
Abstract
Background/Objective:
Body mass index (BMI) is a surrogate measure of adiposity but does not distinguish fat from lean or bone mass. The genetic determinants of BMI are thought to predominantly influence adiposity but this has not been confirmed. Here we characterise the association between BMI-related genetic variants and body composition in adults.
Subjects/Methods:
Among 9667 adults aged 29–64 years from the Fenland study, a genetic risk score for BMI (BMI-GRS) was calculated for each individual as the weighted sum of BMI-increasing alleles across 96 reported BMI-related variants. Associations between the BMI-GRS and body composition, estimated by dual-energy X-ray absorptiometry (DXA) scans, were examined using age-adjusted linear regression models, separately by sex.
Results:
The BMI-GRS was positively associated with all fat, lean and bone variables. Across body regions, associations of the greatest magnitude were observed for adiposity variables, for example, for each s.d. increase in BMI-GRS predicted BMI, we observed a 0.90 s.d. (95% confidence interval (CI): 0.71, 1.09) increase in total fat mass for men (P=3.75 × 10−21) and a 0.96 s.d. (95% CI: 0.77, 1.16) increase for women (P=6.12 × 10−22). Associations of intermediate magnitude were observed with lean variables, for example, total lean mass: men: 0.68 s.d. (95% CI: 0.49, 0.86; P=1.91 × 10−12); women: 0.85 s.d. (95% CI: 0.65, 1.04; P=2.66 × 10−17) and of a lower magnitude with bone variables, for example, total bone mass: men: 0.39 s.d. (95% CI: 0.20, 0.58; P=5.69 × 10−5); women: 0.45 s.d. (95% CI: 0.26, 0.65; P=3.96 × 106). Nominally significant associations with BMI were observed for 28 single-nucleotide polymorphisms. All 28 were positively associated with fat mass and 13 showed adipose-specific effects.
Conclusions:
In adults, genetic susceptibility to elevated BMI influences adiposity more than lean or bone mass. This mirrors the association between BMI and body composition. The BMI-GRS can be used to model the effects of measured BMI and adiposity on health and other outcomes.
Similar content being viewed by others

Introduction
Obesity describes a state of excess adiposity that confers risks to health.1 It is most often identified using body mass index (BMI; weight(kg)/height(m2)), a non-invasive proxy measure of adiposity, according to which 67% of women and 57% of men in the UK are classified as either overweight (BMI⩾25 and <30 kg m−2) or obese (BMI⩾30 kg m−2).2 Together overweight and obesity represent a major, global public health concern with severe ramifications for individuals, communities, health care services and the economy.
The primary purpose of BMI is to estimate metabolic health by approximating fat mass. Across diverse settings, BMI shows strong, linear associations with body fat in largely sedentary populations.3, 4, 5 However, there are limitations to the ability of BMI to characterise health status. In addition to fat mass, BMI is also positively influenced by both lean tissue and bone mass. This can lead to inaccuracies in estimating adiposity.4, 6, 7 BMI is also uninformative with regard to body fat distribution. Both total adiposity and fat distribution have associations with health outcomes that are independent of BMI.8, 9, 10
There is a strong heritable component to BMI. Heritability estimates from twin studies range from 47 to 90%.11 The most recent meta-analysis of genome-wide association studies identified 97 genetic variants that exhibit independent, genome-wide significant (P<5 × 10−8) associations with BMI12 in adulthood.
It is assumed that, in combination, these variants have the same patterns of association as measured BMI with fat, lean and bone mass. Thus, they are predominantly associated with variation in fat mass and do not reflect body fat distribution. The implication is that they can be used to model the effect of both measured BMI and total adiposity on health outcomes. However, this assumption has not been tested.
To assess the utility of known BMI-related variants for modelling the effects of BMI and adiposity on health, it is important to determine the components of body composition and patterns of fat distribution most associated with these loci.
