

Abstract
Sedimentary rocks deposited across the Proterozoic–Phanerozoic transition record extreme climate fluctuations, a potential rise in atmospheric oxygen or re-organization of the seafloor redox landscape, and the initial diversification of animals1,2. It is widely assumed that the inferred redox change facilitated the observed trends in biodiversity. Establishing this palaeoenvironmental context, however, requires that changes in marine redox structure be tracked by means of geochemical proxies and translated into estimates of atmospheric oxygen. Iron-based proxies are among the most effective tools for tracking the redox chemistry of ancient oceans3,4. These proxies are inherently local, but have global implications when analysed collectively and statistically. Here we analyse about 4,700 iron-speciation measurements from shales 2,300 to 360 million years old. Our statistical analyses suggest that subsurface water masses in mid-Proterozoic oceans were predominantly anoxic and ferruginous (depleted in dissolved oxygen and iron-bearing), but with a tendency towards euxinia (sulfide-bearing) that is not observed in the Neoproterozoic era. Analyses further indicate that early animals did not experience appreciable benthic sulfide stress. Finally, unlike proxies based on redox-sensitive trace-metal abundances1,5,6, iron geochemical data do not show a statistically significant change in oxygen content through the Ediacaran and Cambrian periods, sharply constraining the magnitude of the end-Proterozoic oxygen increase. Indeed, this re-analysis of trace-metal data is consistent with oxygenation continuing well into the Palaeozoic era. Therefore, if changing redox conditions facilitated animal diversification, it did so through a limited rise in oxygen past critical functional and ecological thresholds, as is seen in modern oxygen minimum zone benthic animal communities7,8,9.
Similar content being viewed by others

Main
Proxies such as iron-speciation chemistry record the redox state of local water masses immediately above accumulating sediments. Decades of work on the behaviour of iron in marine sediments underpin the observation that enrichments in total (Fetot) and highly reactive (Fehr) iron phases track water-column redox conditions (Fehr refers to iron in pyrite plus iron that is reactive to sulfide on early diagenetic timescales)3,4. This robust calibration permits the differentiation between oxic and anoxic water columns, as well as whether anoxic waters were iron- or sulfide-bearing (this calculation is based on the proportion of highly reactive iron that has been converted to pyrite, Fepyr).
Early studies of iron speciation in Proterozoic shales supported the prediction10 of euxinia in subsurface waters of Mesoproterozoic oceans and further suggested deep-ocean oxygenation late in the Neoproterozoic era11,12. However, and perhaps not surprisingly, a more complex and heterogeneous pattern of Earth surface evolution emerged as additional studies increased temporal and spatial coverage. For example, marine strata deposited about 1,500 million years (Myr) ago from different localities show evidence of euxinic, ferruginous and oxic basins11,13,14. Similarly, Ediacaran deep-water sediments in Newfoundland indicate oxygenation at 580 Myr ago12, yet coeval deep-water deposits in the Canadian Cordillera show an increasing prevalence of anoxia15, or no change at all16. Such regional heterogeneity is expected given local controls on water-column redox, and highlights the fact that iron-speciation analyses of a single section or basin cannot be extrapolated to the global ocean.
Palaeontologists have long contended with an analogous problem: how to infer global diversity through time from fossil assemblages in local stratigraphic sections. The solution was to treat tabulated data within a global statistical framework17. Following this template, we have developed a data set of about 4,700 new and published iron-speciation measurements from fine-grained clastic rocks with which to test hypotheses of global redox change in Proterozoic/Palaeozoic oceans and the potential links to animal evolution. Importantly, local proxy data in a global framework can track both the mean and variance of palaeoenvironmental conditions through time. In addition to compiling data spanning the Great Oxidation Event (GOE, around 2,300 Myr ago) through the end-Devonian period, we provide 842 new analyses from Russia, northwestern Canada, Mongolia, Namibia, Svalbard, East Greenland and the western United States (Supplementary Table 2), focusing on Neoproterozoic and Cambrian strata.
Time-binned analysis of the entire data set begins with the most basic distinctions: geographic region and depositional environment (inner shelf, outer shelf, and basinal; following refs 11, 15). We note that the basinal environment does not represent true deep-ocean depths in a modern oceanographic sense, but rather the deepest environments represented by sediments deposited during maximum flooding; ‘basinal’ therefore refers to a recognizable and consistent sub-wave base environment that has been used to track deeper-water redox conditions through time (see Supplementary Information). To test for statistically significant differences, data were compared using analysis of variance (ANOVA) and Kruskal–Wallis tests depending on normality of the data. Post-hoc Tukey–Kramer tests (α = 0.05), pairwise Wilcoxon tests and Steel–Dwass tests were applied to explore significant differences between time bins (see Supplementary Information for binning rationale and sensitivity analyses).
