
Abstract
Smad and STAT proteins are critical signal transducers and transcription factors in controlling cell growth and tumorigenesis. Here we report that the STAT3 signaling pathway attenuates transforming growth factor-β (TGF-β)-induced responses through a direct Smad3–STAT3 interplay. Activated STAT3 blunts TGF-β-mediated signaling. Depletion of STAT3 promotes TGF-β-mediated transcriptional and physiological responses, including cell cycle arrest, apoptosis and epithelial-to-mesenchymal transition. STAT3 directly interacts with Smad3 in vivo and in vitro, resulting in attenuation of the Smad3–Smad4 complex formation and suppression of DNA-binding ability of Smad3. The N-terminal region of DNA-binding domain of STAT3 is responsible for the STAT3–Smad3 interaction and also indispensable for STAT3-mediated inhibition of TGF-β signaling. Thus, our finding illustrates a direct crosstalk between the STAT3 and Smad3 signaling pathways that may contribute to tumor development and inflammation.
Similar content being viewed by others

Introduction
Transforming growth factor-β (TGF-β) is a multifunctional cytokine that regulates diverse cellular responses, including apoptosis, cell growth inhibition and immune surveillance.1, 2, 3 Owing to its potent tumor-suppressive effects, the TGF-β signaling pathway is often inactivated in numerous cancers such as colon, pancreatic and gastric cancers, yet such somatic mutations are rare in other cancers such as breast, prostate and skin cancers.4, 5, 6, 7 Because loss of TGF-β responses is common in all types of cancers, there are alternative mechanisms underlying TGF-β resistance in those cancer types without somatic mutations in the TGF-β pathway. Recent progress suggests that activation of oncogenes can suppress a TGF-β growth inhibitory response.8, 9, 10, 11
TGF-β signals through a heteromeric complex of cell-surface serine–threonine kinase receptors, that is TβRI and TβRII, and intracellular signal transducers Smad2 and Smad3. In response to TGF-β ligands, TβRII transphosphorylates TβRI, which in turn mediates phosphorylation of Smad2 and Smad3. Phosphorylated (activated) Smad2/3 associates with Smad4, and translocates to the nucleus to regulate gene expression.3, 12 Each step of this signal transduction pathway can be regulated by a variety of intracellular factors.13 For example, Erk MAP kinase-dependent phosphorylation of the linker region impairs Smad3 nuclear accumulation.14 In the nucleus, many transcription factors interact with R-Smads to modulate the final gene transcription output.15, 16 The majority of these cooperative partners are also components of other signaling pathways. For example, FAST1/2 and c-Jun/c-Fos cooperate with Smad2–Smad417, 18 and Smad3–Smad4 complexes,19 respectively, to regulate transcription. The cooperative or antagonistic interactions of Smads with other transcriptional factors depend on the physiological contexts and therefore dictate the final physiological outputs.
STAT3 is a common downstream effector of some cytokines overwhelmingly expressed in the tumor environment such as interleukin (IL)-6, IL-11 and vascular endothelial growth factor.20 Moreover, STAT3 is also a critical mediator of epidermal growth factor receptor signaling, which is aberrantly activated in various tumors.20 Therefore, STAT3 is a point of convergence for numerous oncogenic signaling pathways, and this makes it a potential candidate to modulate TGF-β signaling in tumor progression. Several earlier studies have demonstrated two-way interplays between STAT3 and TGF-β signaling pathways. For instance, STAT3 can promote TGF-β1 expression to enhance hepatic fibrosis in hepatocellular carcinoma development;21 activated STAT3 also induces Smad7 expression to desensitize TGF-β signaling.22 TGF-β conversely inhibits IL-6-mediated STAT3 activation and affects its target gene expression.23, 24 Smads can also attenuate the STAT3-mediated pathway by inhibiting its DNA-binding ability and cooperation with p300.25 However, many of these reports are conflicting and the underlying mechanisms behind these observations have not been elucidated.
In this study, we identified and characterized a direct interaction between STAT3 and Smad3. Cellular and molecular evidence lead to the conclusion that the STAT3–Smad3 interaction contributes to STAT3-mediated inhibition of TGF-β signaling. These findings elucidate a novel mechanism underlying the crosstalk between TGF-β antiproliferative signaling and STAT3-dependent growth-promoting signaling pathways.
Results
STAT3 is essential for EGF/IL-6-mediated desensitization of the TGF-β responses
TGF-β and epidermal growth factor (EGF) signaling pathways are both independently implicated as key regulators in tumor formation and progression. Previous studies have shown that EGF may exert as an antagonist of TGF-β signaling in several cellular contexts,26, 27 implying a potential crosstalk between these two signaling pathways. To elucidate mechanisms underlying EGF-mediated regulation of TGF-β signaling, we firstly investigated and confirmed the inhibitory effect of EGF in HaCaT cells. HaCaT is a human keratinocyte cell line that is highly responsive to TGF-β. A Smad3-dependent luciferase reporter CAGA-Luc containing 12 copies of Smad-binding elements (SBE) was transiently transfected into HaCaT cells to quantitatively determine TGF-β signaling sensitivity. Notably, TGF-β-activated stimulation of CAGA-Luc activity was significantly reduced in cells treated with EGF (Figure 1a). This inhibitory effect of EGF on TGF-β signaling is not restricted to HaCaT cells, as EGF exerted a consistently inhibitory role in human hepatoma HepG2 cells (Figure 1b). It was known that IL-6 and EGF stimulate similar downstream signaling pathways such as the JAK/STAT and Ras/MEK/ERK pathways. Aberrant IL-6/gp130 signaling also causes the desensitization of TGF-β signaling in the gastric epithelium.22 Thus, we also examined the effect of IL-6 on TGF-β signaling. Results showed that similarly to EGF, IL-6 decreased TGF-β-meditated CAGA-Luc expression (Figure 1b).

STAT3 is essential for EGF/IL-6-mediated desensitization of the TGF-β responses. (a) EGF suppresses TGF-β-induced CAGA-Luc activity in HaCaT cells. HaCaT cells were transfected with the TGF-β-responsive reporter plasmid CAGA-Luc, and then treated with TGF-β (2 ng/ml) in the presence or absence of EGF (10 ng/ml). TGF-β/EGF treatment and luciferase assays were carried out as described in the Materials and methods section. (b) IL-6 and EGF suppress TGF-β-induced CAGA-Luc activity in HepG2 cells. Experiment was carried out as described in (a). IL-6 was used at a concentration of 10 ng/ml. (c) EGF-mediated inhibition on the TGF-β SBE-luc response is MEK independent. HaCaT cells were transfected with SBE-Luc reporter plasmid, pretreated with or without MEK inhibitor PD98059 (50 μm) for 4 h, and then treated with TGF-β (2 ng/ml) and/or EGF (10 ng/ml) for another 12 h. Luciferase assays were carried out as described in (a). (d) Stable knockdown of STAT3 in HaCaT cells. The protein level of STAT3 was detected by using western blotting. (e) EGF-mediated inhibition on the CAGA-Luc response is STAT3 dependent. Transfection, TGF-β/EGF treatment and luciferase assays in STAT3-depleted stable cells and control cells were carried out as described in the Materials and methods section. (f) IL-6-mediated inhibition of the TGF-β-induced p21 transcription is STAT3 dependent. HaCaT STAT3-depleted stable cells and control cells were treated with TGF-β (2 ng/ml) and/or IL-6 (10 ng/ml), and total RNAs were extracted for quantitative reverse transcription polymerase chain reaction or quantitative real-time polymerase chain reaction (qRT–PCR) analysis of p21 mRNA. (g) IL-6-mediated inhibition of the TGF-β-induced PAI-1 transcription is STAT3 dependent. HaCaT cells were transfected with STAT3 siRNAs, treated with TGF-β (2 ng/ml) and/or IL-6 (10 ng/ml), and total RNAs were extracted for qRT–PCR analysis of PAI-1 mRNA. Abbreviation: RLU, relative light units.
