ctrl CAFs, whereas the activity over NT in CM of cKO CAFs are likely due to other factors (e.g., altered chemokine compositions). Indeed, adding back Ctrl CAF exosomes, but not cKO CAF exosomes, to the exosome-depleted CM promoted tumor cell migration in a dose-dependent manner (Fig. 4c). Purified exosomes from Ctrl CAFs, but not those from cKO CAFs, also increased tumor cell colony formation in soft agar (Fig. 4d). Analysis of human CAFs also showed that the knockdown of FAK by shRNA reduced the ability of CM from these cells to promote tumor cell migration and proliferation (Fig. S1). Previous studies suggested important roles of breast cancer stem cells (BCSCs) in metastasis [31,32,33,34], we therefore also examined the effect of CAF exosomes on BCSCs. We found that exosomes from Ctrl CAF increased tumor sphere formation of PyMT primary tumor cells compared with cKO exosomes or no addition (Fig. 4e). Consistent with these in vitro data, cKO mice showed a decreased amount of BCSCs than Ctrl mice, as measured by ALDH assays (Fig. 4f). Taken together, these results suggest that FAK signaling in CAFs regulates the generation of exosomes and their functions to promote breast cancer metastasis.

a Representative images of wound healing assay of PyMT cells treated with Ctrl or cKO CAF conditioned media with or without exosome removal for 18 h. Scale bar represents 200 μm. b Quantification of wound healing assay as described in a (n = 3). c Wound healing assay of PyMT cells treated with exosome at various doses (0–10 µg) for 18 h (n = 3). Colony formation (d), sphere formation (e), and quantification of ALDH + cells (f) in PyMT cells that are not treated (NT) or treated with 10 µg of purified exosomes from Ctrl or cKO CAFs. Error bars indicate mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Regulation of miR-16 and miR-148a in CAF exosomes by FAK contributes to altered ability of CAFs to affect tumor cell activity and metastasis
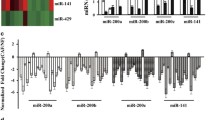
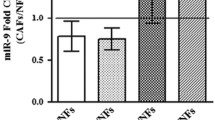
MiRNAs encapsulated in exosomes are abundant and play important roles in intercellular communications [7, 8]. We therefore hypothesized that FAK deletion in CAFs alter miRNAs in exosomes to abolish their activity to promote tumor cell functions and metastasis. To identify the specific miRNAs involved, we performed miRNA sequencing of CAF-derived exosomes to generate miRNA profiles from Ctrl and cKO mice (n = 3 for each). Comparative analysis of miRNA profiles identified three decreased miRNAs and four increased miRNAs in cKO CAF-derived exosomes relative to those from Ctrl mice (Fig. 5a). Using additional preparations of CAF-derived exosomes of Ctrl and cKO mice, qRT-PCR further confirmed reduced levels of miR-34b, miR-409, and miR-494 as well as increased amount of miR-16, miR-148a, and miR-326 in cKO CAF-derived exosomes (Fig. 5b), suggesting that these exosomal miRs may mediate CAF regulation of mammary tumor metastasis in cKO mice.

a Heatmap showing differentially expressed miRs in exosomes from Ctrl or cKO CAFs (n = 3 each). b Quantitative-PCR analysis of miR levels in exosomes from Ctrl and cKO CAFs (n = 3 each). c Quantitative-PCR analysis of miR levels in exosomes from WI-38 fibroblasts transduced with shCtrl or shFAK and educated by MDA-MB-231 cells. Quantitative-PCR analysis of d miR148a target genes and e miR16 target genes in MDA-MB-231 cells that were treated with exosomes from shCtrl or shFAK transduced WI-38 fibroblasts. f Transwell migration assays for PyMT cells treated with exosomes from Ctrl or cKO lung CAFs, along with specified miRNA inhibitors. NC denotes scrambled control oligo. g EdU incorporation assay for PyMT cells treated with exosomes from Ctrl or cKO lung CAFs, along with specified miRNA inhibitors. Error bars indicate mean ± SEM. *p < 0.05, **p < 0.01.