Previous studies of the relationship between the combined effect of known BMI-related loci and body composition in adults have investigated a maximum of 32 variants13 and tested a limited range of body composition measures.13, 14, 15, 16 No previous study has investigated the association of all 96 biallelic reported BMI-related variants with fat, lean and bone mass across body regions.
We characterised the association between 96 BMI-related variants, summarised in a polygenic BMI genetic risk score (BMI-GRS), and fat, lean and bone mass across body regions in a large, population-based adult study.
Subjects and Methods
Subjects and study design
The Fenland study is a population-based cohort study of volunteers recruited from participating General Practices in Cambridge, Ely, Wisbech and the surrounding Cambridgeshire region between 2004 and 2014.17 Eligible individuals were adults residing in Cambridgeshire and registered at a collaborating General Practice at the time of recruitment. Exclusion criteria were: clinically diagnosed diabetes mellitus, inability to walk unaided, terminal illness (life expectancy of ⩽1 year at the time of recruitment), clinically diagnosed psychotic disorder, pregnancy or lactation. The study population of the present analysis comprised 9667 individuals (53.2% female; 92.3% White British ancestry) aged 29–64 years.
Participants attended one of three MRC Epidemiology Unit testing centres for data collection. Total and regional body composition was measured using DXA scans.
Written informed consent was attained from all participants and the study was approved by the Cambridge Local Research Ethics Committee.
Genotyping and the BMI genetic risk score (BMI-GRS)
DNA was genotyped using the Affymetrix UK Biobank Axiom array (Santa Clara, CA, USA. Missing genotypes and those not directly measured were imputed via IMPUTE version218 based on the 1000 Genomes Project haplotype reference.19 All required single-nucleotide polymorphisms (SNPs) within this subpopulation could be imputed in this manner with sufficient accuracy (imputation information value >0.4).
The 97 BMI-related SNPs reported by Locke et al.12 were considered for inclusion in the BMI-GRS. One SNP, rs2033529 (nearest gene: TDRG1), was multi-allelic and could not be incorporated. The remaining 96 SNPs were included.
A weighted BMI-GRS was calculated for each participant using a previously reported method.13 At each locus, participants were assigned a value of 0, 1 or 2, indicating the number of BMI-increasing variants. This value was multiplied by the ‘European only sex-combined’ effect estimate for the BMI-increasing allele reported by Locke et al.12 The products across all 96 SNPs were then summed for each participant.
The European sex-combined SNP estimates were chosen because, owing to the demography of the Fenland population, the participant group of the present analysis was highly ethnically homogenous. Of those who stated their ethnicity, 98.9% identified as white and 92.3% as white British (Supplementary Table 2). Thus the European estimates reflect the demography of the study sample most closely. In the absence of sex-specific effects for the vast majority of loci (only two show evidence of heterogeneity12), the sex-combined effect estimates are likely to be more accurate as they are derived from a larger sample size than the sex-specific estimates.
The effect estimates from Locke et al. were generated using inverse normally transferred residual measurements. These cannot be translated to BMI units. To aid interpretation, the BMI-GRS for each participant was multiplied by the s.d. increase in BMI per unit increase in the BMI-GRS, adjusted for age. Scaling was performed separately in each sex as the main analysis was sex-stratified. After this adjustment, one BMI-GRS unit corresponds to 1 s.d. of BMI-GRS predicted BMI in this sample.
Overall, the formula for the BMI-GRS is as follows:

where GRSj is the BMI-GRS for individual j, 96 is the number of SNPs included in the score, Sij is the number of BMI-increasing alleles at SNP i for individual j and Wi is the effect estimate of SNP i on inverse normally transformed BMI, as reported by Locke et al.12 βGRS is the regression coefficient of the weighted BMI-GRS on BMI z-score, adjusted for age, in the current Fenland study (βGRS = 0.94 in men and 0.83 in women). This last parameter was included to align all effect estimates to a +1 s.d. change in BMI in this population.