The Ras–MAPK pathway, one of the most documented pathways activated by EGF, has been implicated in modulating Smad activation.14 To examine whether this pathway was solely involved in EGF-mediated desensitization of TGF-β signaling, we used a pharmacological inhibitor PD98059 to block MEK activity. PD98059 was not able to block EGF-mediated inhibition of TGF-β-induced SBE-Luc reporter activity (Figure 1c), suggesting that the desensitization of TGF-β pathway by EGF is not through the MEK/ERK signaling.
Another important downstream signaling stimulated by EGF was the JAK/STAT pathway. It has been shown that sustained STAT3 activation results in the inhibition of TGF-β signaling.22 Therefore, we speculated whether STAT3 was involved in EGF-mediated desensitization of TGF-β responses. We established a HaCaT cell line with stable STAT3 knockdown (STAT3-KD) and investigated the effect of STAT3 depletion on TGF-β-induced responses. The protein levels of STAT3 were efficiently reduced in STAT3-KD cells (Figure 1d). Stable knockdown of STAT3 increased TGF-β-activated CAGA-Luc activity and impaired the inhibitory effects of EGF stimulation (Figure 1e). More apparent attenuation of TGF-β response by IL-6 was observed when the TGF-β-induced mRNA level of target p21 was analyzed, and furthermore this attenuation was lost when STAT3 was knocked down (Figure 1f). Similar result was obtained by using transient tranfection. Two sequence-specific siRNAs against STAT3, which could effectively reduce STAT3 protein levels (data not shown), enhanced TGF-β-induced PAI-1 mRNA expression, and partially diminished IL-6-mediated inhibition (Figure 1g). Taken together, these results demonstrate that EGF/IL-6-mediated suppression of TGF-β pathway is dependent of STAT3.
Activated STAT3 suppresses TGF-β-induced growth inhibitory responses
After having shown that STAT3 is involved in EGF/IL-6-mediated inhibition of TGF-β signaling, we next set out to examine the effect of activated STAT3 on TGF-β-induced responses. STAT3 or its two variants was transiently transfected into TGF-β-responsive HaCaT cells. While STAT3C is a constitutively active mutant of STAT3 and can mimic the aberrant STAT3 activation in cancer cells,22 STAT3DN harbors a Y705F mutation that acts as a dominant-negative mutant.22 As shown in Figure 2a, STAT3C strongly decreased TGF-β-activated CAGA-Luc activity in a dose-dependent manner in HaCaT cells. In contrast, STAT3 wild type (WT) had a very weak inhibition, and STATDN has no inhibition at all on TGF-β signaling. Because TGF-β is a potent inhibitor of cell proliferation by arresting epithelial cells in the G1 phase, we reasoned that STAT3 would desensitize cells to TGF-β-mediated growth inhibition. We established a HaCaT cell line to stably express STAT3C, and then determined the expression level of STAT3C which was found to be comparable to that of endogenous STAT3 (Figure 2b). As key effectors and indicators for TGF-β-mediated cell cycle arrest, induction of CDK inhibitors p15 and p21 was evaluated in HaCaT-STAT3C cells. While TGF-β induced 11- and 4-fold increase in p15 and p21 mRNA levels, respectively, STAT3C clearly attenuated this TGF-β-dependent transcriptional induction (Figures 2c and d). Analysis of protein levels also supported the suppressing effect of STAT3C on TGF-β-dependent p15 and p21 induction (Figure 2e). In accordance, expression of STAT3C caused a decrease in the G1 phase of the cell cycle, accompanied by an increase in the S phase, when the TGF-β response were compared in HaCaT-STAT3C cells vs control cells (Figure 2f). Consistently, STAT3C decreased the activity of TGF-β to suppress proliferation of HaCaT cells (Figure 2g).

Activated STAT3 suppresses TGF-β-induced growth inhibitory responses. (a) STAT3C potently attenuates TGF-β-induced reporter expression in a dose-dependent manner. HaCaT cells were transfected with increasing amounts of STAT3 wild type (WT) or variants (STAT3C or STAT3DN), together with CAGA-Luc, and then treated with or without TGF-β (2 ng/ml). (b) Stable expression of STAT3C in HaCaT cells. The protein level of STAT3 was detected by using western blotting. Note that the protein level of exogenous Flag-STAT3C is similar to that of endogenous STAT3. (c) Stable expression of STAT3C decreases TGF-β-induced p15 mRNA expression in HaCaT cells. Cells were treated with or without TGF-β (2 ng/ml) for 2 h, and total RNA was extracted for quantitative reverse transcriptase polymerase chain reaction analysis. (d) STAT3C decreases TGF-β-induced p21 mRNA expression in HaCaT cells. (e) Stable expression of STAT3C decreases TGF-β-induced p15 and p21 protein expression in HaCaT cells. Cells were treated with TGF-β for 0, 2, 4 and 8 h, respectively, and then the protein levels of p15 and p21 were detected by using western blotting. (f) STAT3C promotes G1–S progression in TGF-β-treated cells. STAT3C cells and control cells were treated with TGF-β (2 ng/ml) for 2 days. Cell cycle progression was determined by flow cytometry as described in the Materials and methods section. (g) STAT3C interferes with TGF-β-mediated inhibition on cell proliferation. STAT3C stable cells and control cells were treated with TGF-β, and cell growth rates were examined by actual cell numbers being counted at indicated time point (0, 1 and 2 days) of TGF-β treatment. Abbreviation: RLU, relative light units.
Loss of STAT3 enhances TGF-β-induced physiological responses
To elucidate the physiological functions of endogenous STAT3 in regulating TGF-β signaling, we examined the effect of STAT3 depletion on TGF-β responses in HaCaT cells. We found that shRNA-mediated stable knockdown of STAT3 expression resulted in a significant increase in the TGF-β-induced mRNA levels of p15 (Figure 3a) and p21 (Figure 3b). TGF-β-induced protein level of p15 was similarly enhanced by STAT3 depletion (Figure 3c). In supporting these results, cell proliferation analysis showed that enhanced p15/p21 induction correlated to sensitization of cells to cell cycle arrest induced by a low dose of TGF-β (Figure 3d).