We next prepared exosomes from human WI-38 cells with or without FAK knockdown (see Fig. 2e) that had been treated MDA-231 CM and examined the levels of these miRs. MiR-16 and miR-148a showed increased expression in WI-38 cells with FAK knockdown, consistent with results in mouse CAFs, although miR-326 was not increased after FAK knockdown (Fig. 5c). Surprisingly, we did not find the decreased expression of miR-409 or miR-494 in WI-38 cells after FAK knockdown, and miR-34b was not detected in WI-38 cells with or without FAK knockdown. Similar analysis in WI-38 cells treated with MCF-7 CM showed that FAK knockdown did not change the levels of any of these miRs (Fig. S2), which is consistent with the observation that WI-38 cells treated with MCF-7 CM did not affect migration of these cells (see Fig. 2d). These results further support that exosomal miR-16 and/or miR-148a play a role in mediating CAF regulation of recipient tumor cells. Indeed, both miR-16 and miR-148a have been reported to act as tumor suppressive miRs in different cancers including breast cancer [35,36,37]. Thus, it is possible that exosomes from CAFs lacking FAK (either from cKO mice, or human WI-38 cells with FAK knockdown) and enriched with miR-16 and/or miR-148a inhibit various tumor cell activities compared with exosomes from Ctrl CAFs (see Figs. 4c–f and S1).
To further evaluate this notion, we examined the expression of a series of putative targets of miR-16 and miR-148a in the recipient MDA-231 cells treated by exosomes from human CAFs with or without FAK knockdown. We found that miR-16 targets, CCNE1 and TWIST1, as well as miR-148a targets, WNT1 and WNT10B, were significantly decreased in tumor cells treated with exosomes from CAFs with FAK knockdown compared with those treated with exosomes from Ctrl CAFs (Fig. 5d, e). Moreover, decreased levels of Twist1 and Ccne1 proteins were found in tumor cells treated with exosomes from cKO CAFs vs those from Ctrl CAFs (Fig. S3). We next used inhibitors against miR-16 and miR-148a to further validate their roles in decreasing mammary tumor cell activities. As expected, transfection of miR-148a and miR-16 inhibitors restored the levels of miR-148a targets CCNE1 and Twist1 and miR-16 target Wnt10b, respectively, in tumor cells treated with exosomes from cKO CAFs (Fig. S4). Importantly, transfection of either of these inhibitors reversed the inhibition of tumor cell migration and proliferation by exosomes from cKO CAFs (Fig. 5f, g). Moreover, transfection of these two inhibitors into cKO CAFs also reduced the levels of corresponding miR-148a and miR-16 in exosomes derived from these cells (Fig. S5A), as expected, and these exosomes induced more tumor cell migration compared with exosomes from cKO CAFs treated with scrambled control (Fig. S5B). These results confirmed the efficacy of the inhibitors, and are consistent with the idea that the increased amount of miR-16 and miR-148 in exosome from cKO CAF is responsible for the lower migration of tumor cells compared with those treated with Ctrl CAF exosomes. Lastly, miR-148a and miR-16 inhibitors did not affect the base line proliferation of tumor cells (i.e., those without exosome treatment) (Fig. S6), supporting that they acted on reserving the effect of corresponding miRNAs enriched in exosomes from cKO CAFs which were delivered to tumor cells. Together, these results suggested that miR-16 and miR-148a enriched in exosomes from FAK-null CAFs play a key role in decreasing mammary tumor cell functions and metastasis (Fig. S7).