Anthropometry and body composition
Anthropometric and body composition measures were collected by trained observers following an established protocol.17 Volunteers were barefoot and wore light clothing. Weight was measured to the nearest 100 g using a calibrated scale (TANITA model BC-418 MA; Tanita, Tokyo, Japan) and height was measured to the nearest 0.1 cm using a calibrated wall-mounted stadiometer (SECA 240; Seca, Birmingham, UK). Waist and hip circumferences were measured to the nearest 0.1 cm using a non-stretchable fibre-glass insertion tape (D-loop tape; Chasmors Ltd, London, UK). Waist and hip measurements were taken twice. If these measures differed by more than 3 cm, a third measurement was taken. The mean of the two or three measurements was used in the present analysis.
Full body DXA scans (GE Lunar Prodigy Advanced, GE Medical Systems, Hartfield, UK) were used to derive fat, lean and bone mass measurements across body regions. Scans were performed for all consenting participants weighing ⩽140 kg. Beyond this threshold, the scanner was considered to be insufficiently precise to warrant performing the scan.
Fat, lean and bone mass measures in the total body, trunk, android, gynoid and leg regions were generated and appendicular lean mass (lean mass in the legs + lean mass in the arms) was calculated. The DXA software (enCORE software (version 14.10.022 to 16, GE Medical Systems) generated estimates of visceral adipose tissue within the android region (VAT) for individuals whose girth allowed them to fit within the scanning area and who had >1 g of VAT. From the VAT measures, values for subcutaneous adipose tissue within the android region (SAT) were generated (SAT = android fat mass-VAT) and VAT/SAT ratio was calculated.
The following variables were included in the present analysis: BMI (kg m−2), weight (kg), WC (cm), HC (cm), waist-to-hip ratio (WHR; WC/HC), height (cm) and body fat percentage (BF%). Fat, lean and bone mass measurements in the total body, trunk, android and gynoid regions were included alongside bone and fat mass in the legs and appendicular lean mass. Finally, SAT (kg), VAT (kg) and VAT/SAT ratio measures were included.
Statistical analysis
The pattern of missing data was examined for body composition variables missing data for ⩾1% of study participants. Individuals with missing values were compared with the rest of the cohort for sex, age, ethnicity and BMI using logistic regression, χ2-tests or Fisher’s exact tests.
The BMI-GRS was tested for cross-sectional associations with body composition variables in age-adjusted least-squares linear regression models. Body composition variables were analysed separately in each sex. The distribution of the residuals was checked to ensure that the body composition variables did not need to be transformed. A Bonferroni-corrected P-value of P<1.04 × 10−3, corrected for 48 tests (24 tests in each sex), was used to assess significance.
Just two out of the 96 BMI-related SNPs show evidence of heterogeneity between the sexes for BMI. However, SNPs associated with some included outcome measures (for example, WHR,20 VAT,21 SAT21 and BF%) show sex-specific effects. For example, IRS1 shows a stronger effect on BF% in men than in women.22, 23 It is possible that BMI-related variants exhibit heterogeneous effects with regard to traits of interest to this analysis. As such, our analysis is sex-stratified.
Two sensitivity analyses were conducted. The first repeated the main analysis using sex-specific GRSs generated using sex-specific effect estimates from Locke et al.12 The second repeated the main analyses amongst white participants only.
To facilitate comparison of variables measured in different units, body composition variables were standardised to z-scores with mean=0 and s.d.=1. The results of the regressions are displayed alongside the R2 values from the age-adjusted regressions of BMI z-score on the body composition variable z-scores in each sex.
In a secondary analysis, we investigated the relationship between individual BMI-related SNPs and body composition. For this analysis, we only included SNPs that demonstrated nominally significant associations with BMI in this cohort (P<0.05). Our sample size is only a fraction of that of the genome-wide association studies meta-analysis in which the SNPs were identified and is not powered to detect associations between all 96 SNPs and BMI individually. As this investigation explores the relationship between the genetic determinants of BMI and body composition, it is logical only to test the subset of SNPs associated with BMI in this cohort for associations with body composition measures.
