
Abstract
Metastasis is the main reason for high mortality in hepatocellular carcinoma (HCC) patients and the molecular mechanism remains unclear. Therefore, it is important to elucidate the mechanism underlying HCC metastasis. Here, we report a novel role of SIX homeobox 4 (SIX4), one of the SIX gene family, in promoting HCC metastasis. The elevated expression of SIX4 was positively correlated with loss of tumor encapsulation, microvascular invasion, higher TNM stage, and poor prognosis in human HCC. SIX4 expression was an independent and significant risk factor for the recurrence and survival in HCC patients. Upregulation of SIX4 promoted HCC invasion and metastasis, whereas downregulation of SIX4 decreased HCC invasion and metastasis. SIX4 transactivated Yes1 associated transcriptional regulator (YAP1) and MET proto-oncogene, receptor tyrosine kinase (MET) expression through directly binding to their promoters. Knockdown of YAP1 and c-MET inhibited SIX4-medicated HCC metastasis, while the stable overexpression of YAP1 and c-MET reversed the decreased metastasis induced by SIX4 knockdown. Hepatocyte growth factor (HGF), the specific ligand of c-MET, upregulated SIX4 expression through ERK/NF-κB pathway. Knockdown of SIX4 significantly decreased HGF-enhanced HCC metastasis. In human HCC tissues, SIX4 expression was positively correlated with nuclear YAP1, c-MET and HGF expression. Patients with positive coexpression of SIX4/ nuclear YAP1, SIX4/c-MET or HGF/SIX4 had the poorest prognosis. Moreover, the combination treatment of YAP1 inhibitor Verteporfin and c-MET inhibitor Capmatinib significantly suppressed SIX4-mediated HCC metastasis. In conclusion, SIX4 is a prognostic biomarker in HCC patients and targeting the HGF-SIX4-c-MET positive feedback loop may provide a promising strategy for the treatment of SIX4-driven HCC metastasis.
Similar content being viewed by others

Introduction
Hepatocellular carcinoma (HCC) is the third most common cause of cancer death worldwide and most patients with HCC have limited treatment options [1, 2]. Although rational approaches have been achieved, the outcomes of HCC patients remain unsatisfactory. HCC is a highly invasive tumor with frequent intrahepatic and distant metastasis, which is the main reason for high recurrence and poor survival of HCC after surgical resection [3]. Thus, it is critical to discover the mechanism underlying HCC metastasis.
The Sine oculis homeobox (SIX) gene family, which belong to a superfamily of homeobox gene family, encode transcription factors containing two evolutionarily conserved domains, homeodomain (HD) and SIX domains. The HD domain is involved in DNA binding, whereas the SIX domain participates in protein-protein interaction [4]. SIX family members are initially identified as the key regulator of the tissue and organ development and construction during embryogenesis [4,5,6]. In humans, SIX family consists of six members, namely, SIX1, SIX2, SIX3, SIX4, SIX5, and SIX6. Accumulating evidence demonstrated that the deregulation of SIX family genes contribute to cancer initiation, progression, and metastasis [4]. Overexpression of SIX1 and SIX2 promotes cancer proliferation, invasion and metastasis, and indicates poor prognosis in several human cancers including colorectal cancer, breast cancer, gastric cancer, and lung cancer [7,8,9,10,11,12]. In contrast, SIX3 expression is dramatically decreased in many human cancers, and functions as a tumor suppressor gene to suppress proliferation, invasion, and metastasis [13, 14]. These studies indicate that deregulation of SIX family genes play critical roles in cancer progression and metastasis.
To investigate the role of SIX genes in HCC, we detected SIX genes expression in 10 normal liver tissues and 30 paired HCC tissues and adjacent nontumor tissues. Among the 6 SIX genes, SIX4 was the most upregulated genes (Supplementary Fig. S1). SIX4 contains 760 amino acids and localizes to the nucleus [4]. SIX4 functions as a transactivator and participates in organ development including myogenesis and neurogenesis [4]. Recent studies reported that elevated expression of SIX4 was associated with poor prognosis in breast cancer, lung cancer, and colorectal cancer, and SIX4 functioned as an oncogene by facilitating cancer cell proliferation and metastasis [15,16,17]. However, the expression and functional role of SIX4 in human HCC remains unknown.
Hepatocyte growth factor (HGF) binds to its receptor tyrosine kinase c-MET and activates mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K) pathways [18]. HGF/c-MET signaling regulates the epithelial-to-mesenchymal and cell migration during embryogenesis and promotes liver regeneration and skin repair [18]. Genetic mouse lacking c-MET or HGF are embryonic lethal [19]. HGF/c-MET signaling promotes liver repair through inducing hepatocyte proliferation, survival, regeneration, and suppress liver fibrosis [19, 20]. Recent studies reported that HGF/c-MET signaling play an important role in promoting HCC growth, angiogenesis, and metastasis [19,20,21,22]. HGF and c-MET transgenic mice are more likely to form hepatocellular adenomas and carcinomas [19]. The elevated expression of c-MET and HGF have been reported in human HCC tissues and indicates poor prognosis [19, 21]. Moreover, c-MET is identified as a critical therapeutic target in human HCC [20, 23]. These studies demonstrate the important role of HGF/c-MET signaling in HCC progression and metastasis. However, the exact mechanism of HGF/c-Met signaling in HCC metastasis is still largely unknown.
To date, no studies have reported the expression and potential function of SIX4 in HCC. In this study, we demonstrated that overexpression of SIX4 promoted HCC metastasis by upregulating Yes1 associated transcriptional regulator (YAP1) and c-MET expression. HGF upregulated SIX4 expression through the extracellular-signal-regulated kinase (ERK)/nuclear factor κB (NF-κB) pathway, which formed an HGF-SIX4-c-MET positive feedback loop. Furthermore, combination treatment of YAP1 inhibitor Verteporfin and c-MET inhibitor Capmatinib significantly suppressed SIX4-mediated HCC metastasis.
Results
SIX4 is significantly upregulated in HCC tissues and indicates poor prognosis in HCC patients
We first screened the expression profile of SIX genes in HCC tissues compared to that of adjacent nontumor tissues by Real-time PCR. The mRNA levels of SIX1, SIX2 SIX4, and SIX5 were increased in HCC tissues than in adjacent nontumor tissues. Among them, SIX4 exhibited the largest fold change. In contrast, the mRNA levels of SIX3 were significantly decreased in HCC tissues. In addition, the mRNA level of SIX6 showed no obvious change in HCC tissues compared with those in adjacent nontumor tissues (Supplementary Fig. S1).
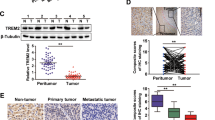
To investigate the potential role of SIX4 in HCC, we analyzed its expression in a cohort of 50 paired HCC tissues. The mRNA levels of SIX4 were dramatically up-regulated in HCC tissues than in adjacent nontumorous tissues and normal liver tissues (Fig. 1a left). The SIX4 mRNA expression was higher in patients with recurrence or metastasis than in patients without recurrence or metastasis (Fig. 1a middle). In addition, the mRNA levels of SIX4 were compared in primary and metastatic HCCs in 30 pairs of HCC specimens. Real-time PCR analysis showed that SIX4 mRNA expression was much higher in metastatic HCC tissues than in primary HCC tissues (Fig. 1a right). A representative case of immunohistochemical staining of SIX4 was shown in Fig. 1b. A higher protein level of SIX4 expression was observed in metastatic HCC samples than in primary HCC samples.