Discussion
Cancer is increasingly recognized as a disease involving abnormalities in both tumor cells and other surrounding cells within TME as well as altered communications between the cells [38]. It is well established that malignant cancer cells can activate multiple mechanisms to “educate” both local and distant fibroblasts into CAFs, which in turn influence various cancer cell behavior to promote cancer metastasis to distant organs. By creating a fibroblast-specific inducible FAK knockout in a mouse model of breast cancer as well as knocking down FAK in human CAFs, we showed a critical role for FAK signaling in CAFs for metastasis. Therefore, unlike some other key signaling molecules that could play opposing roles in cancer vs stromal cells for cancer progression and metastasis [39, 40], FAK’s function in CAFs as well as its pro-tumorigenesis role in tumor and endothelial cells [17,18,19,20,21,22,23] highlight significant potential for FAK inhibitors for systemic cancer therapy by attacking both tumor cells and multiple cells in TME crucial for cancer metastasis.
Besides well-established roles of FAK in mediating intracellular signaling in tumor and other cells, recent studies suggested new functions for FAK in the regulation of TME through affecting intercellular communications. FAK in tumor cells has been implicated in creating an immune-suppressive TME through inflammatory chemokine production and tumor-associated regulatory T cells recruitment [41, 42]. Other studies suggested a role for FAK in regulating the composition of the fibrotic and immune-suppressive niche in pancreatic tumor model [43, 44]. Our results reveal new FAK functions in fibroblasts required for their activation to CAFs in response to tumor cells, and more importantly the ability of CAFs to promote breast tumor cell migration. FAK can regulate diverse signaling pathways through its phosphorylation of and interaction with multiple other molecules. Using FAK mutant knockin alleles in breast cancer models, we previously showed that FAK kinase activity was required for some, but not other, subtypes of breast cancer [45], and that FAK interaction with endophilin A2 specifically affects breast cancer stem cell pool but not bulk tumor cells [46]. It will be interesting to determine the role of FAK kinase activity and specific downstream pathways in its regulation of CAF-derived exosomal miRNAs to promote breast cancer metastasis using similar strategies.
Exosomes have emerged as an important mediator of intercellular communication between various cells. Our findings are consistent with a number of previous studies showing the promotion of breast cancer cell migration and metastasis by CAF-derived exosomes [6, 47]. Moreover, these data provide strong support for a role of exosomal miRNAs in mediating the reduced metastasis upon FAK ablation in CAFs of cKO mice, although we could not exclude potential contribution by changes in exosomal RNAs and/or proteins from cKO CAFs (e.g., decreased amount of integrin β4 and/or CD63, see Fig. 3e, f). We found that FAK ablation in CAFs increased levels of exosomal miR-148a, which decreased WNT1 and WNT10B expression in the recipient cancer cells. These data provided further support the previous observation that CAF exosomes stimulate Wnt-PCP signaling to drive breast cancer cell invasive activities [6]. Our studies also showed increased levels of exosomal miR-16 from FAK-null CAFs that decreased other genes CCNE1 and TWIST1 to reduced cancer cell proliferation and EMT. Besides the increased tumor suppressive miR-148a and miR-16, we also found reduced levels of oncogenic miRs-34b, 409 and 494 in CAF-derived exosomes of cKO mice. However, we did not detect similar decreases of these oncogenic miRs in human CAFs following FAK knockdown, which could be due to incomplete ablation of FAK in these cells by knockdown approach. Nevertheless, it is conceivable that the inhibition of one or more of the oncogenic miRs will reduce the ability of CAF exosomes to promote metastasis in cKO mice, possibly in conjunction with the increased levels of tumor suppressive miR-16 and/or miR-148a, as discussed above. Indeed, one caveat in support of the roles of increased miR-16 and miR-148a in cKO CAFs is the reduced amount of cKO exosomes compared with that from Ctrl CAFs (thus canceling out the net effect of the enriched miRs on per exosome basis). However, multiple miRs may act in an integrated manner to influence recipient tumor cell behavior, and in such a case, both the reduced oncogenic miRs and the enriched suppressive miRs from the overall reduced exosome amount in cKO CAFs may be involved.