SNPs that were nominally associated with BMI in this cohort were considered in linear regression models to characterise their age and sex-adjusted relationships with total body fat, lean and bone mass. The body composition measures were standardised to z-scores and, in the absence of significant heterogeneity in the associations between the BMI-GRS and body composition, or an interaction between sex and the BMI-GRS, both sexes were combined to maximise power. The results were used to construct a heat map colour-coding the z-statistic for the association of each SNP with each body composition variable. To avoid spurious precision, z-statistic values between −0.5 and 0.5 were displayed as neutral.
All analyses were performed in Stata version 14.1 (TX, USA). The heat map was produced using R version 3.2 (Vienna, Austria).
Results
Characteristics of study participants
The characteristics of the 9667 participants are summarised in Table 1. All anthropometric and body composition variables exhibited significant sexual dimorphism. Reflecting UK population norms, the majority of participants were either overweight or obese (3181 men (70.3%) and 2778 women (54.0%)). The majority of participants (9137; 98.9%) were of white ethnicity. The single largest ethnic group were those reporting white British ethnicity (8526; 92.3%). Detailed ethnic information is included in Supplementary Table 2.
VAT, SAT and VAT/SAT ratio were the only variables with ⩾1% missing data (221 missing; 2.29% of the cohort). The majority (n=207) were missing due to low VAT mass (<1g) as the DXA software could not estimate values for these people. The remainder of participants with missing values (n=14) were too large for estimates to be made. Compared with the rest of the cohort, individuals with missing data exhibited higher mean age (48.6 versus 44.7 years; P<0.01), were more likely to be female (97.7% versus 52.2%; P<0.01) and had a lower median BMI (21.5 versus 26.3 kg m−2; P<0.01).
Associations with the BMI-GRS
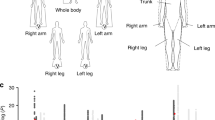
Associations between the BMI-GRS and the body composition variables were universally positive (Figure 1 and Table 2). Out of the 48 associations tested, 43 were statistically significant at the Bonferroni-corrected P-value threshold for 48 tests (P<1.04 × 10−3). Only VAT/SAT ratio (men: β=0.06 s.d. (95% CI: −0.12, 0.24); P=0.54; women: 0.28 s.d. (95% CI: 0.09, 0.47); P=4.29 × 10−3), android bone mass in men (β= 0.29 s.d. (95% CI: 0.10, 0.48); P=2.74 × 10−3) and height (men: β= −0.02 s.d. (95% CI: −0.21, 0.17); P=0.85; women: 0.10 s.d. (95% CI: −0.10, 0.30); P=0.31), did not reach statistical significance at this threshold.

Associations between the BMI-GRS and body composition (columns) and measured BMI and body composition (squares). A shows the results for men and B shows the results for women. Columns display the standardised beta estimates from linear regression of the BMI-GRS on body composition z-scores, adjusted for age, with 95% confidence intervals. They represent the s.d. change in each body composition variable per s.d. increase in BMI-GRS predicted BMI. The squares display the variance in each body composition variable explained by measured BMI.
Associations with the BMI-GRS were of the greatest magnitude for adiposity variables, intermediate for lean tissue variables and lowest for bone variables (Figure 1). This pattern held for the total body as well as within all measured body regions. Alongside the beta estimates, Figure 1 displays the variance in each of the body composition variables explained by measured BMI (R2 values). The R2 values largely mirror the association of the BMI-GRS with body composition.