a Relative SIX4 mRNA expression in 10 normal liver tissues and 50 paired HCC and adjacent nontumorous tissues. Relative SIX4 mRNA expression in HCC patients with or without recurrence (n = 30). Relative SIX4 mRNA expression in HCC patients with or without metastasis (n = 30). Relative SIX4 mRNA expression in HCC patients with or without metastasis (n = 30) and in adjacent nontumorous tissues (n = 30). b Representative image of SIX4 expression using IHC staining in adjacent nontumorous tissues, HCC tissues and metastatic HCC tissues. c Representative image of the IHC staining and IHC scores of SIX4 in two HCC cohorts. Statistical analysis was performed by chi-squared test. d Kaplan–Meier analysis of the association of SIX4 expression and recurrence or overall survival in two cohorts of HCC. e Relative mRNA and protein expression of SIX4 in normal liver tissue and HCC cell lines. f Western blot analysis was used to show SIX4 expression in PLC/PRF/5 and MHCC97H cells after lentivirus transfection. g Transwell shown the migratory and invasive abilities of HCC cells after the changs of SIX4 expression. h–l In vivo assays shown that SIX4 knockdown can inhibit HCC metastasis. h The nude mice were injected with the indicated cells in the liver. Bioluminescent images were shown. i Incidence of lung metastasis in the treated nude mice. j The number of metastatic nodules in lung. k Overall survival time of nude mice in different groups was shown. l Representative HE staining images of lung tissues from the different groups were shown. *P < 0.05.
We next analyzed the protein expression and the clinical significance of SIX4 with a tissue array of 220 HCC patient samples (Cohort I, n = 220) using immunohistochemical (IHC) staining. SIX4 protein expression was significantly up-regulated in HCC tissues than in adjacent nontumorous tissues, and SIX4 expression was primarily localized to the nucleus (Fig. 1c). The IHC score of HCC tissues was higher than that of adjacent nontumor tissues (Fig. 1c, and Supplementary Fig. S2). Patients with positive SIX4 expression had shorter overall survival times and higher recurrence rates than patients with negative SIX4 expression (Fig. 1d). The elevated expression of SIX4 was significantly correlated with loss of tumor encapsulation, microvascular invasion, and a higher tumor-nodule-metastasis (TNM) stage (Table 1). Multivariate analysis showed that SIX4 expression was an independent and significant risk factor for recurrence and reduced survival (Supplementary Table S1). The expression and clinical significance of SIX4 was validated in an independent cohort of HCC tissues (Cohort II, n = 190). Similarly, overexpression of SIX4 was significantly correlated with loss of tumor encapsulation, microvascular invasion, higher TNM stage (Table 1), and indicated poor prognosis (Fig. 1d). Multivariate analysis showed that SIX4 over-expression was an independent predictor for postoperative recurrence and overall survival (Supplementary Table S1). Taken together, these studies suggested that SIX4 was a prognostic biomarker in HCC patients.
Overexpression of SIX4 promotes HCC invasion and metastasis
We then detected SIX4 expression in HCC cell lines and found that SIX4 expression was higher in HCC cells with high metastatic capability than in HCC cells with low metastatic capability (Fig. 1e). PLC/PRF/5 and MHCC97H cells were used to establish stable cell lines, PLC/PRF/5-SIX4, MHCC97H-shSIX4-1 and MHCC97H-shSIX4-2, with lentivirus infection (Fig. 1f). Overexpression of SIX4 increased the migrative and invasive ability of PLC/PRF/5 cells, while the knockdown of SIX4 decreased the migratory and invasive ability of MHCC97H cells (Fig. 1g). The in vivo metastatic assay showed that overexpression of SIX4 expedited the lung metastasis rate and increased the number of metastatic lung nodules and lowered the survival time of the nude mice. In contrast, downregulation of SIX4 expression largely impaired the lung metastasis rate and the number of metastatic lung nodules and prolonged the survival time of the nude mice (Fig. 1h–l). These studies suggested that SIX4 promoted HCC invasion and metastasis.
We detected the effect of SIX4 on HCC proliferation and tumor growth. Cell Counting Kit-8 (CCK8) and colony formation assays showed that overexpression of SIX4 increased cell proliferation of PLC/PRF/5 cells, whereas knockdown of SIX4 decreased cell proliferation of MHCC97H cells (Supplementary Fig. S3A, B). In vivo tumorigenicity assays showed that overexpression of SIX4 increased tumor growth of PLC/PRF/5 cells, whereas knockdown of SIX4 decreased tumor growth of MHCC97H cells, which was further confirmed by the IHC staining for Ki67 in the xenograft tumors of different groups (Supplementary Fig. S3C–E). These studies suggested that SIX4 promoted HCC cell proliferation.
YAP1 and MET are two direct transcriptional targets of SIX4
To investigate the molecular mechanism underlying SIX4-mediated HCC metastasis, we compared the mRNA expression profiles between PLC/PRF/5-SIX4 and PLC/PRF/5-control cells using a Liver cancer RT2 Profiler PCR Array. Overexpression of SIX4 induced the expression of several liver cancer-related genes, including MET, YAP1, TLR4, EGFR, HGF, MYC, CTNNB1, CXCR4, and ADAM17 (Supplementary Table S2). Among these genes, both c-MET and YAP1 have been reported to be dramatically upregulated in HCC tissues and indicated poor prognosis [19, 24]. Overexpression of either c-MET or YAP1 promoted HCC invasion and metastasis. Considering the critical role of c-MET and YAP1 in metastasis, we determined whether they were involved in SIX4-mediated HCC metastasis. Overexpression of SIX4 upregulated YAP1 and c-MET expression, whereas knockdown of SIX4 reduced YAP1 and c-MET expression (Fig. 2a). Luciferase reporter assay showed that overexpression of SIX4 transactivated the YAP1 and MET promoter activities (Fig. 2b).

a Relative mRNA expression and protein levels of YAP1 and c-MET were analyzed by RT-qPCR and western blotting in the indicated cells. b Luciferase reporter assay was performed in the indicated cells cotransfected with pCMV-SIX4 and the YAP1 or MET promoter luciferase construct. c, d Serially truncated and mutated YAP1 or MET promoter constructs were cotransfected with pCMV-SIX4 in PLC/PRF/5 cells and relative luciferase activities were measured. e, f ChIP assays revealed the binding enrichment of SIX4 in YAP1 or MET promoters in HCC cell lines and in HCC specimens. g Western blotting analysis of the expression of nuclear YAP1 in the indicated HCC cells. h Western blotting analysis of the expression of YAP1 target genes JAG1, MYC, CYR61 and CTGF in the indicated HCC cells. i Western blotting analysis of the expression of p-c-MET and downstream signaling pathway in the indicated HCC cells.