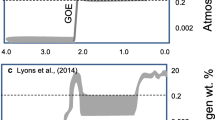
We first investigated the proportion of anoxic water columns through time. It has been hypothesized that a major oxygenation event occurred around the Proterozoic–Phanerozoic transition, oxygenating the world's deep oceans and facilitating Cambrian animal diversification. This idea has been bolstered by redox-sensitive trace-metal abundance data, which show evidence of increasing oxygen levels1,5,6, although the timing and magnitude remain poorly resolved1,2. Aggregated iron-speciation data provide an informative complement to global trace-metal data. Since the redox state of basinal water masses has traditionally been used as a proxy for the overall ocean–atmosphere system, and shallow-water samples are rare and heterogeneously distributed through time (Supplementary Table 1), this analysis includes only samples from outer shelf and basinal environments. The proportion of samples probably deposited beneath an anoxic water column (Fehr/Fetot > 0.38)3 was calculated for each region, and the mean and standard error were determined for each time bin. In contrast to trace-metal data, analysis of iron-speciation data does not show a significant change in the proportion of anoxic water columns from the Proterozoic into the early Palaeozoic (ANOVA F4,52 = 0.78, P = 0.54; Kruskal–Wallis χ2 = 3.30, P = 0.51) (Fig. 1a and Supplementary Table 4), which is consistent with qualitative observations in a previous compilation15.

The bins are: 2,300–1,000 Myr ago, 1,000–635 Myr ago, 635–542 Myr ago, 542–485 Myr ago and 485–360 Myr ago. The number of regions included in each bin is shown in grey text in parentheses. In b and c, the grey letters a, b or c represent the results of pairwise Wilcoxon tests. Bins joined by the same letter are not statistically significant (P > 0.05). a, The proportion of samples deposited beneath anoxic water columns (Fehr/Fetot > 0.38; refs 3, 19) from outer shelf and deep basin depositional environments. Each circle represents the average of regional proportions and the whiskers represent standard error. No bins are statistically different from one another (ANOVA P = 0.54; Kruskal–Wallis P = 0.51), and the proportion of oxic samples using a conservative threshold of Fehr/Fetot < 0.22 is also not significant (Supplementary Table 4). b, Proportion of samples deposited beneath ferruginous conditions from anoxic water columns (Fehr/Fetot > 0.38; Fepyr/Fehr < 0.70; ref. 3) from outer shelf and deep basin depositional environments. Each circle represents the average of regional proportions and whiskers represent standard error. c, Weight per cent iron in pyrite from samples deposited under oxic water columns from all depositional environments. Each circle represents the average of regional medians and whiskers represent standard error. The dashed line represents the modern oxic average from ref. 19. GOE, Great Oxidation Event.
Iron speciation more robustly identifies anoxia as opposed to oxic conditions, because Fehr enrichments can be muted during rapid deposition or in pervasively anoxic oceans where mass-balance requirements may not result in modern-like iron enrichment. Nonetheless, the proportion of oxic samples (using a conservative threshold of Fehr/Fetot < 0.22)3 was tested, and again no significant differences were found (Supplementary Table 4). This result raises a number of questions that we discuss below, ranging from diagnosing the nature of basinal anoxia to reconciling the seemingly divergent results between trace-metal geochemistry and our database analysis.
To assess the nature of anoxic waters through time we focused on samples from deeper-water environments with Fehr/Fetot > 0.38. The average proportion of ferruginous samples between 2,300 Myr ago and 1,000 Myr ago is 0.59 (the balance being euxinic), consistent with recent arguments that basinal waters through the middle of the Proterozoic were predominantly ferruginous3,13 (the effect of subdividing the Proterozoic using a shorter time bin of 1,600–1,000 Myr ago was also tested; Supplementary Table 4). In fact, anoxic waters throughout the Proterozoic and Palaeozoic are more likely to be ferruginous than euxinic. However, real differences exist between time bins (Kruskal–Wallis χ2 = 13.9, P = 0.008). Specifically, the late Palaeoproterozoic/Mesoproterozoic bin is more likely to capture euxinic conditions than the early Neoproterozoic, Ediacaran and Cambrian intervals, where the proportion of ferruginous samples approaches unity. The Ordovician–Devonian then marks a return to limited euxinia that is statistically distinct from the Neoproterozoic bins (Fig. 1b). Our analyses thus demonstrate that although a globally euxinic deep ocean10 did not exist, Mesoproterozoic oceans were statistically more prone to euxinia than those of the Neoproterozoic.