The beta estimates for fat mass variables were comparable across body regions. For men, the highest estimate was for SAT mass and the lowest was for VAT mass. For women, the highest estimate was for total body fat and the lowest was for VAT mass. In both sexes, the beta estimates were greater for SAT than for VAT mass. For each s.d. increase in BMI-GRS predicted BMI, we saw a 0.98 s.d. (95% CI: 0.79, 1.16) increase in SAT mass for men and a 0.92 s.d. (95% CI: 0.72, 1.12) increase for women. The corresponding VAT estimates were 0.62 s.d. (95% CI: 0.44, 0.80) and 0.70 s.d. (95% CI: 0.51, 0.90).
The BMI-GRS showed significant positive associations with WHR in both sexes: β=0.61 s.d. (95% CI: 0.43, 0.79), P=5.33 × 10−11 for men and 0.41 s.d. (95% CI: 0.21, 0.60), P=4.24 × 10−5 for women. Directionally consistent but not statistically significant associations were observed for VAT/SAT ratio in both sexes: β=0.06 s.d. (95% CI: −0.12, 0.24), P=0.54 for men and 0.28 s.d. (95% CI: 0.09, 0.47), P=4.29 × 10−3 for women.
Associations between the BMI-GRS and body composition variables followed similar patterns in both sexes and no statistical differences between the effect estimates for men and women were found for any of the variables. Sensitivity analyses of sex-specific BMI-GRSs (Supplementary Tables 5 and 6) and of white only participants (Supplementary Tables 3 and 4) did not alter the conclusions.
Associations with individual SNPs
Nominally significant associations with BMI were observed for 28 SNPs in this cohort (P<0.05). All 28 (100%) showed positive associations with fat mass, of which 22 were nominally significant. Positive associations with lean mass were observed for 24 SNPs (85.7%), of which 11 were nominally significant. Finally, 20 SNPs (71.4%) showed positive associations with bone mass, of which eight were nominally significant. For all 28 SNPs, the greatest magnitude of association was with fat mass (see Supplementary Table 1 for beta estimates).
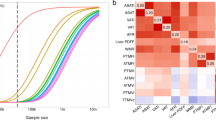
On the heat map of individual SNP to body composition associations (Figure 2), the primary clustering of body composition variables on the X axis separated fat from lean and bone mass. The primary clustering of the SNPs on the Y axis separated a group of 15 SNPs that are associated with a global increase in all three body composition measures, from a group of 13 SNPs with apparent adipose-specific effects. MTCH2 is notable for exhibiting a nominally significant, negative association with bone mass: β=−0.78 (95% CI: −1.52, −0.04); P=0.04 (Supplementary Table 1).

Heat map showing the 28 SNPs that exhibited a nominally significant association with BMI in this cohort, clustered by their associations with fat, lean and bone mass The values and colour-coding indicates the z-statistic (beta/standard error) from the age and sex-adjusted linear regression of each SNP with the standardised body composition variables (mean=0; s.d.=1).
Discussion
In this study, we characterised the associations between a genetic risk score for BMI (BMI-GRS) comprised of 96 SNPs and body composition in a population-based sample of 9667 adults aged 29–64 years of predominantly white British ancestry.
The BMI-GRS demonstrated positive, age-adjusted associations with total body and regional fat, lean and bone mass. Associations of the greatest magnitude were observed for adiposity variables, intermediate associations were observed for lean tissue variables, and associations of the lowest magnitude were observed for bone variables. This pattern was repeated across all body regions and mirrored the relationship between measured BMI and body composition in this participant group. It is also consistent with the relationship between measured BMI and BF% previously reported in adult populations24, 25 and replicated in this study (Figure 1).
We found a significant, positive association between the BMI-GRS and BF%. This corroborates and extends the results of other adult studies.13, 14, 26, 27 The most recent study of a BMI-GRS and BF% in a European population reported that each BMI-increasing allele was associated with a 0.14% (95% CI: 0.05, 0.24) increase in BF% (P=0.004).26 The study included 31 SNPs combined to form an unweighted score and 1578 adult participantsparticipants aged 35–74 years. Our results confirm this positive association using a weighted BMI-GRS comprised of a much greater number of BMI-associated variants and a sample size over six times as large (9667 versus 1578 participants).