In order to test how SIX4 regulated YAP1 and MET expression, the promoter sequences of YAP1 and MET were analyzed and two putative SIX4 binding motifs were found in the YAP1 and MET promoters respectively. A series of reporter plasmid constructs containing truncated or mutated YAP1 and MET promoter sequences were designed. We found that the deletion of the region between −940 and −238 bp significantly reduced the YAP1 reporter activity medicated by SIX4 overexpression and the mutation of putative binding site 1 in the YAP1 promoter reduced the activity of the luciferase reporter that was mediated by SIX4 overexpression (Fig. 2c). Similarly, deletion of the region between −1427 and −504 bp decreased the reporter activity of MET and the mutation of putative binding site 1 in the MET promoter reduced the activity of the luciferase reporter induced by SIX4 overexpression (Fig. 2d). Moreover, chromatin immunoprecipitation (ChIP) assays showed that SIX4 directly bound to the YAP1 and MET promoters in PLC/PRF/5-SIX4 cells lines and human HCC samples (Fig. 2e, f). These findings demonstrated that YAP1 and MET were direct transcriptional targets of SIX4.
Nuclear YAP1, which represents the activated status of YAP1, plays an important role in HCC progression [25]. Overexpression of SIX4 increased the expression of nuclear YAP1, whereas knockdown of SIX4 decreased the expression of nuclear YAP1 (Fig. 2g). Furthermore, overexpression of SIX4 increased the expression of YAP1 target genes, such as CTGF, CYR61, JAG1 and c-MYC, whereas knockdown of SIX4 decreased YAP1 target genes’ expression (Fig. 2h).
c-MET is predominantly regulated by its phosphorylation [20]. Several studies reported that overexpression of c‐MET causes receptor dimerization and activation and c-MET can be auto-phosphorylated in MHCC97H cells [20, 26, 27]. To determine whether SIX4-mediated c-MET overexpression regulates c-MET phosphorylation and the activation of downstream signaling pathway, we detected the phosphorylation of c-MET and the downstream signaling pathway of c-MET. Western blotting analysis showed that overexpression of SIX4 increased the phosphorylation of c-MET and activated downstream signaling pathway, such as ERK1/2, JNK, p38, AKT, and NF-κB pathway (Fig. 2i).
SIX4 promotes HCC invasion and metastasis through upregulating YAP1 and c-MET
To explore whether YAP1 and c-MET were involved in SIX4-medicated HCC metastasis, we knocked down the expression of YAP1 and c-MET in PLC/PRF/5-SIX4 cells and ectopically overexpressed YAP1 and c-MET in MHCC97H-shSIX4 cells with lentivirus transfection (Fig. 3a). Transwell assay showed that knockdown of YAP1 and c-MET significantly decreased SIX4-enhanced migratory and invasive abilities, whereas overexpression of YAP1 and c-MET rescued the reduced migratory and invasive abilities induced by SIX4 knockdown (Fig. 3b). The in vivo metastatic assay showed that knockdown of YAP1 and c-MET lowered the incidence of lung metastasis and the number of metastatic lung nodules and prolonged the overall survival of the PLC/PRF/5-SIX4 group (Fig. 3c–g). In contrast, overexpression of YAP1 and c-MET reversed decreased lung metastasis and the number of metastatic lung nodules in MHCC97H-shSIX4 group and decreased the overall survival of this group (Fig. 3c–g). These results indicated that SIX4 promoted HCC metastasis by upregulating YAP1 and c-MET expression.

a Western blotting was used to show the SIX4, YAP1 and c-MET expression in HCC cells transfected with lentivirus. b Transwell shown the migratory and invasive abilities in PLC/PRF/5-SIX4 cells with YAP1 or c-MET knockdown and in MHCC97H-shSIX4 cell with YAP or c-MET overexpression. c–g In vivo assays shown that SIX4 promotes HCC metastasis through upregulating YAP1 and c-MET. c The nude mice were implanted with the indicated cells in the liver. Representative bioluminescent images in the different groups were shown. d Incidence of lung metastasis in the treated nude mice. e The number of lung metastatic nodules in the lung was counted. f Overall survival time of the treated nude mice in different groups was shown. g Representative HE staining images of lung tissues from different groups were shown. *P < 0.05.
HGF upregulates SIX4 expression through the c-MET/ERK/NF-kB signaling pathway
Since c-MET was involved in SIX4-mediated HCC metastasis, its specific ligand HGF attracted our attention. HGF-c-MET signaling plays a critical role in promoting HCC metastasis [28]. HGF was identified as a driver oncogene and elevated expression of HGF was positively correlated with poor prognosis in human HCC [21]. Considering the important roles of both HGF and SIX4 in HCC metastasis, we determined whether HGF regulates SIX4 expression. To test this hypothesis, PLC/PRF/5 cells with low endogenous SIX4 expression were treated with HGF. HGF treatment increased SIX4 expression in a dose-dependent manner (Fig. 4a). Of note, HGF treatment transactivated SIX4 promoter activity (Fig. 4b).

a Relative mRNA and protein level of SIX4 were analyzed in PLC/PRF/5 cells after treated with HGF. b SIX4 promoter luciferase activity was measured after HGF treatment in PLC/PRF/5 cells. c Serially truncated and mutated SIX4 promoter constructs were transfected into PLC/PRF/5 cells and treatment with HGF and relative luciferase activity were detected. d, e PLC/PRF/5 cells were transfected with p65 siRNA or control siRNA, control or NF-kB inhibitor BAY 11-7082 and then treatment with HGF. SIX4 promoter activity and expression were measured by luciferase activity assay, RT-qPCR and Western blotting. f PLC/PRF/5 cells were treated with inhibitor of ERK, JNK, P38 and PI3K and then stimulated with HGF. Western blotting was used to detect the expression of SIX4 as well as the total and phosphorylated levels of ERK, JNK, P38, AKT and P65. g A ChIP assay shown the relative enrichment of P65 on SIX4 promoter when the PLC/PRE/5 cells were treated with HGF and inhibitor of ERK, JNK, P38 or PI3K. h Representative IHC staining images of SIX4 and HGF in HCC samples. i The correlation analysis of the SIX4 and HGF expression in two independent cohorts of HCC patients. j, k Overall survival time and recurrence in HCC patients with positive or negative expression of HGF in cohort I (j) and cohort II (k) were shown. l, m Kaplan–Meier analyzed recurrence and overall survival times of patients with coexpression of HGF/SIX4 or HGF/SIX4 in cohort I (l) and cohort II (m). *P < 0.05.
To identify the cis-regulatory elements, which involved in HGF-induced SIX4 expression, the −1845 to +135 bp region and a series of truncations and mutations of the human SIX4 promoter were generated. A significant reduction of SIX4 promoter activity was observed when PLC/PRF/5 cells were transfected with the truncated (−365 to −43) SIX4 promoter construct, suggesting that this region was crucial for HGF-induced SIX4 promoter transactivation. Two potential NF-kB binding sites located in this region. Site-directed mutagenesis showed that mutation of second binding site significantly reduced the SIX4 promoter activity induced by HGF (Fig. 4c). Knockdown of p65 which is a key subunit of NF-kB significantly impaired HGF-induced SIX4 overexpression and SIX4 promoter transactivation (Fig. 4d). Similarly, an NF-kB inhibitor treatment (BAY11-7082) significantly inhibited HGF-mediated SIX4 promoter transactivation and SIX4 upregulation (Fig. 4e).