Cancer metastasis is a complex, multiple stage process including tumor cell intravasation into the blood or lymphatic vasculature at the primary tumor site, survival in the blood/lymphatic vessels, and colonization and growth at the metastatic site such as the lung. Interestingly, we found that FAK ablation in CAFs inhibited breast cancer metastasis, but it had no apparent effects on the development and growth of primary tumors. Thus, it is possible that defective CAFs at the lung metastatic sites could be responsible for the reduced colonization and/or growth at the tumor cells to inhibit metastasis observed in cKO mice. Alternatively, our data showing that FAK ablation diminished the ability of CAFs to promote tumor cell migration are also consistent with the possibility that cKO CAFs affected metastasis by reduced support for tumor cell intravasation despite not affecting the primary tumor development or growth. Future studies will be directed towards examining these possibilities to further dissect the mechanism involved. We also noted that FAK-null CAF exosomes with enriched miR-16 and miR-148a inhibited the proliferation of tumor cells in vitro, although primary tumor growth was not affected in cKO mice in vivo. It is possible that primary tumors developed compensatory mechanisms so that their growth was less dependent on possible effects of FAK-null CAFs in vivo. Another, not mutually exclusive possibility is that tumor cells metastasized to the lungs have a more stringent requirement for their microenvironment including CAFs to support their growth to form lung nodules (and FAK deletion in CAFs compromising such supportive function), which is reflected in the in vitro assays. Lastly, though less likely, it is also possible that FAK signaling is preferentially more important in lung CAFs (where they were isolated for the in vitro assays) than CAFs associated with primary tumors in mammary gland. Future studies will be necessary to gain additional mechanistic insights using our unique mouse models and reagents derived from them. Likewise, it will also be interesting to evaluate potential differential changes in specific subsets of CAFs upon FAK deletion and their contributions to the reduced metastasis in vivo, given recent studies showing multiple subsets of CAFs that could support or suppress tumor progression [3, 48, 49]. Lastly, defective CAFs upon FAK ablation could modulate other TME components such as immune cells and ECs to impact indirectly on cancer metastasis. The availability of the fibroblast-specific cKO mice described here, along with FAK cKO mice in ECs [50] and mammary tumor cells [18] will offer appropriate models to fully understand the complex mechanism of FAK signaling in the regulation of metastasis in vivo.
In summary, our studies revealed a new function for FAK in intercellular communication between CAFs in TME and mammary tumor cells important for cancer cell migration and metastasis. FAK ablation in CAFs impaired their ability to promote cancer cell migration and other activities due to alterations in exosomal miRs. These findings provide new insights into the complex role of FAK signaling in TME and cross-talk with mammary tumor cells and further highlight the potential of targeting FAK in the development of systemic and effective therapy for advanced and metastatic breast cancer.
Materials and methods
Tumor mice
MMTV-PyMT;FAKf/f mice have been described previously [18], and Col1a2-creERT mice obtained from Jackson Lab. The tumor cohorts were maintained on a FVB/N genetic background after extensive back-crossing (7× or more). Mice were housed and handled according to local, state, and federal regulations, and all experimental procedures were carried out as per the guidelines of the Institutional Animal Care and Use Committee at the University of Cincinnati. Tumors were measured by calipers and volume was calculated as (1/2)(length)(width)2. To induce deletion, mice were treated with 2 mg tamoxifen for three times every other day, intraperitoneally.
Cell culture and treatment
MDA-MB-231, MCF-7, and WI-38 cells were obtained from ATCC. These cells were cultured in DMEM with 10% fetal bovine serum. Cells were routinely tested for mycoplasma contamination. Murine fibroblasts were prepared from primary tumors and lung tissues, as described previously [9, 27]. Isolated fibroblasts were cultured in DMEM/F12 supplemented with 10% FBS, insulin (10 ng/ml) and EGF (10 ng/ml). Wound healing, transwell invasion, and sphere-forming assays were carried out as described previously [18]. For transwell assays, 10% serum media was used as a chemoattractant in the lower chamber while tumor cells were seeded in the upper chamber with 0.1% FBS and tumor cells were assayed for 16 h. For coculture transwell assays (Fig. 2c, d), tumor CM pre-educated WI-38 cells were seeded in the lower chamber with respective tumor CM before plating tumor cells in the upper chambers 2 h later. For CM, cells were cultured in 10% serum media and media was collected after 72 h. For treatment with miRNA inhibitors (synthesized from IDT), cells were treated at the indicated concentrations for 48 h prior to starting an assay.