Loss of STAT3 enhances TGF-β-induced physiological responses. (a) Depletion of STAT3 increases TGF-β-induced p15 mRNA expression in HaCaT cells. Cells were treated with or without TGF-β (2 ng/ml) for 2 h, and total RNA was extracted for quantitative reverse transcriptase polymerase chain reaction analysis. (b) Depletion of STAT3 increases TGF-β-induced p21 mRNA expression in HaCaT cells. (c) Depletion of STAT3 enhances TGF-β-induced p15 protein expression in HaCaT cells. Cells were treated with TGF-β for 0, 4 and 8 h, respectively, and then the protein level of p15 was detected by using western blotting. (d) Depletion of STAT3 in HaCaT cells enhances TGF-β-induced growth arrest. STAT3 knockdown stable cells and control cells were treated with TGF-β, and cell growth rates were examined by actual cell numbers being counted at indicated time point (0, 1 and 2 days) of TGF-β treatment (1 ng/ml). (e) Stable knockdown of STAT3 in Hep3B cells. The protein level of STAT3 was detected by using western blotting. (f) Depletion of STAT3 enhances TGF-β-induced cell apoptosis. STAT3 knockdown stable cells and control cells were treated with TGF-β at indicated concentrations. Apoptotic cells were analyzed by flow cytometry. STAT3 knockdown stable cells and control cells were treated with or without TGF-β (2 ng/ml) for 48 h. (g) Apoptotic nuclei and DNA fragments were visualized by nuclei staining. (h) Quantification of apoptotic cells in panel (g). (i) Depletion of STAT3 enhances TGF-β-induced EMT marker expression. STAT3 knockdown stable cells and control cells were treated with or without TGF-β (2 ng/ml) for 36 h, and the protein levels of E-cadherin, N-cadherin and Fibronectin were detected by western blotting. (j) Depletion of STAT3 enhances TGF-β-mediated EMT. STAT3 knockdown stable cells and control cells were treated with TGF-β (2 ng/ml) for 0, 24 and 48 h, respectively. Cells were fixed and immuno-stained with anti-E-cadherin or stained with Phalloidin, and imaged by a confocal microscope. (k) Depletion of STAT3 promotes cell migration. STAT3 knockdown stable cells and control cells were grown to confluence, followed by a scratch on the cell patch. The status of cell gap closure was recorded at 24 h post-wound in the presence or absence of TGF-β (2 ng/ml).
In addition to the cell cycle arrest, the tumor suppressing function of TGF-β also comes from its role in apoptosis. A good cell model is the human hepatoma Hep3B cells that undergo apoptosis when exposed to TGF-β. To this end, we established a stable cell line in Hep3B cells with STAT3 knockdown (STAT3-KD) (Figure 3e) and investigated the effect of STAT3 depletion on TGF-β-induced apoptosis. In control cells, while TGF-β at a concentration of 200 pm induced apoptosis at a rate of 10%, the effect of a low concentration of TGF-β (5 pm) was not detectable (Figure 3f). Notably, STAT3-KD cells underwent apoptosis with much increased sensitivity to TGF-β as 5 pm of TGF-β could elicit an apparent apoptosis response and 200 pm of TGF-β further increased apoptosis rate (Figure 3f). We further examined condensed chromatin and nuclear fragmentation, two typical properties of apoptotic cell nuclei, by using Hoechest 33258 staining. The staining result showed that STAT3 depletion markedly sensitized cells to TGF-β-mediated apoptosis (Figures 3g and h).
As a multifunctional cytokine, TGF-β has a dual role in tumorigenesis. Whereas TGF-β induces cell cycle arrest and apoptosis in epithelial cells and early tumor cells, it promotes epithelial-to-mesenchymal transition (EMT), cell motility and tumor cell metastasis. Thus, we also sought to examine the role of STAT3 in TGF-β-induced EMT. The defined typical characteristics of cells undergoing EMT are downregulation of E-cadherin (an epithelial marker), upregulation of N-cadherin and Fibronectin (mesenchymal markers), and reorganization of actin to stress fiber. As shown in Figure 3i, whereas STAT3 depletion abolished expression of E-cadherin, it enhanced TGF-β-induced expression of EMT-associated proteins such as N-cadherin and Fibronectin in HaCaT cells. Loss of epithelial characteristics and acquisition of mesenchymal features were also analyzed by confocal microscopy. Compared with controls, STAT3 depletion strengthened EMT-associated changes in both unstimulated and TGF-β-stimulated cells. The effect of STAT3 was already apparent at an early time point of TGF-β treatment (24 h). At 24 h, wild-type HaCaT cells exhibited little changes in EMT markers and morphology (Figure 3j). Depletion of STAT3 expression resulted in a dramatic loss of E-cadherin and gain of F-actin fiber (Figure 3j). Consistent with the changes in cell morphology, STAT3-KD cells exhibited stronger migratory ability when treated with TGF-β in the wound healing analysis (Figure 3k). These results support the notion that STAT3 also suppresses the EMT and migratory responses of TGF-β.
STAT3 selectively attenuates the formation of Smad3 signaling complexes
To elucidate the molecular mechanism underlying STAT3-mediated suppression of TGF-β signaling, we tested whether STAT3 could affect activation and signaling capacity of Smads, including Smad phosphorylation, nuclear localization, complex formation and occupancy on the target gene promoters. By examining TGF-β-induced Smad3 phosphorylation, we found that stable knockdown of STAT3 did not obviously influence TGF-β-induced phosphorylation of endogenous Smad3 in HaCaT cells (Figure 4a), suggesting that STAT3 probably does not regulate the expression or activity of TGF-β ligands or receptors in HaCaT cells. Moreover, STAT3 had no effect on TGF-β-induced Smad3 nuclear accumulation (Figure 4b). Interestingly, TGF-β treatment stimulated not only the nuclear accumulation of Smad3 but also appeared to enhance that of STAT3 and thus co-localization of the two proteins in the nucleus (Figure 4b).

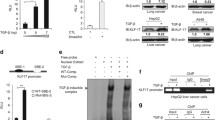
STAT3 selectively attenuates the formation of Smad3 signaling complexes. (a) Depletion of STAT3 does not affect TGF-β-induced phosphorylation of Smad3. STAT3 knockdown stable cells and control cells were treated with or without TGF-β (2 ng/ml) for 1 h. The protein levels of p-Smad3, Smad3 and STAT3 were detected by using western blotting. (b) STAT3C does not alter TGF-β-stimulated nuclear accumulation of Smad3. STAT3C stable cells were treated with or without TGF-β (2 ng/ml) for 1 h, fixed and immuno-stained with anti-Smad3 and anti-Flag antibodies. DNA was stained with DAPI. (c) STAT3C attenuates the interaction between Smad3 and Smad4. HEK293T cells were transfected with indicated plasmids. Levels of these proteins in immunoprecipitation products and whole-cell lysates were analyzed by western blotting. (d) STAT3C disrupts the endogenous Smad3–Smad4 complex formation. STAT3C stable cells and control cells were treated with or without TGF-β (2 ng/ml). Cell lysates were immunoprecipitated with anti-Smad4 antibody or control IgG antibody. The immuno-complexes and input were analyzed by using western blotting with indicated antibodies. (e) STAT3C, but not STAT3DN, inhibits the transactivation activity of Smad3. HaCaT cells were transfected with various amounts of STAT3C or STAT3DN together with pFR-luc, Gal4-dbd or Gal4-Smad3, and treated with TGF-β (2 ng/ml) for 8 h. Luciferase assay was performed as described in the Materials and methods section. (f) STAT3C decreases DNA-binding activity of Smad3. HEK293T cells were transfected with indicated plasmids. Cell lysates were immunoprecipitated with biotinylated SBE and streptavidin beads. DNA-bound Smad3 was then assessed by using western blotting. (g) STAT3 inhibits Smad3 binding to the PAI-1 promoter. Isolated chromatins from control and STAT3C cells (treated with or without TGF-β) were immunoprecipitated with IgG or anti-Smad3 antibody for chromatin immunoprecipitation assay. Specific fragments of the PAI-1 promoter DNA were determined by polymerase chain reaction. (h) STAT3 depletion increases Smad3 binding to the PAI-1 promoter. The experiments were carried out as described in (g). (I) STAT3 depletion increases Smad3 binding to the p15 promoter. The experiments were carried out as described in (g). Abbreviations: DAPI, 4',6-diamidino-2-phenylindole; WCL, whole-cell lysate.