HGF has been reported to activate MAPK and PI3K-AKT pathways [18]. In order to test which pathway was involved in HGF-mediated SIX4 overexpression, PLC/PRF/5 cells were treated with ERK, c-Jun-N-terminal kinase (JNK), p38 kinases or PI3K inhibitors. Pretreatment of cells with the ERK inhibitor significantly inhibited HGF-mediated SIX4 overexpression, whereas pretreatment of cells with the JNK, P38 or PI3K inhibitors had no significant effect (Fig. 4f). Furthermore, a ChIP assay demonstrated that the ERK inhibitor treatment inhibited the binding of NF-kB to the SIX4 promoter, while the JNK, P38 or PI3K inhibitor treatment showed no effect (Fig. 4g). These results suggested that HGF upregulated SIX4 expression through the ERK/NF-kB signaling pathway.
To confirm the clinical importance of HGF in HCC, IHC analysis was performed in two independent HCC cohorts. HGF expression was upregulated in HCC tissues than that in adjacent nontumor tissues (Fig. 4h). In both cohorts, overexpression of HGF was positively correlated with microvascular invasion, poorer tumor differentiation and higher TNM stage (Supplementary Table S3). SIX4 expression was positively correlated with HGF expression (Fig. 4i). Patients with positive expression of HGF had a higher recurrence rate and poorer overall survival time than patients with negative expression of HGF (Fig. 4j, k). Furthermore, Kaplan–Meier analysis exhibited that patients with positive coexpression of HGF/SIX4 suffered from the highest recurrence risk and shortest survival times in both cohorts (Fig. 4l, m). Moreover, we detected HGF expression in the metastatic lung nodules from PLC/PRF/5-SIX4 xenograft group, indicating that HGF expression was upregulated in the metastatic milieu in the tumor models when SIX4 was expressed (Supplementary Fig. S4).
SIX4 is essential for HGF-induced HCC metastasis
As SIX4 was upregulated by HGF and promoted HCC metastasis, we determined whether SIX4 was involved in HGF-mediated HCC metastasis. PLC/PRF/5 cells were infected with Lentivirus LV-shSIX4 and then were treated with HGF (Fig. 5a). HGF treatment significantly increased the migratory and invasive abilities of PLC/PRF/5 cells, whereas knockdown of SIX4 largely lowered the increased migratory and invasive abilities induced by HGF (Fig. 5b). We then established a stable cell line PLC/PRF/5-HGF through lentiviral transduction and knocked down the expression of SIX4 in PLC/PRF/5-HGF cells (Fig. 5c). Stable overexpression of HGF significantly increased the migratory and invasive abilities of PLC/PRF/5 cells while SIX4 knockdown decreased the enhanced migratory and invasive abilities of PLC/PRF/5-HGF cells (Fig. 5d). The in vivo metastasis experiment showed that overexpression of HGF increased the incidence of lung metastasis and the number of metastatic lung nodules and decreased the overall survival in PLC/PRF/5-HGF group compared with that in control group (PLC/PRF/5-control). However, knockdown of SIX4 decreased the incidence of lung metastasis and the number of metastatic lung nodules while extended the overall survival in the PLC/PRF/5-HGF group (Fig. 5e–i). These results demonstrated that SIX4 was essential for HGF-induced HCC metastasis.

a Western blotting was used to detect the SIX4 expression in PLC/PRF/5 cells treated with HGF or combined HGF treatment and SIX4 knockdown. b Transwell shown the migratory and invasive abilities of PLC/PRF/5 cells treated with HGF or combined HGF treatment and SIX4 knockdown. c Western blotting analyzed the HGF and SIX4 expression in PLC/PRF/5 cells transfected with LV-shcontrol or LV-HGF lentivirus and in PLC/PRF/5-HGF cells transfected with LV-shcontrol or LV-shSIX4 lentivirus. d Transwell shown the migratory and invasive abilities of PLC/PRF/5-HGF cells transfected with LV-shcontrol or LV-shSIX4 lentivirus. e–i In vivo assays shown that SIX4 knockdown can inhibit HGF-induced HCC metastasis. e The nude mice were injected with the indicated cells in the liver. Representative Bioluminescence images were shown in different groups. f Incidence of lung metastasis in the nude mice. g The number of metastatic lung nodules in lung. h Overall survival time of nude mice in different groups. i Representative H&E staining images of lung tissues from nude mice in different groups. *P < 0.05.
SIX4 expression is positively correlated with nuclear YAP1 and c-MET expression in human HCC tissues
We further evaluated the possible association between SIX4 and nuclear YAP1 or c-MET in two independent cohorts of HCC patients. Representative images of the IHC staining were shown in Fig. 6a. IHC staining showed that YAP1 was mainly localized in the nucleus (Fig. 6a). SIX4 expression was positively correlated with nuclear YAP1 and c-MET expression in both cohorts (Fig. 6b, c). The elevated expression of both nuclear YAP1 and c-MET were positively correlated with microvascular invasion and higher TNM stage (Supplementary Tables S4, 5). Patients with positive expression of nuclear YAP1 or c-MET exhibited a higher recurrence rate and shorter overall survival compared with patients with negative expression of nuclear YAP1 or c-MET (Fig. 6d, e). Furthermore, Kaplan–Meier analysis exhibited that HCC patients with positive coexpression of either SIX4/ nuclear YAP1 or SIX4/c-MET had the highest recurrence risk and shortest survival time in both HCC cohorts (Fig. 6f, g).

a Representative IHC staining images shown SIX4, nuclear YAP1 and c-MET expression in human HCC tissues. b, c The correlation analysis of the expression of SIX4 and nuclear YAP1 or SIX4 and c-MET in human HCC tissues from cohort I (b) and cohort II (c). d, e Overall survival time and recurrence in HCC patients with positive or negative expression of nuclear YAP1 or c-MET in cohort I (d) and cohort II (e) were shown. f, g Kaplan–Meier analyzed recurrence and overall survival times of patients with coexpression of SIX4/ nuclear YAP1 or SIX4/c-MET in cohort I (f) and cohort II (g).
Moreover, the mRNA and protein levels of YAP1 and c-MET were compared in primary and metastatic HCCs in 30 pairs of HCC specimens. Real-time PCR analysis showed that the mRNA expressions of both YAP1 and MET were much higher in metastatic HCC tissues than in primary HCC tissues (Supplementary Fig. S5A). Representative cases of immunohistochemical staining of YAP1 and c-MET were shown in Supplementary Fig. S5B, C. The protein levels of both YAP1 and MET were much higher in metastatic HCC tissues than in primary HCC tissues (Supplementary Fig. S5B, C).