EDU incorporation assay
Cells were cultured with 5-ethynyl-2′-deoxyuridine (EdU) treatment (100 nM) for 4 h. The formalin-fixed cells were stained with Tris (100 mM), CuSO4 (1 mM), fluorescent-488 azide (100 μM), ascorbic acid (50 mM), and DAPI (1 μg/ml) for 10 min. Three independent field of views for stained cells were imaged by fluorescence microscopy and counted for statistical analysis.
Antibodies
Antibodies used in this study were Integrin β4 (Thermo Fisher, MA180984), Pan-cytokeratin (Cell Signaling, 4545), Cyclin E (Santa Cruz, sc-481), beta-Actin (Sigma-Aldrich, A5441), FAK (Santa Cruz, sc558), pY397-FAK (Abcam, 39967), SMA (Abcam, 7817), CD63 (System Biosciences, EXOAB-CD63A-1), CD81 (Cell Signaling, 10037), ALIX (Cell Signaling, 2171), and GAPDH (Cell Signaling, 2118).
Histology and immunohistochemistry
For histological analysis of tissues, samples were fixed overnight in 10% phosphate-buffered formalin, dehydrated in alcohol gradients, xylene, and paraffin before being embedded. Then, they were sectioned (5 μm) and subjected to immunohistochemistry as previously described [18].
Immunoblotting
Cell lysates were prepared with modified radio immunoprecipitation assay buffer as described previously [18] with the addition of Halt protease and phosphatase inhibitors (Thermo Scientific, 78425; 78428). Protein concentrations were quantified by the bicinchoninic acid method, subjected to SDS-PAGE and analyzed by immunoblotting as described previously [18].
Exosome isolation and characterization
Exosomes were isolated by ultracentrifugation as described previously [29, 30]. Exosome size was determined using Nanosight and purity by CD63 flow cytometry. Scanning electron microscopy was used to validate the size distribution of exosomes.
Quantitative PCR
Total RNA was isolated from cells using an RNAeasy kit (Qiagen, 74004) per the manufacturer’s instructions. miRNAs were prepared from isolated exosomes using the mirVana miRNA Isolation Kit (Thermo Scientific) according to the manufacturer’s instructions. Equal amounts of RNA were then reverse-transcribed using SuperScript III first-strand synthesis kit (Invitrogen, 18080–044) using random hexamers as primers. cDNA samples were then subjected to qRT-PCR analysis with SYBR Green (BioRad, 1725121) in a BioRad CFXConnect (Bio-Rad, Hercules, CA, USA) thermo-cycler. For miRNA analysis, snRNA RNU6B was used to normalize expression levels. List of primers used are available upon request.
miRNA-sequencing analysis
MiRNA sequencing was performed at the Genomics, Epigenomics and Sequencing Core in University of Cincinnati. Briefly, NEBNext small RNA sample library preparation kit (NEB, Ipswich, MA) was used to prepare the library for sequencing, with the following modification for precise library size selection. First, RNA 3′ adapter is specifically modified to target miRNAs and other small RNAs that have a 3′ hydroxyl group resulting from enzymatic cleavage by Dicer or other RNA processing enzymes. After ligation, the excess 3′ adapter is removed by hybridization to prevent adapter-dimer formation. Then, the 5′ ends of miRNAs that have a 5′-phosphate are ligated to the 5′ adapter. Following 3′ and 5′ ligation, reverse transcription is performed to convert the ligated small RNAs into cDNA, and then uniquely indexed by PCR to generate the sequencing library. The libraries containing DNAs ranging from 135 to 146 bp were purified on 2.75% agarose gels (using a custom-designed DNA ladder for 135–146 bp mixed with library DNA) and eluted in 20 μl. Two microliters of the libraries were then diluted (1:104) and analyzed by NEBNext Library Quant kit (NEB) using QuantStudio 5 Real-Time PCR System (Thermofisher, Waltham, MA). The quantified libraries were clustered onto a flow cell at the concentration of 15 pM using Illumina’s TruSeq SR Cluster Kit v3, and sequenced for 51 cycles using TruSeq SBS kit on Illumina HiSeq system. Sequence reads were aligned and analyzed for differential levels of individual miRNAs by The Laboratory for Statistical Genomics and Systems Biology in the University of Cincinnati.