The BMI-GRS was positively associated with WHR and reached statistical significance, even after Bonferroni correction. The findings from two previous studies of a BMI-GRS on WHR were directionally consistent with our findings but were not statistically significant.15, 26 This might be explained by the smaller sample size and lower statistical power of those studies, which included 1578 and 740 participants, respectively.15, 26
The association of the BMI-GRS with VAT/SAT ratio, while positive, did not reach statistical significance. To our knowledge, no other study has investigated a BMI-GRS in relation to VAT/SAT ratio. This novel finding suggests that the genetic regulation of BMI is largely independent of mechanisms that regulate the relative distribution of visceral and subcutaneous fat in the abdominal region. This supports investigations using different methods that show only modest overlap between the genetic regulation of BMI and VAT/SAT ratio. For example, only seven out of 32 BMI-related loci were associated with VAT/SAT ratio among adults of European ancestry in one study21 and only one out of 12 BF%-associated SNPs was associated with VAT/SAT ratio in another study.23
The discrepancy between findings for WHR and VAT/SAT ratio in our study suggests the possibility that WHR and VAT/SAT measure different aspects of central adiposity. While WHR provides a measure of central relative to peripheral fat, VAT/SAT ratio measures the relative distribution of internal and sub-cutaneous fat within the abdominal region. An alternative explanation is that differential measurement error is not consistent between WHR and VAT/SAT.
We found significant, positive associations between the BMI-GRS and WC, HC and weight, but not with height. These findings are consistent with the results of previous studies.13, 14, 15, 26
In this cohort, 28 SNPs exhibited nominally significant individual associations with BMI. All 28 showed a greater magnitude of association with fat mass than lean or bone mass (Supplementary Table 1). The SNPs clustered into two main groups by their associations with body composition. Approximately half (n=15) were associated with a global increase in fat, lean and bone mass, while the remaining 13 SNPs exhibited more adipose-specific effects. The SNP tagging MTCH2 is notable for a nominally significant, negative association with bone mass. The SNP tagging MC4R shows significant and similar positive effects on fat, lean and bone mass. This is in keeping with the observation that MC4R-deficient individuals have increased lean mass and bone mineral density in addition to increases in fat mass.28
This study has several strengths. To our knowledge, it is the first to examine the relationship between a BMI-GRS and fat, lean and bone mass across body regions. We present novel findings representing an extension of previous investigations through increased sample size, SNP number and body composition outcome measures.
The results of this study pertain only to adults aged 29–64 years. Many genetic variants are associated with BMI throughout life.29 However, associations between specific BMI-related loci and some body composition phenotypes are age-dependent.30, 31 For example, a meta-analysis of data from four birth cohort studies did not find an association between a BMI-GRS (comprised of 16 SNPs related to adult BMI) and BF% at 1 year, 2–3 years or 4–5 years.30 The BMI-GRS was associated with both fat and lean mass, suggesting that BMI-related loci promote gains in both fat and lean mass in early life. Other studies have reported inconsistent associations between specific SNPs and BMI at different stages of adulthood.31, 32 In general, declining effect sizes in older adults may reflect the greater accumulation of lifestyle factors in later adulthood or cohort effects.31
The results of this study cannot be extrapolated to individuals who exceed the weight limit of the DXA scanner (140 kg), those of non-European descent or those with known metabolic disease. The results for VAT, SAT and VAT/SAT ratio cannot be extrapolated to those with less than 1g of visceral android fat as the DXA scanner software was unable to provide VAT estimates below this limit. Our sample size limited the power to robustly detect associations between individual SNPs and body composition. Finally, the cross-sectional nature of the study did not facilitate exploration of causal relationships between the components of body composition.