Combined treatment of YAP1 and c-MET inhibitor significantly suppress SIX4-medicated HCC metastasis
Verteporfin is a YAP1 inhibitor, which disrupts YAP-TEAD interactions [29]. Capmatinib is a highly selective and potent c-Met inhibitor which inhibit c-MET activation [30]. Our above research has demonstrated that HGF/c-MET signaling induced SIX4 overexpression, and SIX4 promoted HCC metastasis through upregulating YAP1 and c-MET expression. Therefore, we determined whether combination treatment of YAP1 and c-MET inhibitors had any effect on SIX4-mediated HCC metastasis. PLC/PRF/5-SIX4 cells were treated with YAP1 inhibitor Verteporfin or c-MET inhibitor Capmatinib alone, or the combination of both inhibitors, and the protein levels of YAP1, c-MET and phospho-MET were shown (Fig. 7a). Treatment with Verteporfin or Capmatinib alone partially decreased the migratory and invasive abilities of PLC/PRF/5-SIX4 cells, whereas combination of the two agents dramatically lowered the migratory and invasive abilities of PLC/PRF/5-SIX4 cells (Fig. 7b). In order to further investigate this effect, we designed the in vivo experiment (Fig. 7c). The in vivo metastasis assay showed that Verteporfin or Capmatinib treatment alone partially decreased the incidence of lung metastasis and the number of metastatic lung nodules while partially increasing the overall survival time of the PLC/PRF/5-SIX4 group, whereas the combination of Verteporfin and Capmatinib dramatically inhibited the number of lung metastasis and largely prolonged survival time compared with control or single agent treatment (Fig. 7d–h). These studies suggested that the combination treatement of YAP1 inhibitor Verteporfin and c-MET inhibitor Capmatinib significantly suppressed SIX4-mediated HCC metastasis.

a PLC/PRF/5-SIX4 cells treated with YAP1 inhibitor VP or c-MET inhibitor Capmatinib alone, or the combination of both inhibitors. Western blotting shown the expression of SIX4, YAP, and c-MET. b Transwell shown the migratory and invasive abilities of PLC/PRF/5-SIX4 cells in different treatment groups. c The diagram of vivo treatment in nude mice. One week after injection of PLC-PRF/5-SIX4 cells, these mice were randomly divided into four group and treated with vehicle, Verteporfin or Capmatinib or combined treatment. d–h In vivo assays shown that combined treatment of YAP1 and c-MET inhibitor can largely block HCC metastasis. d Representative Bioluminescence images were shown in different groups. e Incidence of lung metastasis in the nude mice. f The number of metastatic lung nodules in lung. g Overall survival time of nude mice in different groups. h Representative H&E staining images of lung tissues from nude mice in different groups. i A schematic diagram of the role of HGF-SIX4 signaling in HCC metastasis. HGF-c-MET signaling upregulates SIX4 expression through ERK/NF-kB pathway. SIX4 promotes HCC metastasis through transactivating YAP1 and c-MET. Combined YAP1 inhibitor Verteporfin and c-MET inhibitor Capmatinib almost abolished SIX4-induced HCC metastasis. *P < 0.05.
Discussion
Metastasis is still the main reason for the high mortality of HCC patients [3]. Thus, exploring the metastatic mechanism and the potential therapeutic target is urgent needed. In this study, we found that SIX4 was upregulated in HCC tissues compared with adjacent nontumorous tissues. Overexpression of SIX4 was significantly correlated with loss of tumor encapsulation, microvascular invasion, and a higher TNM stage. HCC patients with positive SIX4 expression had shorter overall survival times and higher recurrence rates than patients with negative SIX4 expression. A multivariate analysis revealed that SIX4 expression was an independent risk factor for higher recurrence and shorter overall survival in HCC patients. Furthermore, SIX4 expression was higher in HCC tissues from patients who developed metastasis than in HCC tissues from patients who did not develop metastasis. In addition, we found that upregulation of SIX4 promoted HCC invasion and metastasis and knockdown of SIX4 expression decreased HCC invasion and metastasis. These studies suggested that SIX4 was a prognostic biomarker of HCC. The first step of cancer metastasis is growth of neoplastic cells [31]. Primary tumor grows need to develop a blood supply for metabolic needs, which is called angiogenesis. This process can also provide an escape route by which cells can leave the tumor and enter into the body’s circulatory blood system. Once the metastatic cells in the new site, cells must initiate and maintain growth for a macroscopic tumor to form [32, 33]. In this work, we found that upregulation of SIX4 promoted HCC proliferation and knockdown of SIX4 expression decreased HCC proliferation. Therefore, SIX4-mediated HCC proliferation may be a reason for SIX4-mediated HCC metastasis.
YAP1 is essential for cancer initiation, progression, and metastasis in several solid tumors through interacting with TAZ and other transcription factors [34]. YAP1 is an independent prognostic marker in HCC and promote liver tumorigenesis through the cooperation between MYC and β-catenin [35, 36]. Liver-specific YAP1 overexpression leads to liver tumor in transgenic mice [37]. YAP1 overexpression promotes HCC progression and metastasis through enhancing the expression of genes involved in proliferation and stemness and regulating metabolism and cytoskeleton [24, 38, 39]. c-MET is a well-known oncogene in human cancers and is required to maintain the transformed and metastatic phenotype [40, 41]. c-MET aberrations occur in nearly half of HCC patients through several ways and is identified as an oncogenic driver in HCC [19, 20]. Therefore, c-MET is a promising target in HCC [23]. These evidences indicate the crucial roles of both YAP1 and c-MET in promoting HCC progression. In this study, we illustrated that SIX4 upregulated YAP1 and c-MET expression through directly binding to their promoters. Knockdown of YAP1 and c-MET decreased SIX4-enhanced HCC metastasis, whereas ectopic overexpression of YAP1 and c-MET rescued the decreased HCC metastasis induced by SIX4 knockdown. In human HCC samples, SIX4 expression was positively correlated with YAP1 and c-MET expression, and patients with positive coexpression of SIX4/ nuclear YAP1 or SIX4/c-MET exhibited the worst prognosis. Taken together, both clinical evidence and experimental data demonstrated that SIX4 promoted HCC metastasis through upregulating YAP1 and c-MET expression.
The regulatory mechanism of SIX4 overexpression in human HCC remains unclear. SIX1 is upregulated in Ink4a/Arf-deficient mice transgenic for hepatocyte growth factor/scatter factor (HGF/SF) and has been proved to be an important factor in promoting metastasis of rhabdomyosarcoma (RMS) [42]. This study suggested that HGF may regulate other SIX family members. Therefore, HGF, the specific ligand of c-MET, attracted our attention. Physiologically, HGF-c-MET signaling participates in embryogenesis, wound healing, organ regeneration, angiogenesis and morphogenic differentiation [18, 43]. Moreover, HGF-c-MET signaling plays a critical role in promoting HCC angiogenesis, invasion, and metastasis [40, 41, 44, 45]. The elevated expression of both HGF and c-MET are associated with poor prognosis in human HCC patients [21, 46, 47]. In this study, we found that HGF-c-MET signaling upregulated SIX4 expression through the ERK/NF-kB pathway. SIX4 upregulated the receptor c-MET expression, which formed an HGF-SIX4-c-MET positive feedback loop. Furthermore, knockdown of SIX4 dramatically decreased HGF-mediated HCC metastasis. Thus, we defined an HGF-SIX4-c-MET positive feedback loop that played a pivotal role in HCC metastasis, and targeting this pathway may provide new therapeutic strategy to inhibit HCC metastasis.