We next investigated the effect of STAT3 on the TGF-β-induced Smad complex formation. We found that STAT3C could compete with Smad4 for Smad3 binding as it inhibited TGF-β receptor-mediated Smad3–Smad4 interaction in HEK293T cells (Figure 4c). Furthermore, stable expression of STAT3C abolished the endogenous Smad3–Smad4 complex in HaCaT cells (Figure 4d). Notably, this STAT3C-induced disruption of the Smad complex was selective as it affected the Smad3–Smad4, but not Smad2–Smad4 association (Figure 4d). Because the transcriptional activity of Smad3 depends on the activator role of Smad4,28 we examined the effect of STAT3C on Smad3-mediated transcriptional activation using the heterologous Gal4 system. TGF-β stimulation could induce transcription activity of Gal4-fused Smad3, which was reflected by expression of the Gal4-driven luciferase reporter FR-Luc (Figure 4e). STAT3C dramatically inhibited transcriptional activity of Gal4-Smad3 (Figure 4e). In contrast, a dominant-negative mutant of STAT3 (STAT3DN) did not exhibit any inhibitory effect (Figure 4e).
A few transcription factors modulate the activity of Smad3 to bind to DNA in either synergistic or antagonistic manner.15 Therefore, we explored the possibility that STAT3 influences the DNA-binding activity of Smad3. In DNA pull-down assays, where a biotinylated DNA fragment consisting of SBEs was used to precipitate Smad3 in HEK293T cells, activated Smad3 (by TβRI-T202D) exhibited a high level of DNA binding (Figure 4f). In contrast, STAT3C markedly reduced the DNA-binding level of Smad3 (Figure 4f). These results further prompted us to investigate the inhibitory effect of STAT3 on the natural promoters of two representative TGF-β target genes. Occupancy of Smad3 on the promoter regions of p15 and PAI-1 were examined by using chromatin immunoprecipitation. TGF-β induced the enrichment of Smad3 on the PAI-1 promoter, and this enrichment was abolished by STAT3C (Figure 4g). Conversely, depletion of STAT3 profoundly increased TGF-β-induced accumulation of Smad3 on the promoters of PAI-1 (Figure 4h) and p15 (Figure 4i). Taken together, STAT3 impairs the Smad3–Smad4 complex formation and decreases Smad3 binding to chromatin, thereby resulting in inhibition of Smad3-mediated transcriptional activation.
STAT3 selectively and directly interacts with Smad3 under physiological conditions
Based on the convincing evidence that STAT3 attenuates the transcriptional capacity of Smad3, we speculated that STAT3 might directly interact with Smad3. We analyzed the potential STAT3–Smad3 interactions by using co-immunoprecipitation (co-IP) and in vitro pull-down assays. In HEK293T cells transfected with expression plasmids for Flag-tagged Smad3 and Myc-tagged STAT3WT or STAT3C, we could detect both STAT3WT and STAT3C in the anti-Flag-Smad3 immunoprecipitates (Figure 5a).

STAT3 directly interacts with Smad3 under physiological conditions. (a) STAT3 interacts with Smad3 in HEK293T cells. HEK293T cells were transfected with Myc-Smad3 and Flag-STAT3WT (wild-type STAT3), Flag-STAT3C (constitutively active STAT3) or Flag-STAT3DN (dominant-negative mutant of STAT3). Levels of these proteins in IP products and whole-cell lysates were analyzed by western blotting. (b) STAT3 interacts with endogenous Smad3. STAT3C-expressing HaCaT stable cells were treated with or without TGF-β (2 ng/ml) for 1 h. Cell lysates were immunoprecipitated with anti-Flag antibody or control IgG antibody. The immune-complexes and input were analyzed by western blotting with indicated antibodies. (c) STAT3 interacts with Smad3 with both proteins at endogenous levels. HaCaT cell lysates were immunoprecipitated with anti-Smad3 antibody or control IgG antibody. The immune-complexes and input were analyzed by western blotting with indicated antibodies. (d) STAT3 directly binds to Smad3 in vitro. In vitro binding was carried out with purified GST or GST-Smad3 and in vitro translated STAT3C. (e) Schematic diagram of STAT3 and its deletion mutants. Individual domains of STAT3 are shown. (f) Smad3 binds with the DNA-binding domain of STAT3. Experiments were carried out as descried in (a). (g) Schematic diagram of deletion mutants of DNA-binding domain of STAT3. (h) Smad3 binds with the N-terminal region of STAT3 DNA-binding domain. Experiments were carried out as descried in (a). (i) Schematic diagram of Smad3 and its deletion mutants. (j) STAT3 binds with the MH2 domain of Smad3. Experiments were carried out as descried in (a). Abbreviations: IP, immunoprecipitation; WCL, whole-cell lysate..
To further investigate the specificity and physiological relevance of the STAT3–Smad3 interaction, we analyzed the STAT3–Smad3 interaction under physiological conditions. We first used co-IP experiments to examine the association between endogenous Smad2/3 and stably expressed Flag-STAT3C, the level of which is comparable to that of endogenous STAT3 (Figure 2b). As shown in Figure 5b, anti-Flag immunoprecipitates (STAT3C) specifically retrieved Smad3, but not Smad2. This specificity was consistent with the result that STAT3 inhibited binding of Smad3, but not Smad2, to Smad4. We further examined the endogenous STAT3–Smad3 interaction in HaCaT cells, and found that STAT3 could be detected in the anti-Smad3 immunoprecipitates, but not in that of control IgG (Figure 5c).
To evaluate whether the STAT3–Smad3 interaction is direct, we conducted an in vitro interaction assay where only recombinant proteins were used. Smad3 was expressed and purified from Escherichia coli as a glutathione S-transferase (GST) fusion protein, whereas STAT3C was obtained from in vitro coupled transcription/translation in rabbit reticulocyte lysate. As shown in Figure 5d, in vitro synthesized STAT3C was retrieved by GST-fused Smad3 protein, but not GST alone, indicating that STAT3 directly interacts with Smad3. Taken together, STAT3 directly interacts with Smad3 under physiological conditions.