We further estimated sedimentary sulfide generation through Earth's history. This property cannot be measured directly, but can be evaluated indirectly, because sulfide generated within sediments will bond with reactive iron to form pyrite. Hence, reactive iron acts as an effective sulfide sink, meaning that sulfide accumulation in pore waters and advective fluxes into marine waters—the free sulfide that would influence local animal ecology—will only occur in settings where most, if not all, highly reactive iron has been pyritized18. Thus, for shale deposited in oxic environments, pyrite contents broadly serve as a metric for total sulfide generation, and only environments with Fepyr/Fehr > 0.70 could have contained high levels of pore-water sulfide.
Analyses of the weight per cent iron in pyrite from oxic sediments (Fig. 1c) show an inverted pattern from Fig. 1b, with higher pyrite contents in the late Palaeoproterozoic/Mesoproterozoic bin, very low contents in the Neoproterozoic and Cambrian, and higher contents again in the Ordovician–Devonian (Kruskal–Wallis χ2 = 25.44, P < 0.0001; Supplementary Table 4). The Neoproterozoic captures a minimum in pyrite preservation that is about five times smaller than in modern oxic samples19. Similar results are seen for the proportion of oxic samples with inferred high levels of pore-water sulfide (Supplementary Table 4). It is worth emphasizing that the outlier is the Neoproterozoic—whether in the water column or the sediments, far more sulfide was generated in Mesoproterozoic and Palaeozoic basins.
These results have important implications for the physiology and oxygen tolerance of early animals, which probably began to diverge about 800 Myr ago20. From observations in modern oxygen minimum zones21 and experiments on sponges22, it has been suggested that early animals would have tolerated the low-oxygen conditions believed to characterize the Neoproterozoic era. With oxygen partially removed as a handbrake on earliest animal evolution, other inhibitors such as ambient sulfide23 should be considered. Sulfide is a synergistic stressor in low-oxygen conditions because it binds to cytochrome oxidase and consequently inhibits aerobic respiration, lowering survival times under conditions of hypoxia24. But in contrast to some modern oxygen minimum zones where sulfide often reaches the sediment–water interface, Neoproterozoic animals would have experienced little, if any, benthic sulfide flux. In fact, out of 1,243 oxic Neoproterozoic samples analysed, only 14 (about 1.1%) show possible evidence of pore-water sulfide. This bolsters suggestions that while earlier Neoproterozoic oceans may have prohibited large, metabolically active and carnivorous animals with higher oxygen demands, they could have accommodated early animals with small and thin body plans21,22. Continued research on other proxies for the partial pressure of oxygen,  , will also help to place more precise constraints on early animal ecosystems25.
, will also help to place more precise constraints on early animal ecosystems25.
These results raise the question of whether observed trends reflect biases in the data set, as there are known caveats when interpreting iron-speciation data, most prominently including the effects of weathering and diagenesis3,4,15 (Supplementary Information). However, as long as the data are sufficiently numerous, and geological and analytical biases are randomly distributed with respect to time, these processes will not affect our results (see ref. 26 regarding analogous errors in palaeobiological data). The impact of random and systematic error can be tested with resampling and sensitivity analyses. Sensitivity analyses excluding possibly inappropriate samples and regions of low data coverage, and a further analysis using only Mesoproterozoic (1,600–1,000 Myr ago) samples for the oldest time bin, are consistent with results from the entire data set (Supplementary Table 4). Further, in synthetically re-sampled data sets, the Cambrian distribution of anoxic samples is indistinguishable from the Ediacaran distribution (Supplementary Fig. 2). To test whether inappropriate binning may contribute to the invariance in Fig. 1a, data from each region from the interval of 800–360 Myr ago were plotted individually with respect to time (Fig. 2). Although there is clear spatial heterogeneity (as in the modern ocean), there are no apparent ‘oxygenation events’, and a linear regression is not significant (P = 0.45; see also a local regression (LOESS) of geographically unbinned data, Supplementary Fig. 1).