To design pharmacological strategy against this positive feedback loop, we focused on both YAP1 and c-MET inhibitors. c-MET inhibitor has been proved to inhibit tumor growth in mice model [48]. However, clinical trials of c-MET inhibitor treatment alone have failed to produce satisfactory outcomes in HCC [20, 23, 30]. Combined treatment of c-MET and EGFR inhibitors in therapy of colorectal cancer, non-small cell lung cancer and HCC have been processed [49,50,51]. Verteporfin is a YAP1 inhibitor through disrupting the formation of the YAP1-TEAD complex [29]. Verteporfin inhibits liver cancer initiation and enhances the effect of sorafenib in HCC [52, 53]. Based on these studies, we hypothesized that whether the combination of YAP1 inhibitor Verteporfin and c-MET inhibitor Capmatinib had any effect on HCC metastasis. Our in vivo data showed that the combination of both inhibitors dramatically suppressed SIX4-mediated HCC metastasis compared with control or single agent alone. These results provided a new therapeutic strategy to inhibit SIX4-driven HCC metastasis.
In conclusion, we demonstrated that upregulated SIX4 induced by HGF promoted HCC metastasis through transactivating YAP1 and c-MET expression. The combination treatment of YAP1 and c-MET inhibitors significantly suppressed SIX4-mediated HCC metastasis. Thus, SIX4 is a prognostic biomarker in HCC patients and targeting the HGF- SIX4-c-MET positive feedback loop may provide a promising strategy for the treatment of SIX4-driven HCC metastasis.
Materials and methods
Cell culture
Immortalized liver cell lines (HL-7702) and HCC cells (Huh6 and JHH-7) were purchased from the Institute of Biochemistry and Cell Biology, Chinese Academy of Science, China. Human HCC cells (HepG2, Huh-7, Hep3B, PLC/PRF/5, SNU423, SNU398, and SNU449) were purchased from the American Type Culture Collection. Additional human HCC cells (MHCC97H, HCCLM3, and HCCLM6) were kindly provided by Dr Tang ZY (Liver Cancer Institute, Zhongshan Hospital, Fudan University, Shanghai, China). MHCC97H, HCCLM3, and HCCLM6 cells are stepwise metastatic potential cell lines with the same genetic background but different lung metastatic potentials. HepG2, Hep3B, Huh7, and PLC/PRF/5 are HCC cells with low metastatic potential, whereas MHCC97H, HCCLM3, and HCCLM6 are HCC cells with high metastatic potential. Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) at 37 °C in a 5% CO2 incubator. The medium was supplemented with 10% FBS, 100 μg/ml penicillin, and 100 μg/ml streptomycin. These above cell lines were authenticated by short tandem repeats (STRs) DNA profiling. All cells were tested for mycoplasma contamination before use with the Universal Mycoplasma Detection Kit (ATCC 30–1012 K) and were not contaminated by mycoplasma.
Patients and follow-up
This study was approved by the Ethics Committee of Tongji Medical College. All patients provided full consent for the study. Cohort I included 220 adult patients with HCC who underwent curative resection between 2003 and 2005 at the Tongji Hospital of Tongji Medical College (Wuhan, China). Cohort II included 190 adult patients with HCC who underwent curative resection between 2006 and 2008 at the Tongji Hospital of Tongji Medical College (Wuhan, China). A preoperative clinical diagnosis of HCC was based on the diagnostic criteria of the American Association for the Study of Liver Diseases. The inclusion criteria were as follows: (a) distinctive pathologic diagnosis; (b) no preoperative anticancer treatment or distant metastases; (c) curative liver resection; and (d) complete clinicopathologic and follow-up data. The differentiation statuses were graded according to the method of Edmondson and Steine. The pTNM classification for HCC was based on The American Joint Committee on Cancer/International Union Against Cancer staging system (6th edition, 2002). Follow-up data were summarized at the end of December 2013 (Cohort I) and December 2016 (Cohort II, range 4–96 months) respectively. The patients were evaluated every 2–3 months during the first 2 years and every 3–6 months thereafter. All follow-up examinations were performed by physicians who were blinded to the study. During each check-up, the patients were monitored for tumor recurrence by measuring the serum AFP levels and by performing abdominal ultrasound examinations. A computed tomography and/or magnetic resonance imaging examination was performed every 3–6 months, together with a chest radiographic examination. The diagnostic criteria for HCC recurrence were the same as the preoperative criteria. The time to recurrence and overall survival were the primary endpoints. The time to recurrence was calculated from the date of resection to the date of a diagnosis with tumor recurrence. The overall survival was calculated from the date of resection to the date of death or of the last follow-up.
In addition, 10 normal liver tissues, 50 pairs of fresh HCC tissues and adjacent nontumor tissue samples and 30 pairs of fresh metastatic and matched primary HCC tissues and adjacent nontumor tissue samples were collected after surgical resection and were used to investigate the mRNA expression levels of SIX4.
Construction of tissue microarrays and immunohistochemistry
HCC samples and the corresponding adjacent liver tissues were used to construct a tissue microarray (Shanghai Biochip Co., Ltd. Shanghai, China). IHC was performed on 4 μm-thick, routinely processed paraffin-embedded sections. Briefly, the tissue sections were deparaffinized after baking at 60 °C for an hour. Endogenous peroxidase activity was blocked by 3% (vol/vol) hydrogen peroxide in methanol for 12 min and washes with phosphate-buffered saline (PBS). Then the slides were immersed in 0.01 mol/L citrate buffer solution (pH 6.0) and placed in a microwave oven for 30 min. After washed with PBS, the sections were incubated with the primary antibody diluted in PBS containing 1% (wt/vol) bovine serum albumin in 4 °C for overnight. The tissue microarray was stained for SIX4 (MilliporeSigma, SAB1405112), YAP1 (Cell signaling technology, #14074), c-MET (Cell signaling technology, #8198), HGF (Abcam, ab83760), HGF (Cell signaling technology, # 52445) and KI67 (Abcam, ab15580) expression. Negative controls were performed by replacing the primary antibody with preimmune mouse serum. After washed with PBS, the sections were treated with a peroxidase-conjugated second antibody (Santa Cruz) for 30 min at room temperature and then washed with PBS. Reaction product was visualized with diaminobenzidine for 2 min. Images were obtained under a light microscope (Olympus, Japan) equipped with a DP70 digital camera.
Analyses were performed by two independent observers who were blinded to the clinical outcome. The immunostaining intensity was scored on a scale of 0–3: 0 (negative), 1 (weak), 2 (medium) or 3 (strong). The percentage of positive cells was evaluated on a scale of 0–4: 0 (negative), 1 (1–25%), 2 (26–50%), 3 (51–75%), or 4 (76–100%). The final immuno-activity scores were calculated by multiplying the above two scores, resulting an overall score which range from 0 to 12. Each case was ultimately considered “negative” if the final score ranges from 0 to 3, and “positive” if the final score ranges from 4 to 12 as described previously [54].
In vivo metastatic model and bioluminescent imaging
All animal experiments were approved by the Committee on the Tongji Hospital of Tongji Medical College, Huazhong University of Science and Technology. BALB/C nude mice (male, 5 weeks old) were housed and cared according to the institutional guidelines for animal care. Mice were randomly assigned into experimental or control groups, blinding was not possible. For in vivo metastasis assay, 2 × 106 cells were resuspended in 50 μL PBS/matrigel mixture. Under anesthesia, mice were orthotopically inoculated in the left hepatic lobe with the indicated cells through an 8 mm transverse incision in the upper abdomen (n = 10 mice/group). For drug-based drug intervention, mice were given daily oral doses of Capmatinib 10 mg/kg [30] and/or intraperitoneal injection of Verteporfin (VP) 50 mg/kg [52] twice every week. The in vivo tumor formation and metastases were monitored using the bioluminescence. For in vivo signal detection, D-luciferin (Perkin-Elmer) at 100 mg/kg was injected intraperitonially into the nude mice. Bioluminescent images were captured using a Lago X optical imaging system Imaging System (SI Imaging). At the 9 weeks, the mice were sacrificed and the lungs were collected for histological examination.