In conclusion, we show that, in combination, 96 BMI-related genetic variants are positively associated with adiposity, with intermediate effects on lean mass and weaker effects on bone mass. This pattern mirrors the relationship between measured BMI and body composition in this age group. The findings support the use of the BMI-GRS for causal modelling of the impact of adult BMI and total body adiposity on health and other outcomes in both men and women.
Future research could explore whether BMI-related variants with apparent adipose-specific effects operate through distinct mechanisms to BMI-related variants associated with a global increase in mass.
References
World Health Organization ICD-10: The ICD-10 International Statistical Classification of Diseases and Related Health Problems. World Health Organization: Geneva, 1992.
Moody A Adult anthropometric measures, overweight and obesity. In: Craig R, Mindell J (eds). Health Survey for England - 2013. Health and Social Care Information Centre: London, 2014. p 1–39.
Bouchard C . BMI, fat mass, abdominal adiposity and visceral fat: where is the 'beef'? Int J Obes 2007; 31: 1552–1553.
Shah NR, Braverman ER . Measuring adiposity in patients: The utility of body mass index (BMI), percent body fat, and leptin. PLoS One 2012; 7: e33308.
Lear SA, Humphries KH, Kohli S, Birmingham CL . The use of BMI and waist circumference as surrogates of body fat differs by ethnicity. Obesity 2007; 15: 2817–2824.
Romero-Corral A, Somers VK, Sierra-Johnson J, Thomas RJ, Collazo-Clavell ML, Korinek J et al. Accuracy of body mass index in diagnosing obesity in the adult general population. Int J Obes (Lond) 2008; 32: 959–966.
Frankenfield DC, Rowe WA, Cooney RN, Smith JS, Becker D . Limits of body mass index to detect obesity and predict body composition. Nutrition 2001; 17: 26–30.
Langenberg C, Sharp SJ, Schulze MB, Rolandsson O, Overvad K, Forouhi NG et al. Long-term risk of incident type 2 diabetes and measures of overall and regional obesity: the EPIC-InterAct case-cohort study. PLoS Med 2012; 9: e1001230.
Despres J-P . Body fat distribution and risk of cardiovascular disease: an update. Circulation 2012; 126: 1301–1313.
Petursson H, Sigurdsson JA, Bengtsson C, Nilsen TIL, Getz L . Body configuration as a predictor of mortality: comparison of five anthropometric measures in a 12 year follow-up of the Norwegian HUNT 2 study. PLoS One 2011; 6: e26621.
Elks CE, den Hoed M, Zhao JH, Sharp SJ, Wareham NJ, Loos RJF et al. Variability in the heritability of body mass index: a systematic review and meta-regression. Front Endocrinol (Lausanne) 2012; 3: 1–16.
Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015; 518: 197–206.
Speliotes EK, Willer CJ, Berndt SI, Monda KL, Thorleifsson G, Jackson AU et al. Association analyses of 249796 individuals reveal 18 new loci associated with body mass index. Nat Genet 2010; 42: 937–948.
Willer CJ, Speliotes EK, RJF Loos, Li S, Lindgren CM, Heid IM et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat Genet 2009; 41: 25–34.
Badsi MN, Mediene-Benchekor S, Ouhaibi-Djellouli H, Lardjam-Hetraf SA, Boulenouar H, Meroufel DN et al. Combined effect of established BMI loci on obesity-related traits in an Algerian population sample. BMC Genet 2014; 15: 128.
Bauer F, Elbers CC, Adan RAH, Loos RJF, Onland-Moret NC, Grobbee DE et al. Obesity genes identified in genome-wide association studies are associated with adiposity measures and potentially with nutrient-specific food preference. Am J Clin Nutr 2009; 90: 951–959.
Rolfe EDL, Stolk RP, Ekelund U, Griffin SJ, Forouhi NG, Loos RJF et al. Association between birth weight and visceral fat in adults. Am J Clin Nutr 2010; 92: 347–352.