Ages for different regions based on best geological estimates; Neoproterozoic samples from the same region were separated based on the global Sturtian and Marinoan glaciations, the Gaskiers glaciation or the mid-Ediacaran Shuram carbon isotope excursion and its equivalents, and the Ediacaran–Cambrian boundary. Grey bars represent 95% binomial confidence intervals.
It has been argued that trace metals in anoxic shales capture the spatial contraction of basinal anoxia across the Ediacaran–Cambrian transition1,5,6, probably driven by increasing atmospheric  . To evaluate the consistency between iron-speciation and trace-metal results, we re-analysed a well vetted sedimentary uranium data set6 using statistical methods similar to those employed in the iron analyses, although lower data density precludes a basin-normalized approach. The maximum ratios of metals to total organic carbon (TOC) are often taken as a guide to the metal inventory in ancient seawater; however, without a priori knowledge of basin restriction and secondary mineralization or local redistribution for each sample1,27, statistical approaches based on the entire population of data are appropriate.
. To evaluate the consistency between iron-speciation and trace-metal results, we re-analysed a well vetted sedimentary uranium data set6 using statistical methods similar to those employed in the iron analyses, although lower data density precludes a basin-normalized approach. The maximum ratios of metals to total organic carbon (TOC) are often taken as a guide to the metal inventory in ancient seawater; however, without a priori knowledge of basin restriction and secondary mineralization or local redistribution for each sample1,27, statistical approaches based on the entire population of data are appropriate.
When anoxic, organic-rich shales (TOC > 0.4%) are binned into Neoproterozoic, Cambrian–Silurian and Devonian–Permian domains, uranium/TOC significantly increases with younger age (Kruskal–Wallis χ2 = 75.53, P < 0.0001; all pairwise Wilcoxon tests P < 0.0001; see Supplementary Table 5). The Devonian–Permian time bin contains a much higher number of enriched outlier values relative to the Cambrian–Silurian time bin (Supplementary Fig. 3). Thus, while the uranium/TOC record does show a punctuated increase in oxygenation at the Ediacaran–Cambrian boundary, it is also consistent with iron geochemical data (Fig. 1a) that suggest that full oxygenation of the oceans did not occur until later.
The question then becomes the magnitude of oxygenation implied by the iron and trace-metal data sets. Recent models indicate that relatively subtle changes in seafloor anoxia and the proportion of the sea floor that was ferruginous rather than euxinic will lead to dramatic changes in seawater trace-metal inventories, and by inference, trace-metal enrichments in shales1,5,6,28. Trace-metal enrichments thus respond to the total size of anoxic sinks, whereas the binned iron data are tracking the percentage of sediments sampled in the stratigraphic record bathed by anoxic waters. As large changes in anoxic sink size can manifest as small shifts in the percentage of anoxic sea floor, we propose that trace-metal abundances and the binned iron-speciation records are complementary but have different thresholds; that is, binned iron data require a larger change in global oxygen to record a statistically significant (see above) signal.
Although absolute values of  in the geological record are notoriously difficult to track, the iron-speciation database results constrain the magnitude of the latest Proterozoic
in the geological record are notoriously difficult to track, the iron-speciation database results constrain the magnitude of the latest Proterozoic  increase indicated by trace-metal compilations. Canfield10 earlier posited that at atmospheric
increase indicated by trace-metal compilations. Canfield10 earlier posited that at atmospheric  PAL (Present Atmospheric Level), deeper water masses tend towards anoxia, albeit dependent upon phosphorus fluxes. Although this was intended to constrain oxygen levels before Ediacaran oxygenation, it also provides an upper bound on Cambrian
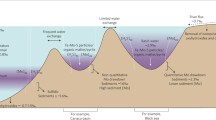
PAL (Present Atmospheric Level), deeper water masses tend towards anoxia, albeit dependent upon phosphorus fluxes. Although this was intended to constrain oxygen levels before Ediacaran oxygenation, it also provides an upper bound on Cambrian  , given the lack of statistical change through time. The distribution of animals in modern oceans8,29 suggests that the Cambrian metazoans recorded by fossils required oxygen levels above about 10% PAL, but not much more than that, given that equally large, mobile and skeletonized animals live at and even below this level in the modern ocean8,29. The combined constraints from iron-speciation and palaeontological data are therefore consistent with molybdenum isotope data28, global sedimentary sulfate reduction rates30, uranium/TOC6 (Supplementary Table 5) and some models of atmospheric oxygen through time31. All offer evidence that oxygenation of the ocean–atmosphere system to essentially modern levels and a persistently oxygenated deep ocean is in large part a post-Cambrian phenomenon, as has been separately hypothesized for black shale distribution32. Overall, these analyses imply a modest increase in oxygen during the Ediacaran and Cambrian (Fig. 3).