Statistical analysis
Data were plotted as means ± SEM and statistical significance was determined using a two-tailed t-test.
References
Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16:582–98.
Orimo A, Weinberg RA. Stromal fibroblasts in cancer: a novel tumor-promoting cell type. Cell Cycle. 2006;5:1597–601.
Costa A, Kieffer Y, Scholer-Dahirel A, Pelon F, Bourachot B, Cardon M, et al. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell. 2018;33:463–79 e410.
Ruivo CF, Adem B, Silva M, Melo SA. The biology of cancer exosomes: insights and new perspectives. Cancer Res. 2017;77:6480–8.
Simons M, Raposo G. Exosomes-vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21:575–81.
Luga V, Zhang L, Viloria-Petit AM, Ogunjimi AA, Inanlou MR, Chiu E, et al. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell. 2012;151:1542–56.
Tkach M, Thery C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164:1226–32.
Hannafon BN, Carpenter KJ, Berry WL, Janknecht R, Dooley WC, Ding WQ. Exosome-mediated microRNA signaling from breast cancer cells is altered by the anti-angiogenesis agent docosahexaenoic acid (DHA). Mol Cancer. 2015;14:133.
Donnarumma E, Fiore D, Nappa M, Roscigno G, Adamo A, Iaboni M, et al. Cancer-associated fibroblasts release exosomal microRNAs that dictate an aggressive phenotype in breast cancer. Oncotarget. 2017;8:19592–608.
Parsons JT. Focal adhesion kinase: the first ten years. J Cell Sci. 2003;116:1409–16.
Schaller MD. Biochemical signals and biological responses elicited by the focal adhesion kinase. Biochim Biophys Acta. 2001;1540:1–21.
Schlaepfer DD, Mitra SK. Multiple connections link FAK to cell motility and invasion. Curr Opin Genet Dev. 2004;14:92–101.
Siesser PM, Hanks SK. The signaling and biological implications of FAK overexpression in cancer. Clin Cancer Res. 2006;12:3233–7.
Zhao J, Guan JL. Signal transduction by focal adhesion kinase in cancer. Cancer Metastasis Rev. 2009;28:35–49.
Golubovskaya VM, Cance WG. Focal adhesion kinase and p53 signaling in cancer cells. Int Rev Cytol. 2007;263:103–53.
McLean GW, Carragher NO, Avizienyte E, Evans J, Brunton VG, Frame MC. The role of focal-adhesion kinase in cancer—a new therapeutic opportunity. Nat Rev Cancer. 2005;5:505–15.
Lahlou H, Sanguin-Gendreau V, Zuo D, Cardiff RD, McLean GW, Frame MC, et al. Mammary epithelial-specific disruption of the focal adhesion kinase blocks mammary tumor progression. Proc Natl Acad Sci USA. 2007;104:20302–7.
Luo M, Fan H, Nagy T, Wei H, Wang C, Liu S, et al. Mammary epithelial-specific ablation of the focal adhesion kinase suppresses mammary tumorigenesis by affecting mammary cancer stem/progenitor cells. Cancer Res. 2009;69:466–74.