To determine the structural features for STAT3–Smad3 interaction, we first mapped the region in STAT3 that mediates the STAT3–Smad3 interaction. STAT3 consists of several protein–protein interaction domains including coil–coil (CC), DNA-binding domain (DBD), and Src homology 2 (SH2) domains. Interaction of Smad3 with each of these individual domains of STAT3 was assessed by using co-IP assays (Figure 5e). As shown in Figure 5f, DBD of STAT3 strongly bound to Smad3, whereas all other domains did not bind to Smad3. To further narrow down the interacting region in the DBD, three truncated mutants were created. While STAT3C-ΔDBD lacks the entire DBD, STAT3C-ΔDBDc and STAT3C-ΔDBDn lack the C- and N-terminal regions of the STAT3 DBD, respectively (Figure 5g). It is apparent that the N-terminal half of DBD was critical for the STAT3–Smad3 interaction (Figure 5h). We then determined the domains of Smad3 for STAT3 binding. Smads are structurally conserved proteins consisting of MH1 domain in the N terminus and MH2 domain in the C terminus, linked with a relatively less conserved linker region (Figure 5i). Our co-IP binding assay found that Smad3 mutants with deletion of either the MH1 domain or the linker, but not the MH2 domain, retained the ability to bind with STAT3 (Figure 5j, lanes 2 and 3). Notably, deletion of the MH2 domain completely abolished Smad3 binding with STAT3. These results suggest that STAT3 binds to the MH2 domain of Smad3.
Binding to Smad3 is indispensable for STAT3 to inhibit TGF-β responses
We further assessed the importance of the Smad3-interacting region of STAT3 in STAT3-mediated suppression of TGF-β signaling. In HaCaT cells, both the full-length STAT3C and STAT3C-ΔDBDc retained the inhibitory effect on TGF-β-induced CAGA-Luc activity. In contrast, STAT3C-ΔDBDn, which failed to interact with Smad3, was unable to suppress TGF-β signaling (Figure 6a). Similarly, only the full-length STAT3C and STAT3C-ΔDBDc could obviously attenuate Smad3-induced transcriptional activation (Figure 6b). Therefore, binding to Smad3 is essential for STAT3 to inhibit TGF-β signaling.

STAT3 requires a direct interaction with Smad3 to antagonize TGF-β signaling. (a) STAT3-ΔDBDn mutant fails to inhibit TGF-β-induced CAGA-Luc reporter activity. Experiments were carried out as descried in Figure 1a. (b) STAT3-ΔDBDn mutant fails to inhibit Smad3-activated CAGA-Luc reporter activity. Experiments were carried out as descried in Figure 1a. (c) A working model for STAT3-mediated repression of Smad-induced transcription. In normal cells, Smad3 mediates TGF-β growth inhibitory and transcriptional responses. In tumors where STAT3 is aberrantly activated, STAT3 sequesters Smad3 from the Smad nucleoprotein complex and thus suppresses TGF-β growth inhibitory and transcriptional responses.
Discussion
Cancer and inflammation are two reciprocally regulated events. Malignant cells can usually generate a local inflammatory environment, and conversely, inflammatory conditions will further promote oncogenic transformation. Increased expression of soluble cytokines and growth factors establish and maintain a tumor microenvironment where tumor cells, stroma cells and inflammatory cells are associated. TGF-β is recognized as a major mediator in either tumor-associated inflammation or inflammation-associated tumor progression. Crosstalks between TGF-β and other cytokines may play an important role in the diverse functions of TGF-β. There are several studies, albeit conflicting, reporting crosstalks between TGF-β and STAT3 signaling pathways. TGF-β reduces phosphorylation of STAT3, and then downregulates the expression of its downstream genes.23, 24, 25, 29, 30 Conversely, STAT3 induces the expression of TGF-β1 to promote fibrosis.21 It has also been previously reported that there are p300 coactivator-dependent interactions between STAT3 and Smad3,31 between STAT1 and Smad332 and between STAT3 and Smad1.33 However, how STAT3 growth-promoting signaling antagonizes TGF-β growth inhibitory signaling remains elusive.
In the present study, we have identified a direct physical and functional interaction between STAT3 and Smad3. The interaction between STAT3 and Smad3 in hepatocytes was previously reported, yet it was indirect as it was bridged by p300.31 We for the first time demonstrate that the STAT3–Smad3 interaction is direct by means of in vitro interaction assays. The interaction enables STAT3 to compete with Smad4 for Smad3, thereby disrupting the Smad3–Smad4 complex, and thus attenuate TGF-β-mediated Smad3-dependent growth inhibitory and transcriptional responses in epithelial cells (Figure 6c), which is in contrast to the cooperative action of STAT3–Smad3 in STAT3-mediated gene expression in hepatocytes.31 The current finding is consistent with the oncogenic property of STAT3 and growth suppressor role of the TGF-β signaling. Loss of TGF-β tumor-suppressing response, which is a hallmark in cancer,34 can be achieved through somatic mutations in the genes encoding components of the TGF-β tumor-suppressing response pathway. Indeed, mutations are frequent in the Smad4 or TβRII gene in the gastrointestinal cancers such as pancreatic and colorectal cancers.4, 6, 35 In addition to genetic lesions in the genes encoding Smads and TGF-β receptors, accumulating evidence has demonstrated that the tumor suppressor functions of Smads are compromised by oncogene products such as c-Ski, Bcl6, c-Myc and Evi-1 through direct Smad–oncoprotein interactions.8, 9, 10, 11 Overexpression or amplification of STAT3 is often observed in cancers.20 Thus, our study extends and expands the role of oncoproteins in suppressing the TGF-β tumor-suppressing response.
It is previously reported that hyperactive STAT3 induces expression of inhibitory Smad7, and thus desensitizes TGF-β responses.22, 27 However, there is a minimal induction of Smad7 mRNA by STAT3 overexpression in HaCaT cells, which cannot explain the profound suppressing effect of STAT3 on TGF-β responses (data not shown). STAT3 is one of the effectors mediating the growth-promoting effect of EGF, IL-6 and related factors. Indeed, EGF and IL-6 have suppressing effects on TGF-β-mediated transcriptional responses. We rule out the possibility that the effect of EGF may be mediated via an Erk-dependent manner. Although it is known that EGF-activated Erks mediate hyper-phosphorylation of Smad2/3 that can affect the nuclear translocation of Smad2/3,14 MEK1 inhibitor PD98059 does not reverse the EGF-mediated attenuation of TGF-β signaling in our system. Neither does STAT3 alter nuclear accumulationof Smad3. Mechanistically, STAT3 competes with Smad4 for direct Smad3 binding and disables Smad3 to bind to DNA and to form the hetero-oligomeric complex with Smad4. As a consequence, STAT3 attenuates TGF-β-mediated growth inhibitory and transcriptional responses. Consistently, STAT3-depleted cells exhibit enhanced TGF-β-mediated responses.
Our current study has important implication in the development of more effective targeted cancer therapies, since TGF-β and JAK/STAT signaling have critical functions in tumor cell behaviors and tumor microenvironment. Our study provides compelling evidence demonstrating that STAT3 is a new oncogenic partner that interacts with Smads and suggests that STAT3 promotes tumorigenesis partly through disruption of the TGF-β pathway. Therefore, targeting of STAT3 with small molecules disrupting its canonical signaling in tumor cells may not be sufficient to halt tumor progression. Development of new STAT3 inhibitors targeting STAT3’s DBD, which confers both DNA binding and Smad3 binding, may provide an advantage for not only inhibiting STAT3’s canonical growth-promoting functions but also restoring TGF-β tumor-suppressing functions.