, given the lack of statistical change through time. The distribution of animals in modern oceans8,29 suggests that the Cambrian metazoans recorded by fossils required oxygen levels above about 10% PAL, but not much more than that, given that equally large, mobile and skeletonized animals live at and even below this level in the modern ocean8,29. The combined constraints from iron-speciation and palaeontological data are therefore consistent with molybdenum isotope data28, global sedimentary sulfate reduction rates30, uranium/TOC6 (Supplementary Table 5) and some models of atmospheric oxygen through time31. All offer evidence that oxygenation of the ocean–atmosphere system to essentially modern levels and a persistently oxygenated deep ocean is in large part a post-Cambrian phenomenon, as has been separately hypothesized for black shale distribution32. Overall, these analyses imply a modest increase in oxygen during the Ediacaran and Cambrian (Fig. 3).

Data are based on the combined absence of a statistically significant oxygenation event in iron-speciation data and the presence of an oxygenation event in redox-sensitive trace-metal inventories. Oxygen constraints include: (1) persistently anoxic subsurface waters requiring less than 40% PAL10 (iron-speciation data for the Ordovician–Devonian is not statistically different from that of previous time bins, but data are sparse and may be subject to sampling biases—see Supplementary Information); (2) a minimum oxygen level of ∼0.5–1% PAL, required for the appearance of mass-dependent sulfur isotope fractionation, red beds, and the earliest animals1,21, although oxygen levels before ∼810 Myr ago may have been lower25; (3) oxygen levels exceeding 10% PAL8,29, required by the Cambrian biota; and (4) oxygen levels must have exceeded 70% PAL in the latest Silurian, as deduced from the presence of fires33. Within these constraints, oxygenation could have followed many different paths, but full oxygenation of the ocean–atmosphere system is a Palaeozoic phenomenon. Ediac., Ediacaran; Cryo., Cryogenian; C., Cambrian; O., Ordovician; S., Silurian; Dev., Devonian.
This evolving picture of Earth's redox state would seem to diminish the impact of oxygen as a causal factor in Cambrian animal radiation. Observations from modern oxygen minimum zones, however, suggest that a small increase in  could still be a critical environmental trigger owing to nonlinear threshold effects at very low oxygen levels. Many important ecological responses for macrofaunal organisms, including feeding efficiency9, species-level diversity8, and carnivore abundance and species richness7 exhibit threshold changes in the range of 5–20 µM oxygen, or ∼2–7% of modern surface ocean oxygen concentrations—results that are strikingly similar to the changes accommodated by this analysis. Thus, a relatively small increase in
could still be a critical environmental trigger owing to nonlinear threshold effects at very low oxygen levels. Many important ecological responses for macrofaunal organisms, including feeding efficiency9, species-level diversity8, and carnivore abundance and species richness7 exhibit threshold changes in the range of 5–20 µM oxygen, or ∼2–7% of modern surface ocean oxygen concentrations—results that are strikingly similar to the changes accommodated by this analysis. Thus, a relatively small increase in  could reasonably have moved animals past critical ecological thresholds, especially with respect to carnivory7, which might have driven Cambrian diversification. It remains possible, though, that sufficient oxygen for large, muscular carnivores existed before the Cambrian (Fig. 3). The critical question is whether oxygen availability before the Ediacaran–Cambrian transition was in the ∼1–5% PAL range (at which modern animal ecology is severely limited), or higher.
could reasonably have moved animals past critical ecological thresholds, especially with respect to carnivory7, which might have driven Cambrian diversification. It remains possible, though, that sufficient oxygen for large, muscular carnivores existed before the Cambrian (Fig. 3). The critical question is whether oxygen availability before the Ediacaran–Cambrian transition was in the ∼1–5% PAL range (at which modern animal ecology is severely limited), or higher.