Provenzano PP, Inman DR, Eliceiri KW, Beggs HE, Keely PJ. Mammary epithelial-specific disruption of focal adhesion kinase retards tumor formation and metastasis in a transgenic mouse model of human breast cancer. Am J Pathol. 2008;173:1551–65.
Pylayeva Y, Gillen KM, Gerald W, Beggs HE, Reichardt LF, Giancotti FG. Ras- and PI3K-dependent breast tumorigenesis in mice and humans requires focal adhesion kinase signaling. J Clin Invest. 2009;119:252–66.
Tavora B, Batista S, Reynolds LE, Jadeja S, Robinson S, Kostourou V, et al. Endothelial FAK is required for tumour angiogenesis. EMBO Mol Med. 2010;2:516–28.
Tavora B, Reynolds LE, Batista S, Demircioglu F, Fernandez I, Lechertier T, et al. Endothelial-cell FAK targeting sensitizes tumours to DNA-damaging therapy. Nature. 2014;514:112–6.
Zhao X, Peng X, Sun S, Park AY, Guan JL. Role of kinase-independent and -dependent functions of FAK in endothelial cell survival and barrier function during embryonic development. J Cell Biol. 2010;189:955–65.
Bou-Gharios G, Garrett LA, Rossert J, Niederreither K, Eberspaecher H, Smith C, et al. A potent far-upstream enhancer in the mouse pro alpha 2(I) collagen gene regulates expression of reporter genes in transgenic mice. J Cell Biol. 1996;134:1333–44.
Hu B, Wu Z, Nakashima T, Phan SH. Mesenchymal-specific deletion of C/EBPbeta suppresses pulmonary fibrosis. Am J Pathol. 2012;180:2257–67.
Zheng B, Zhang Z, Black CM, de Crombrugghe B, Denton CP. Ligand-dependent genetic recombination in fibroblasts: a potentially powerful technique for investigating gene function in fibrosis. Am J Pathol. 2002;160:1609–17.
Costea DE, Hills A, Osman AH, Thurlow J, Kalna G, Huang X, et al. Identification of two distinct carcinoma-associated fibroblast subtypes with differential tumor-promoting abilities in oral squamous cell carcinoma. Cancer Res. 2013;73:3888–901.
Shiga K, Hara M, Nagasaki T, Sato T, Takahashi H, Takeyama H. Cancer-associated fibroblasts: their characteristics and their roles in tumor growth. Cancers. 2015;7:2443–58.
Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol. 2015;1295:179–209.
Hoshino A, Costa-Silva B, Shen TL, Rodrigues G, Hashimoto A, Tesic Mark M, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527:329–35.
Dalerba P, Cho RW, Clarke MF. Cancer stem cells: models and concepts. Annu Rev Med. 2007;58:267–84.
Rosen JM, Jordan CT. The increasing complexity of the cancer stem cell paradigm. Science. 2009;324:1670–3.
Stingl J, Caldas C. Molecular heterogeneity of breast carcinomas and the cancer stem cell hypothesis. Nat Rev Cancer. 2007;7:791–9.
Wicha MS, Liu S, Dontu G. Cancer stem cells: an old idea-a paradigm shift. Cancer Res. 2006;66:1883–90.discussion 1895–96.
Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA. 2002;99:15524–9.
Li Y, Deng X, Zeng X, Peng X. The role of Mir-148a in Cancer. J Cancer. 2016;7:1233–41.
Rivas MA, Venturutti L, Huang YW, Schillaci R, Huang TH, Elizalde PV. Downregulation of the tumor-suppressor miR-16 via progestin-mediated oncogenic signaling contributes to breast cancer development. Breast Cancer Res. 2012;14:R77.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
Duran A, Hernandez ED, Reina-Campos M, Castilla EA, Subramaniam S, Raghunandan S, et al. p62/SQSTM1 by binding to vitamin D receptor inhibits hepatic stellate cell activity, fibrosis, and liver cancer. Cancer Cell. 2016;30:595–609.