The observation that STAT3 selectively binds to Smad3, not Smad2, is very intriguing. Smad2 and Smad3 are the two closely related signal transducers for TGF-β/activin/myostatin in the TGF-β superfamily. Both are essential in most of TGF-β responses.15 However, while they share 92% sequence identity, Smad2 lacks the β-hairpin region that confers the DNA-binding function in Smad3. This DNA-binding difference may explain the differential roles of Smad2 and Smad3 in the regulation of certain TGF-β responses.36, 37, 38, 39, 40 Additionally, differential ability of Smad2 and Smad3 to bind to partner transcriptional factors can also contribute to differential TGF-β responses. Thus, the Smad3–STAT3 interaction may be involved in certain TGF-β responses. We previously reported that Smad2 is required for Th-17 cell differentiation, where loss of Smad3 leads to enhanced Th-17 differentiation.41, 42 Although this study only describes the antagonistic activity of STAT3 on the Smad3 signaling, it is conceivable that there is mutual inhibition between STAT3 and Smad3. Lastly, given its oncogenic role, the ability of STAT3 to differentially block Smad3 signaling further implicates that Smad3 is a key player in the TGF-β tumor suppressor signaling. This is consistent with the fact that loss of the Smad3 expression occurs in human cancer.43, 44
Besides tumor and inflammation contexts, the interplay between TGF-β signaling and STAT3-dependent signaling exist in various physiological contexts, such as Th-17 cell differentiation where TGF-β and IL-6 collaborate,45 and murine embryonic stem cell self-renewal and differentiation where LIF and TGF-β/BMP signaling cooperatively control.46, 47 How the direct interplay between STAT3 and Smad3 impacts these biological functions in vivo warrants more exciting investigation.
Materials and methods
Plasmids
Expression plasmids for Myc- or Flag-tagged Smads and T202D (constitutively active TGF-β type I receptor) were described previously.48 Myc- or Flag-tagged STAT3WT, STAT3C22 and STAT3DN 21 were generated by polymerase chain reaction and cloned into the EcoRI and SalI sites of pXF6F and pXF3HM. The pXF6F and pXF3HM were derived from pRK5 (Genentech, South San Francisco, CA, USA).
Antibodies
Antibodies against Smad2 (#5339), Smad3 (#9523), Smad2/3 (#8685), p-Smad2 (#3108), p-Smad3 (#9520), STAT3 (#9139), p-STAT3 (#9145), HA tag C29F4 (#3724), p21 (#2947) and E-cadherin (#3195) were purchased from Cell Signaling Technology (Danvers, MA, USA). Antibodies against Flag tag M2 (#F3165) and GAPDH (#G8795) were bought from Sigma (St Louis, MO, USA). Antibodies against Myc tag 9E10 (#sc-40) and Fibronectin (#sc-8422) were from Santa Cruz Biotechnology (Dallas, TX, USA). Antibodies against p15 (#C0287) were purchased from Assaybiotech (Sunnyvale, CA, USA). Antibody against N-cadherin (#610920) was from BD Biosciences (San Jose, CA, USA).
Cell culture and transfection
HEK293T cells were grown in Dulbecco’s modified Eagle’s medium (Corning, Tewksbury, MA, USA) with 10% fetal bovine serum (GIBCO, Grand Island, NY, USA). HaCaT, Hep3B and HepG2 cells were cultured in minimum essential medium (Corning) with 10% fetal bovine serum. HEK293T cells were transfected with polyethylenimine (Polyscience Inc., Warrington, PA, USA). HaCaT cells were transfected with X-treme GENE HP DNA transfection reagent (Roche, Basel, Switzerland). Hep3B and HepG2 cells were transfected with Lipofectamine2000 (Invitrogen, Carlsbad, CA, USA). Stable cell lines were selected by puromycin with appropriate concentrations.
Luciferase reporter assays
Reporters CAGA-Luc, SBE-Luc and 3TP-Luc were used to measure TGF-β-induced transcription. Cells were co-transfected with indicated reporter plasmids and a Renilla luciferase plasmid to normalize transfection efficiency. Briefly, 24–36 h after transfection, cells were treated with TGF-β (2 ng/ml), IL-6 (10 ng/ml) and EGF (10 ng/ml) for 12 h. Cells were harvested and measured by Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA). All assays were carried out in duplicates and the activities of firefly were normalized against Renilla luciferase activities.
RNA interference and real-time polymerase chain reaction
siRNAs targeting STAT3 were transfected into cells with Lipofectamine RNAiMAX reagent (Invitrogen). siRNAs were synthesized by RIOBIO CO (#1 target sequence: 5-CCAACGACCUGCAGCAAUAUU; #2 target sequence: 5-CUCAGAGGAUCCCGGAAAUUU).
Total RNAs were isolated with TRIzol reagent (Sigma) and corresponding cDNAs (complementary DNA) were obtained using the PrimeScript RT reagent kit. Real-time polymerase chain reactions were performed with SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA) and specific primers. The primers for the following human genes were used: STAT3 (Forward: CATCCTGAAGCTGACCCAGG; Reverse: TATTGCTGCAGGTCGTTGGT); p15 (Forward: AAGCTGAGCCCAGGTCTCCTA; Reverse: CCACCGTTGGCCGTAAACT); p21 (Forward: ACCATGTGGACCTGTCACTGT; Reverse: TTAGGGCTTCCTCTTGGAGAA); PAI-1 (Forward: GTGTTTCAGCAGGTGGCGC; Reverse: CCGGAACAGCCTGAAGAAGTG); β-actin (Forward: CAAAGTTCACAATGTGGCCGAGGA; Reverse: GGGACTTCCTGTAACAACGCATCT).
Immunofluorescence
HaCaT cells grown on coverslips were fixed with 4% formaldehyde for 15 min at 4 °C, permeabilized with 0.5% Triton X-100 for 15 min, followed by 5% bovine serum albumin blocking for 1 h. Cells were subsequently probed with indicated primary antibodies, and then with Alexa Fluor 488-conjugated or Alexa Fluor 546-secondary antibodies (Invitrogen). Fluoresence images were captured by a Zeiss LSM710 confocal microscope (Carl Zeiss, Jena, Germany).
DNA pull-down assay
DNA pull-down assay was carried out as previously described.49 HEK293T cells were lysated in buffer (10 mm HEPES (pH 7.5), 150 mm NaCl, 1 mm MgCl2, 0.5 mm EDTA, 0.5 mm DTT and 0.1% NP-40, 10% glycerol), then incubated with 0.5 μmol biotinylated SBE oligonucleotides together with 10 μg poly(dI-dC). DNA–protein complexes were then collected by precipitation on streptavidin beads (GE Healthcare, Waukesha, WI, USA) for 15 min, washed extensively with binding buffer and detected by western blotting.