Coupled with other geochemical data, our global database of iron-speciation measurements provides an increasingly resolved and quantitative picture of redox evolution in Proterozoic and Palaeozoic oceans. These data point to proportionally higher basinal euxinia in Mesoproterozoic and younger Palaeozoic basins, with sediment and water-column sulfide generation reaching a minimum in the Neoproterozoic oceans. Ediacaran oxygenation was relatively modest, but may have been sufficient to remove environmental barriers to Cambrian animal evolution. Future sedimentary geochemical sampling of both iron and redox-sensitive trace-metal data will increase temporal resolution and the power of inference tests, with statistical analysis in a basin-normalized context providing more robust hypotheses of deep-time global change.
References
Lyons, T. W., Reinhard, C. T. & Planavsky, N. J. The rise of oxygen in Earth's early ocean and atmosphere. Nature 506, 307–315 (2014)
Lenton, T. M., Boyle, R. A., Poulton, S. W., Shields-Zhou, G. A. & Butterfield, N. J. Co-evolution of eukaryotes and ocean oxygenation in the Neoproterozoic era. Nature Geosci. 7, 257–265 (2014)
Poulton, S. W. & Canfield, D. E. Ferruginous conditions: a dominant feature of the ocean through Earth's history. Elements 7, 107–112 (2011)
Lyons, T. W. & Severmann, S. A critical look at iron paleoredox proxies: new insights from modern euxinic marine basins. Geochim. Cosmochim. Acta 70, 5698–5722 (2006)
Reinhard, C. T. et al. Proterozoic ocean redox and biogeochemical stasis. Proc. Natl Acad. Sci. USA 110, 5357–5362 (2013)
Partin, C. A. et al. Large-scale fluctuations in Precambrian atmospheric and oceanic oxygen levels from the record of U in shales. Earth Planet. Sci. Lett. 369–370, 284–293 (2013)
Sperling, E. A. et al. Oxygen, ecology, and the Cambrian radiation of animals. Proc. Natl Acad. Sci. USA 110, 13446–13451 (2013)
Levin, L. A. Oxygen Minimum Zone benthos: adaptation and community response to hypoxia. Oceanogr. Mar. Biol. Annu. Rev. 41, 1–45 (2003)
Woulds, C. et al. Oxygen as a control on seafloor biological communities and their roles in sedimentary carbon cycling. Limnol. Oceanogr. 52, 1698–1709 (2007)
Canfield, D. E. A new model for Proterozoic ocean chemistry. Nature 396, 450–453 (1998)
Shen, Y., Knoll, A. H. & Walter, M. R. Evidence for low sulphate and anoxia in a mid-Proterozoic marine basin. Nature 423, 632–635 (2003)
Canfield, D. E., Poulton, S. W. & Narbonne, G. M. Late-Neoproterozoic deep-ocean oxygenation and the rise of animal life. Science 315, 92–95 (2007)
Planavsky, N. J. et al. Widespread iron-rich conditions in the mid-Proterozoic ocean. Nature 477, 448–451 (2011)
Sperling, E. A. et al. Redox heterogeneity of subsurface waters in the Mesoproterozoic ocean. Geobiology 12, 373–386 (2014)
Canfield, D. E. et al. Ferruginous conditions dominated later Neoproterozoic deep-water chemistry. Science 321, 949–952 (2008)
Johnston, D. T. et al. Searching for an oxygenation event in the fossiliferous Ediacaran of northwestern Canada. Chem. Geol. 362, 273–286 (2013)
Alroy, J. et al. Phanerozoic trends in the global diversity of marine invertebrates. Science 321, 97–100 (2008)
Canfield, D. E. Reactive iron in marine sediments. Geochim. Cosmochim. Acta 53, 619–632 (1989)
Raiswell, R. & Canfield, D. E. Sources of iron for pyrite formation in marine sediments. Am. J. Sci. 298, 219–245 (1998)
Erwin, D. H. et al. The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science 334, 1091–1097 (2011)
Sperling, E. A., Halverson, G. P., Knoll, A. H., Macdonald, F. A. & Johnston, D. T. A basin redox transect at the dawn of animal life. Earth Planet. Sci. Lett. 371–372, 143–155 (2013)
Mills, D. B. et al. Oxygen requirements of the earliest animals. Proc. Natl Acad. Sci. USA 111, 4168–4172 (2014)
Martin, W. et al. Early cell evolution, eukaryotes, anoxia, sulfide, oxygen, fungi first (?), and a tree of genomes revisited. IUBMB Life 55, 193–204 (2003)