Statistical analysis
All values were recorded as the mean ± standard deviation (sd). P values were statistically analyzed by the χ2 test for categorical variables and by Student’s test for quantitative data. Survival was calculated with the Kaplan–Meier method (log-rank test). Multivariate analysis was performed by Cox regression analysis. P < 0.05 was considered statistically significant. Statistical values were calculated with SPSS software (Version 20.0).
All other materials and methods can be found in the supplementary materials.
References
Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391:1301–14.
Craig AJ, von Felden J, Garcia-Lezana T, Sarcognato S, Villanueva A, et al. Tumour evolution in hepatocellular carcinoma. Nat Rev Gastroenterol Hepatol. 2020;17:139–52.
Yang JD, Hainaut P, Gores GJ, Amadou A, Plymoth A, Roberts LR. A global view of hepatocellular carcinoma: trends, risk, prevention and management. Nat Rev Gastroenterol Hepatol. 2019;16:589–604.
Christensen KL, Patrick AN, McCoy EL, Ford HL. The six family of homeobox genes in development and cancer. Adv Cancer Res. 2008;101:93–126.
Shah AM, Krohn P, Baxi AB, Tavares ALP, Sullivan CH, Chillakuru YR, et al. Six1 proteins with human branchio-oto-renal mutations differentially affect cranial gene expression and otic development. Dis Model Mech. 2020;13:dmm043489.
Liu Z, Li C, Xu J, Lan Y, Liu H, Li X, et al. Crucial and Overlapping Roles of Six1 and Six2 in Craniofacial Development. J Dent Res. 2019;98:572–9.
Li L, Liang Y, Kang L, Liu Y, Gao S, Chen S, et al. Transcriptional Regulation of the Warburg Effect in Cancer by SIX1. Cancer Cell. 2018;33:368–85. e367.
Coletta RD, Christensen K, Reichenberger KJ, Lamb J, Micomonaco D, Huang L, et al. The Six1 homeoprotein stimulates tumorigenesis by reactivation of cyclin A1. Proc Natl Acad Sci USA. 2004;101:6478–83.
McCoy EL, Iwanaga R, Jedlicka P, Abbey NS, Chodosh LA, Heichman KA, et al. Six1 expands the mouse mammary epithelial stem/progenitor cell pool and induces mammary tumors that undergo epithelial-mesenchymal transition. J Clin Investig. 2009;119:2663–77.
Wang CA, Jedlicka P, Patrick AN, Micalizzi DS, Lemmer KC, Deitsch E, et al. SIX1 induces lymphangiogenesis and metastasis via upregulation of VEGF-C in mouse models of breast cancer. J Clin Investig. 2012;122:1895–906.
Ono H, Imoto I, Kozaki K, Tsuda H, Matsui T, Kurasawa Y, et al. SIX1 promotes epithelial-mesenchymal transition in colorectal cancer through ZEB1 activation. Oncogene. 2012;31:4923–34.
Oliphant MUJ, Vincent MY, Galbraith MD, Pandey A, Zaberezhnyy V, Rudra P, et al. SIX2 Mediates Late-Stage Metastasis via Direct Regulation of SOX2 and Induction of a Cancer Stem Cell Program. Cancer Res. 2019;79:720–34.
Yu Z, Feng J, Wang W, Deng Z, Zhang Y, Xiao L, et al. The EGFR-ZNF263 signaling axis silences SIX3 in glioblastoma epigenetically. Oncogene. 2020;39:3163–78.
Zheng Y, Zeng Y, Qiu R, Liu R, Huang W, Hou Y, et al. The Homeotic Protein SIX3 Suppresses Carcinogenesis and Metastasis through Recruiting the LSD1/NuRD(MTA3) Complex. Theranostics. 2018;8:972–89.
Li G, Hu F, Luo X, Hu J, Feng Y. SIX4 promotes metastasis via activation of the PI3K-AKT pathway in colorectal cancer. PeerJ. 2017;5:e3394.
Tang X, Yang Y, Song X, Liu X, Wang X, Huang F, et al. SIX4 acts as a master regulator of oncogenes that promotes tumorigenesis in non-small-cell lung cancer cells. Biochem Biophys Res Commun. 2019;516:851–7.
Sun X, Ma J, Chen Q, Hou Z, Luo X, Wang G, et al. SIX4 promotes metastasis through STAT3 activation in breast cancer. Am J Cancer Res. 2020;10:224–36.
Gherardi E, Birchmeier W, Birchmeier C, Vande Woude G. Targeting MET in cancer: rationale and progress. Nat Rev Cancer. 2012;12:89–103.
Giordano S, Columbano A. Met as a therapeutic target in HCC: facts and hopes. J Hepatol. 2014;60:442–52.
Bouattour M, Raymond E, Qin S, Cheng AL, Stammberger U, Locatelli G, et al. Recent developments of c-Met as a therapeutic target in hepatocellular carcinoma. Hepatology. 2018;67:1132–49.
Yang X, Zhang XF, Lu X, Jia HL, Liang L, Dong QZ, et al. MicroRNA-26a suppresses angiogenesis in human hepatocellular carcinoma by targeting hepatocyte growth factor-cMet pathway. Hepatology. 2014;59:1874–85.
Hu CT, Wu JR, Cheng CC, Wu WS. The Therapeutic Targeting of HGF/c-Met Signaling in Hepatocellular Carcinoma: Alternative Approaches. Cancers (Basel). 2017;9:58.
Faivre S, Rimassa L, Finn RS. Molecular therapies for HCC: Looking outside the box. J Hepatol. 2020;72:342–52.
Zhang S, Zhou D. Role of the transcriptional coactivators YAP/TAZ in liver cancer. Curr Opin Cell Biol. 2019;61:64–71.
Patel SH, Camargo FD, Yimlamai D. Hippo Signaling in the Liver Regulates Organ Size, Cell Fate, and Carcinogenesis. Gastroenterology. 2017;152:533–45.
Liu X, Wang Q, Yang G, Marando C, Koblish HK, Hall LM, et al. A novel kinase inhibitor, INCB28060, blocks c-MET-dependent signaling, neoplastic activities, and cross-talk with EGFR and HER-3. Clin Cancer Res. 2011;17:7127–38.
Lau EY, Lo J, Cheng BY, Ma MK, Lee JM, Ng JK, et al. Cancer-Associated Fibroblasts Regulate Tumor-Initiating Cell Plasticity in Hepatocellular Carcinoma through c-Met/FRA1/HEY1 Signaling. Cell Rep. 2016;15:1175–89.
Wang H, Rao B, Lou J, Li J, Liu Z, Li A, et al. The Function of the HGF/c-Met Axis in Hepatocellular Carcinoma. Front Cell Dev Biol. 2020;8:55.