Umemura A, He F, Taniguchi K, Nakagawa H, Yamachika S, Font-Burgada J, et al. p62, upregulated during preneoplasia, induces hepatocellular carcinogenesis by maintaining survival of stressed HCC-initiating cells. Cancer Cell. 2016;29:935–48.
Serrels A, Lund T, Serrels B, Byron A, McPherson RC, von Kriegsheim A, et al. Nuclear FAK controls chemokine transcription, tregs, and evasion of anti-tumor immunity. Cell. 2015;163:160–73.
Serrels B, McGivern N, Canel M, Byron A, Johnson SC, McSorley HJ, et al. IL-33 and ST2 mediate FAK-dependent antitumor immune evasion through transcriptional networks. Sci Signal. 2017;10:eaan8355.
Jiang H, Hegde S, Knolhoff BL, Zhu Y, Herndon JM, Meyer MA, et al. Targeting focal adhesion kinase renders pancreatic cancers responsive to checkpoint immunotherapy. Nat Med. 2016;22:851–60.
Symeonides SN, Anderton SM, Serrels A. FAK-inhibition opens the door to checkpoint immunotherapy in pancreatic cancer. J Immunother Cancer. 2017;5:17.
Luo M, Zhao X, Chen S, Liu S, Wicha MS, Guan JL. Distinct FAK activities determine progenitor and mammary stem cell characteristics. Cancer Res. 2013;73:5591–602.
Fan H, Zhao X, Sun S, Luo M, Guan JL. Function of focal adhesion kinase scaffolding to mediate endophilin A2 phosphorylation promotes epithelial-mesenchymal transition and mammary cancer stem cell activities in vivo. J Biol Chem. 2013;288:3322–33.
Shimoda M, Principe S, Jackson HW, Luga V, Fang H, Molyneux SD, et al. Loss of the Timp gene family is sufficient for the acquisition of the CAF-like cell state. Nat Cell Biol. 2014;16:889–901.
Augsten M. Cancer-associated fibroblasts as another polarized cell type of the tumor microenvironment. Front Oncol. 2014;4:62.
Su S, Chen J, Yao H, Liu J, Yu S, Lao L, et al. CD10(+)GPR77(+) cancer-associated fibroblasts promote cancer formation and chemoresistance by sustaining cancer stemness. Cell. 2018;172:841–56e816.
Sun S, Wu HJ, Guan JL. Nuclear FAK and its kinase activity regulate VEGFR2 transcription in angiogenesis of adult mice. Sci Rep. 2018;8:2550.
Acknowledgements
We would like to thank Glenn Doerman for his help in the preparation of figures. We are grateful to members of our laboratory for their critical comments and suggestions for the manuscript. The work in this study is funded by NIH grants R01CA163493, R01NS094144, and R01HL073394 to J-LG.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Wu, HJ., Hao, M., Yeo, S.K. et al. FAK signaling in cancer-associated fibroblasts promotes breast cancer cell migration and metastasis by exosomal miRNAs-mediated intercellular communication. Oncogene 39, 2539–2549 (2020). https://doi.org/10.1038/s41388-020-1162-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41388-020-1162-2
- Springer Nature Limited
This article is cited by
-
Exosomal miRNAs: the tumor's trojan horse in selective metastasis
Molecular Cancer (2024)
-
Targeting focal adhesion kinase boosts immune response in KRAS/LKB1 co-mutated lung adenocarcinoma via remodeling the tumor microenvironment
Experimental Hematology & Oncology (2024)
-
Advances in targeting cancer-associated fibroblasts through single-cell spatial transcriptomic sequencing
Biomarker Research (2024)
-
Mechanism insights and therapeutic intervention of tumor metastasis: latest developments and perspectives
Signal Transduction and Targeted Therapy (2024)
-
The role of cancer-associated fibroblasts and exosomal miRNAs-mediated intercellular communication in the tumor microenvironment and the biology of carcinogenesis: a systematic review
Cell Death Discovery (2024)