Chromatin immunoprecipitation assay
HaCaT cells were treated with TGF-β (2 ng/ml) for 2 h. Cells were crosslinked with 1% formaldehyde for 10 min at 37 °C, and then quenched with 0.125 m glycine for 5 min at room temperature. Subsequently, cell pellets were suspended in sodium dodecyl sulfate lysis buffer and sonicated to generate DNA fragments. A Smad3 antibody was used to immunoprecipitated Smad3 and IgG was used as a control. Smad3-bound DNAs were determined by reverse transcriptase polymerase chain reaction. Chromatin immunoprecipitation primers used in the experiment are as follows: p15 Forward CTGCCTGGGGATGAATTTAAC; p15 Reverse GGTTTCACTGTGGAGACGTTG; PAI-1 Forward GCAGGACATCCGGGAGAGA; PAI-1 Reverse: CCAATAGCCTTGGCCTGAGA.
Immunoprecipitation and western blotting
Immunoprecipitation-coupled western blotting was performed as previously described.50 Briefly, cells were transfected with indicated plasmids and harvested 24-48 h after transfection. Co-IP was carried out with appropriate antibody and protein A Sepharose (GE Healthcare), followed by extensive washes. Precipitated proteins were eluted in sodium dodecyl sulfate loading buffer and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and detected in western blotting using appropriate antibodies.
In vitro protein binding assay
Recombinant GST fusion protein of Smad3 was prepared from E. coli strain BL21 (DE3). In vitro translation of STAT3 was carried out using Quick Coupled Transcription/Translation System (Promega). GST-Smad3 was incubated with STAT3 in the in vitro binding buffer (0.5% NP-40, 150 mm NaCl, 50 mm Tris-HCl, 5 mm EDTA) for 2 h. GST-Smad3 was retrieved using glutathione sepharose beads and examined by western blotting.
Cell cycle analysis
Cell cycle distribution was quantitatively evaluated by flow cytometry analysis. HaCaT cells were cultured in the absence or presence of TGF-β (2 ng/ml) for 2 days. Cells were detached and collected in a15- ml tube, and then fixed in 70% ice-cold ethanol for 24 h at −20 °C. Fixed cells were washed once with 1 × phosphate-buffered saline and stained with 1 mg/ml of propidium iodide at room temperature for 30 min. DNA content was then analyzed using a Flow Cytometer (Beckman Coulter, Brea, CA, USA).
Apoptosis assay
Hep3B cells were cultured in the absence or presence of TGF-β at indicated concentrations for 2 days. Cells were fixed as cell cycle analysis and then stained as per the manufacturer’s instruction (Annexin V/PI apoptosis kit; (Multisciences Biotech, Hangzhou, China). The stained cells were analyzed by a flow cytometer (Beckman Coulter).
References
Derynck R, Miyazono K . The TGF-β Family. Cold Spring Harbor Laboratory Press: New York, NY, USA, 2007, pp. 1–1114.
Flavell RA, Sanjabi S, Wrzesinski SH, Licona-Limón P . The polarization of immune cells in the tumour environment by TGFbeta. Nat Rev Immunol 2010; 10: 554–567.
Wu MY, Hill CS . TGF-beta superfamily signaling in embryonic development and homeostasis. Dev Cell 2009; 16: 329–343.
Massagué J . TGFβ in cancer. Cell 2008; 134: 215–230.
Ikushima H, Miyazono K . TGFβ signalling: a complex web in cancer progression. Nat Rev Cancer 2010; 10: 415–424.
Derynck R, Akhurst RJ, Balmain A . TGF-beta signaling in tumor suppression and cancer progression. Nat Genet 2001; 29: 117–129.
Drabsch Y, ten Dijke P . TGF-β signalling and its role in cancer progression and metastasis. Cancer Metastasis Rev 2012; 31: 553–568.
Deheuninck J, Luo K . Ski and SnoN, potent negative regulators of TGF-beta signaling. Cell Res 2009; 19: 47–57.
Wang D, Long J, Dai F, Liang M, Feng X-H, Lin X . BCL6 represses Smad signaling in transforming growth factor-beta resistance. Cell Res 2008; 68: 783–789.
Hirai H, Izutsu K, Kurokawa M, Mitani K . Oncogenic mechanisms of Evi-1 protein. Cancer Chemother Pharmacol 2001; 48: S35–S40.
Feng X-H, Liang Y-Y, Liang M, Zhai W, Lin X . Direct interaction of c-Myc with Smad2 and Smad3 to inhibit TGF-beta-mediated induction of the CDK inhibitor p15(Ink4B). Mol Cell 2002; 9: 133–143.
Massagué J, Gomis RR . The logic of TGFbeta signaling. FEBS Lett 2006; 580: 2811–2820.
Wrighton KH, Lin X, Feng X-H . Phospho-control of TGF-β superfamily signaling. Cell Res 2009; 19: 8–20.
Kretzschmar M, Doody J, Timokhina I, Massagué J . A mechanism of repression of TGFbeta/Smad signaling by oncogenic Ras. Genes Dev 1999; 13: 804–816.
Feng X-H, Derynck R . Specificity and versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev Biol 2005; 21: 659–693.
Massagué J, Seoane J, Wotton D . Smad transcription factors. Genes Dev Genes Dev 2005; 19: 2783–2810.
Labbé E, Silvestri C, Hoodless PA, Wrana JL, Attisano L . Smad2 and Smad3 positively and negatively regulate TGF beta-dependent transcription through the forkhead DNA-binding protein FAST2. Mol Cell 1998; 2: 109–120.
Chen X, Weisberg E, Fridmacher V, Watanabe M, Naco G, Whitman M . Smad4 and FAST-1 in the assembly of activin-responsive factor. Nature 1997; 389: 85–89.
Zhang Y, Feng XH, Derynck R . Smad3 and Smad4 cooperate with c-Jun/c-Fos to mediate TGF-beta-induced transcription. Nature 1998; 394: 909–913.
Yu H, Lee H, Herrmann A, Buettner R, Jove R . Revisiting STAT3 signalling in cancer: new and unexpected biological functions. Nat Rev Cancer 2014; 14: 736–746.
Ogata H, Chinen T, Yoshida T, Kinjyo I, Takaesu G, Shiraishi H et al. Loss of SOCS3 in the liver promotes fibrosis by enhancing STAT3-mediated TGF-beta1 production. Oncogene 2006; 25: 2520–2530.
Jenkins BJ, Grail D, Nheu T, Najdovska M, Wang B, Waring P et al. Hyperactivation of Stat3 in gp130 mutant mice promotes gastric hyperproliferation and desensitizes TGF-β signaling. Nat Med 2005; 11: 845–852.
Gunaje JJ, Bhat GJ . Distinct mechanisms of inhibition of interleukin-6-induced Stat3 signaling by TGF-beta and alpha-thrombin in CCL39 cells. Mol Cell Biol Res Commun 2000; 4: 151–157.
Walia B, Wang L, Merlin D, Sitaraman SV . TGF-beta down-regulates IL-6 signaling in intestinal epithelial cells: critical role of SMAD-2. FASEB J 2003; 17: 2130–2132.
Zauberman A, Lapter S, Zipori D . Smad proteins suppress CCAAT/enhancer-binding protein (C/EBP) beta- and STAT3-mediated transcriptional activation of the haptoglobin promoter. J Biol Chem 2001; 276: 24719–24725.