Gibault F, Bailly F, Corvaisier M, Coevoet M, Huet G, Melnyk P, et al. Molecular Features of the YAP Inhibitor Verteporfin: Synthesis of Hexasubstituted Dipyrrins as Potential Inhibitors of YAP/TAZ, the Downstream Effectors of the Hippo Pathway. ChemMedChem. 2017;12:954–61.
Li H, Li CW, Li X, Ding Q, Guo L, Liu S, et al. MET Inhibitors Promote Liver Tumor Evasion of the Immune Response by Stabilizing PDL1. Gastroenterology. 2019;156:1849–61. e1813.
Fidler IJ. The pathogenesis of cancer metastasis: the ‘seed and soil’ hypothesis revisited. Nat Rev Cancer. 2003;3:453–8.
Chambers AF, Groom AC, MacDonald IC. Dissemination and growth of cancer cells in metastatic sites. Nat Rev Cancer. 2002;2:563–72.
Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331:1559–64.
Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the Roots of Cancer. Cancer Cell. 2016;29:783–803.
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT, Zender L, et al. Yes-associated protein is an independent prognostic marker in hepatocellular carcinoma. Cancer. 2009;115:4576–85.
Bisso A, Filipuzzi M, Gamarra Figueroa GP, Brumana G, Biagioni F, Doni M, et al. Cooperation between MYC and beta-catenin in liver tumorigenesis requires Yap/Taz. Hepatology. 2020. https://doi.org/10.1002/hep.31120.
Dong J, Feldmann G, Huang J, Wu S, Zhang N, Comerford SA, et al. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell. 2007;130:1120–33.
Zhang X, Li Y, Ma Y, Yang L, Wang T, Meng X, et al. Yes-associated protein (YAP) binds to HIF-1alpha and sustains HIF-1alpha protein stability to promote hepatocellular carcinoma cell glycolysis under hypoxic stress. J Exp Clin Cancer Res. 2018;37:216.
Shi C, Cai Y, Li Y, Li Y, Hu N, Ma S, et al. Yap promotes hepatocellular carcinoma metastasis and mobilization via governing cofilin/F-actin/lamellipodium axis by regulation of JNK/Bnip3/SERCA/CaMKII pathways. Redox Biol. 2018;14:59–71.
Comoglio PM, Trusolino L, Boccaccio C. Known and novel roles of the MET oncogene in cancer: a coherent approach to targeted therapy. Nat Rev Cancer. 2018;18:341–58.
Zhang Y, Xia M, Jin K, Wang S, Wei H, Fan C, et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol Cancer. 2018;17:45.
Yu Y, Khan J, Khanna C, Helman L, Meltzer PS, Merlino G. Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators. Nat Med. 2004;10:175–81.
Xiang C, Chen J, Fu P. HGF/Met Signaling in Cancer Invasion: the Impact on Cytoskeleton Remodeling. Cancers (Basel). 2017;9:44.
Garcia-Vilas JA, Medina MA. Updates on the hepatocyte growth factor/c-Met axis in hepatocellular carcinoma and its therapeutic implications. World J Gastroenterol. 2018;24:3695–708.
Papaccio F, Della Corte CM, Viscardi G, Di Liello R, Esposito G, Sparano F, et al. HGF/MET and the immune system: relevance for cancer immunotherapy. Int J Mol Sci. 2018;19:3595.
Vejchapipat P, Tangkijvanich P, Theamboonlers A, Chongsrisawat V, Chittmittrapap S, Poovorawan Y. Association between serum hepatocyte growth factor and survival in untreated hepatocellular carcinoma. J Gastroenterol. 2004;39:1182–8.
Mizuguchi T, Nagayama M, Meguro M, Shibata T, Kaji S, Nobuoka T, et al. Prognostic impact of surgical complications and preoperative serum hepatocyte growth factor in hepatocellular carcinoma patients after initial hepatectomy. J Gastrointest Surg. 2009;13:325–33.
Bladt F, Friese-Hamim M, Ihling C, Wilm C, Blaukat A. The c-Met Inhibitor MSC2156119J Effectively Inhibits Tumor Growth in Liver Cancer Models. Cancers (Basel). 2014;6:1736–52.
Bardelli A, Corso S, Bertotti A, Hobor S, Valtorta E, Siravegna G, et al. Amplification of the MET receptor drives resistance to anti-EGFR therapies in colorectal cancer. Cancer Disco. 2013;3:658–73.
Ou SI, Govindan R, Eaton KD, Otterson GA, Gutierrez ME, Mita AC, et al. Phase I Results from a Study of Crizotinib in Combination with Erlotinib in Patients with Advanced Nonsquamous Non-Small Cell Lung Cancer. J Thorac Oncol. 2017;12:145–51.
Dong Q, Du Y, Li H, Liu C, Wei Y, Chen MK, et al. EGFR and c-MET Cooperate to Enhance Resistance to PARP Inhibitors in Hepatocellular Carcinoma. Cancer Res. 2019;79:819–29.
Chen R, Zhu S, Fan XG, Wang H, Lotze MT, Zeh HJ 3rd, et al. High mobility group protein B1 controls liver cancer initiation through yes-associated protein -dependent aerobic glycolysis. Hepatology. 2018;67:1823–41.
Gavini J, Dommann N, Jakob MO, Keogh A, Bouchez LC, Karkampouna S, et al. Verteporfin-induced lysosomal compartment dysregulation potentiates the effect of sorafenib in hepatocellular carcinoma. Cell Death Dis. 2019;10:749.
Chen J, Du F, Dang Y, Li X, Qian M, Feng W. et al. Fibroblast Growth Factor 19-Mediated Up-regulation of SYR-Related High-Mobility Group Box 18 Promotes Hepatocellular Carcinoma Metastasis by Transactivating Fibroblast Growth Factor Receptor 4 and Fms-Related Tyrosine Kinase 4. Hepatology. 2020;71:1712–31.
Acknowledgements
Research was supported by grants from the National Key Research and Development Program of China 2018YFC1312103 (LX), National Natural Science Foundation of China No. 81871911 (WH), No. 81972237 (LX), and No. 81772623 (LX).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
He, Q., Lin, Z., Wang, Z. et al. SIX4 promotes hepatocellular carcinoma metastasis through upregulating YAP1 and c-MET. Oncogene 39, 7279–7295 (2020). https://doi.org/10.1038/s41388-020-01500-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41388-020-01500-y
- Springer Nature Limited
This article is cited by
-
Metastasis and basement membrane-related signature enhances hepatocellular carcinoma prognosis and diagnosis by integrating single-cell RNA sequencing analysis and immune microenvironment assessment
Journal of Translational Medicine (2024)
-
The Hippo-YAP signaling pathway drives CD24-mediated immune evasion in esophageal squamous cell carcinoma via macrophage phagocytosis
Oncogene (2024)
-
Comprehensive DNA methylation profiling of COVID-19 and hepatocellular carcinoma to identify common pathogenesis and potential therapeutic targets
Clinical Epigenetics (2023)
-
MESP2 binds competitively to TCF4 to suppress gastric cancer progression by regulating the SKP2/p27 axis
Cell Death Discovery (2023)
-
Combination of SIX4-siRNA and temozolomide inhibits the growth and migration of A-172 glioblastoma cancer cells
Naunyn-Schmiedeberg's Archives of Pharmacology (2023)