Vaquer-Sunyer, R. & Duarte, C. M. Sulfide exposure accelerates hypoxia-driven mortality. Limnol. Oceanogr. 55, 1075–1082 (2010)
Planavsky, N. J. et al. Low Mid-Proterozoic atmospheric oxygen levels and the delayed rise of animals. Science 346, 635–638 (2014)
Sepkoski, J. J., Jr Ten years in the library: new data confirm paleontological patterns. Paleobiology 19, 43–51 (1993)
Algeo, T. J. & Rowe, H. Paleoceanographic applications of trace-metal concentration data. Chem. Geol. 324–325, 6–18 (2011)
Dahl, T. W. et al. Devonian rise in atmospheric oxygen correlated to the radiation of terrestrial plants and large predatory fish. Proc. Natl Acad. Sci. USA 107, 17911–17915 (2010)
Rhoads, D. C. & Morse, J. W. Evolutionary and ecologic significance of oxygen-deficient marine basins. Lethaia 4, 413–428 (1971)
Leavitt, W. D., Halevy, I., Bradley, A. S. & Johnston, D. T. Influence of sulfate reduction rates on the Phanerozoic sulfur isotope record. Proc. Natl Acad. Sci. USA 110, 11244–11249 (2013)
Bergman, N. M., Lenton, T. M. & Watson, A. J. COPSE: a new model of biogeochemical cycling over Phanerozoic time. Am. J. Sci. 304, 397–437 (2004)
Berry, W. B. & Wilde, P. Progressive ventilation of the oceans; an explanation for the distribution of the lower Paleozoic black shales. Am. J. Sci. 278, 257–275 (1978)
Belcher, C. M. & McElwain, J. C. Limits for combustion in low O2 redefine paleoatmospheric predictions for the Mesozoic. Science 321, 1197–1200 (2008)
Acknowledgements
We thank D. Cole, A. Masterson, E. Beirne, G. Resendiz, A. Miller and W. Mai for laboratory assistance, E. Smith, T. Petach, M. Laflamme, S. Darroch, P. Myrow, J. Strauss, C. Carbone and G. Narbonne for field assistance, L. Levin, P. Girguis, C. Frieder, L. Duncan, C. Partin, S. Darroch and U. Farrell for discussions, and E. Hammarlund, C. Scott, R. Raiswell, G. Gilleaudeau, S. Sahoo, Y. Shen, J. Creveling, T. Dahl, C. Partin and D. Stolper for providing raw data from published studies. E.A.S. is funded by Agouron Geobiology and NAI Postdoctoral Fellowships and by the NSF-EAR 1324095 grant to L. Levin. This work is supported by the NSF-EAR 1324095 grant to D.T.J. and by the NASA Astrobiology Institute (A.H.K. and F.A.M.).
Author information
Authors and Affiliations
Contributions
E.A.S. and D.T.J. conceived the project, E.A.S., A.S.M., B.C.G., G.P.H., F.A.M. and D.T.J. collected samples, E.A.S., A.S.M., B.C.G. and M.K. completed geochemical measurements, E.A.S. and C.J.W. compiled data and analysed the global data set, and E.A.S. wrote the paper with input from all co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
This file contains Supplementary Text and Data, Supplementary Figures 1-4, Supplementary Tables 1-7 and Supplementary References. (PDF 1592 kb)
Supplementary Data
This file contains Supplementary Dataset 1. (XLSX 638 kb)
Rights and permissions
About this article

Cite this article
Sperling, E., Wolock, C., Morgan, A. et al. Statistical analysis of iron geochemical data suggests limited late Proterozoic oxygenation. Nature 523, 451–454 (2015). https://doi.org/10.1038/nature14589
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14589
- Springer Nature Limited
This article is cited by
-
Late Paleozoic oxygenation of marine environments supported by dolomite U-Pb dating
Nature Communications (2024)
-
Rates of seafloor and continental weathering govern Phanerozoic marine phosphate levels
Nature Geoscience (2023)
-
Spatial pattern of marine oxygenation set by tectonic and ecological drivers over the Phanerozoic
Nature Geoscience (2023)
-
Evidence for high-frequency oxygenation of Ediacaran shelf seafloor during early evolution of complex life
Communications Earth & Environment (2023)
-
Recurrent photic zone euxinia limited ocean oxygenation and animal evolution during the Ediacaran
Nature Communications (2023)