Dunfield LD, Nachtigal MW . Inhibition of the antiproliferative effect of TGFbeta by EGF in primary human ovarian cancer cells. Oncogene 2003; 22: 4745–4751.
Luwor RB, Baradaran B, Taylor LE, Iaria J, Nheu TV, Amiry N et al. Targeting Stat3 and Smad7 to restore TGF-β cytostatic regulation of tumor cells in vitro and in vivo. Oncogene 2012; 32: 2433–2441.
Feng X-H, Zhang Y, Wu R-Y, Derynck R . The tumor suppressor Smad4/DPC4 and transcriptional adaptor CBP/p300 are coactivators for Smad3 in TGF-beta. Genes Dev 1998; 12: 2153–2163.
Wierenga ATJ, Schuringa JJ, Eggen BJL, Kruijer W, Vellenga E . Downregulation of IL-6-induced STAT3 tyrosine phosphorylation by TGF-β1 is mediated by caspase-dependent and -independent processes. Leukemia 2002; 16: 675–682.
Campbell JD, Cook G, Robertson SE, Fraser A, Boyd KS, Gracie JA et al. Suppression of IL-2-induced T cell proliferation and phosphorylation of STAT3 and STAT5 by tumor-derived TGF beta is reversed by IL-15. J Immunol 2001; 167: 553–561.
Yamamoto T, Matsuda T, Muraguchi A, Miyazono K, Kawabata M . Cross-talk between IL-6 and TGF-beta signaling in hepatoma cells. FEBS Lett 2001; 492: 247–253.
Ghosh AK, Yuan W, Mori Y, Sj Chen, Varga J . Antagonistic regulation of type I collagen gene expression by interferon-gamma and transforming growth factor-beta. Integration at the level of p300/CBP transcriptional coactivators. J Biol Chem 2001; 276: 11041–11048.
Nakashima K, Yanagisawa M, Arakawa H, Kimura N, Hisatsune T, Kawabata M et al. Synergistic signaling in fetal brain by STAT3-Smad1 complex bridged by p300. Science 1999; 284: 479–482.
Hanahan D, Weinberg RA . Hallmarks of cancer: the next generation. Cell 2011; 144: 646–674.
Miyazono K, Suzuki H, Imamura T . Regulation of TGF-beta signaling and its roles in progression of tumors. Cancer Sci 2003; 94: 230–234.
Ungefroren H, Groth S, Sebens S, Lehnert H, Gieseler F, Fändrich F . Differential roles of Smad2 and Smad3 in the regulation of TGF-β1-mediated growth inhibition and cell migration in pancreatic ductal adenocarcinoma cells: control by Rac1. Mol Cancer 2011; 10: 67.
Zheng R, Xiong Q, Zuo B, Jiang S, Li F, Lei M et al. Using RNA interference to identify the different roles of SMAD2 and SMAD3 in NIH/3T3 fibroblast cells. Cell Biochem Funct 2008; 26: 548–556.
Huang D, Liu Y, Huang Y, Xie Y, Shen K, Zhang D et al. Mechanical compression upregulates MMP9 through SMAD3 but not SMAD2 modulation in hypertrophic scar fibroblasts. Connect Tissue Res 2014; 55: 391–396.
Kim SG, Kim H-A, Jong H-S, Park J-H, Kim NK, Hong SH et al. The endogenous ratio of Smad2 and Smad3 influences the cytostatic function of Smad3. Mol Biol Cell 2005; 16: 4672–4683.
Schmierer B, Schuster MK, Shkumatava A, Kuchler K . Activin a signaling induces Smad2, but not Smad3, requiring protein kinase a activity in granulosa cells from the avian ovary. J Biol Chem 2003; 278: 21197–21203.
Martinez GJ, Zhang Z, Chung Y, Reynolds JM, Lin X, Jetten AM et al. Smad3 differentially regulates the induction of regulatory and inflammatory T cell differentiation. J Biol Chem 2009; 284: 35283–35286.
Martinez GJ, Zhang Z, Reynolds JM, Tanaka S, Chung Y, Liu T et al. Smad2 positively regulates the generation of Th17 cells. J Biol Chem 2010; 285: 29039–29043.
Han S-U, Kim H-T, Seong DH, Kim Y-S, Park Y-S, Bang Y-J et al. Loss of the Smad3 expression increases susceptibility to tumorigenicity in human gastric cancer. Oncogene 2004; 23: 1333–1341.
Wolfraim LA, Fernandez TM, Mamura M, Fuller WL, Kumar R, Cole DE et al. Loss of Smad3 in acute T-cell lymphoblastic leukemia. N Engl J Med 2004; 351: 552–559.
Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006; 441: 235–238.
Okita K, Yamanaka S . Intracellular signaling pathways regulating pluripotency of embryonic stem cells. Curr Stem Cell Res Ther 2006; 1: 103–111.
Ying QL, Nichols J, Chambers I, Smith A . BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell 2003; 115: 281–292.
Lin X, Duan X, Liang Y-Y, Su Y, Wrighton KH, Long J et al. PPM1A functions as a Smad phosphatase to terminate TGFβ signaling. Cell 2006; 125: 915–928.
Dai F, Lin X, Chang C, Feng X-H . Nuclear export of Smad2 and Smad3 by RanBP3 facilitates termination of TGF-beta signaling. Dev Cell 2009; 16: 345–357.
Lin X, Liang Y-Y, Sun B, Liang M, Shi Y, Brunicardi FC et al. Smad6 recruits transcription corepressor CtBP to repress bone morphogenetic protein-induced transcription. Mol Cell Biol 2003; 23: 9081–9093.
Acknowledgements
We thank David Luskutoff for p800 (PAI-1)-luc, Peter ten Dijke for CAGA-luc, Bert Vogelstein for WWP1 (p21)-luc and SBE-luc and Xiao-Fan Wang for p15-luc. We are grateful to colleagues in our laboratories for helpful discussion and technical assistance. This research was partly supported by grants from MOST (2012CB966600) and NSFC (31571447; 31090360), NIH (R01GM63773, R01 AR053591, R01CA108454 and R01DK073932), Project 111, Project 985 and the Fundamental Research Funds for the Central Universities.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Wang, G., Yu, Y., Sun, C. et al. STAT3 selectively interacts with Smad3 to antagonize TGF-β. Oncogene 35, 4388–4398 (2016). https://doi.org/10.1038/onc.2015.446
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2015.446
- Springer Nature Limited
This article is cited by
-
Double-edged sword of JAK/STAT signaling pathway in viral infections: novel insights into virotherapy
Cell Communication and Signaling (2023)
-
Crosstalk between the JAK2 and TGF-β1 signaling pathways in scleroderma-related interstitial lung disease targeted by baricitinib
Advances in Rheumatology (2023)
-
Interleukin-22 Inhibits Apoptosis of Gingival Epithelial Cells Through TGF-β Signaling Pathway During Periodontitis
Inflammation (2023)
-
DYRK1A reinforces epithelial-mesenchymal transition and metastasis of hepatocellular carcinoma via cooperatively activating STAT3 and SMAD
Journal of Biomedical Science (2022)
-
The JAK/STAT signaling pathway: from bench to clinic
Signal Transduction and Targeted Therapy (2021)