Howie BN, Donnelly P, Marchini J . A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet 2009; 5: e1000529.
Durbin RM, Altshuler DL, Durbin RM, Abecasis GR, Bentley DR, Chakravarti A et al. A map of human genome variation from population-scale sequencing. Nature 2010; 467: 1061–1073.
Shungin D, Winkler TW, Croteau-Chonka DC, Ferreira T, Locke AE, Magi R et al. New genetic loci link adipose and insulin biology to body fat distribution. Nature 2015; 518: 187–196.
Fox CS, Liu Y, White CC, Feitosa M, Smith AV, Heard-Costa N et al. Genome-wide association for abdominal subcutaneous and visceral adipose reveals a novel locus for visceral fat in women. PLoS Genet 2012; 8: e1002695.
Kilpeläinen TO, Zillikens MC, Stan A, Finucane FM, Ried JS, Langenberg C et al. Genetic variation near IRS1 associates with reduced adiposity and an impaired metabolic profile. Nat Genet 2012; 43: 753–760.
Lu Y, Day FR, Gustafsson S, Buchkovich ML, Na J, Bataille V et al. New loci for body fat percentage reveal link between adiposity and cardiometabolic disease risk. Nat Commun 2016; 7: 10495.
Sung Y-A, Oh J-Y, Lee H . Comparison of the body adiposity index to body mass index in Korean women. Yonsei Med J 2014; 55: 1028–1035.
Flegal KM, Shepherd JA, Looker AC, Graubard BI, Borrud LG, Ogden CL et al. Comparisons of percentage body fat, body mass index, waist circumference, and waist-stature ratio in adults. Am J Clin Nutr 2009; 89: 500–508.
Goumidi L, Cottel D, Dallongeville J, Amouyel P, Meirhaeghe A . Effects of established BMI-associated loci on obesity-related traits in a French representative population sample. BMC Genet 2014; 15: 62.
Zhu J, Loos RJF, Lu L, Zong G, Gan W, Ye X et al. Associations of genetic risk score with obesity and related traits and the modifying effect of physical activity in a Chinese Han population. PLoS One 2014; 9: e91442.
Farooqi IS, O’Rahilly S . Monogenic obesity in humans. Annu Rev Med 2005; 56: 443–458.
Bradfield JP, Taal HR, Timpson NJ, Scherag A, Lecoeur C, Warrington NM et al. A genome-wide association meta-analysis identifies new childhood obesity loci. Nat Genet 2012; 44: 526–531.
Elks CE, Heude B, de Zegher F, Barton SJ, Clément K, Inskip HM et al. Associations between genetic obesity susceptibility and early postnatal fat and lean mass. JAMA Pediatr 2014; 168: 1122–1130.
Winkler TW, Justice AE, Graff M, Barata L, Feitosa MF, Chu S et al. The influence of age and sex on genetic associations with adult body size and shape: a large-scale genome-wide interaction study. PLoS Genet 2015; 11: e1005378.
Murphy RA, Nalls MA, Keller M, Garcia M, Kritchevsky SB, Tylavsky FA et al. Candidate Gene Association Study of BMI-related loci, weight, and adiposity in old age. J Gerontol Ser A Biol Sci Med Sci 2013; 68: 661–666.
Acknowledgements
The Fenland Study is supported by the Medical Research Council (MC_U106179471). This work was supported by the Medical Research Council [Unit Programme numbers MC_UU_12015/2 and MC_UU_12015/1]. Genotyping was supported by the Medical Research Council (MC_PC_13046). We are grateful to all the volunteers for their time and help and to the General Practitioners and practice staff for assistance with recruitment. We thank the Fenland Study Investigators, Fenland Study Co-ordination team and the Epidemiology Field, Anthropometry, Data and Laboratory teams. Biochemical assays were performed by the National Institute for Health Research, Cambridge Biomedical Research Centre, Core Biochemistry Assay Laboratory and the Cambridge University Hospitals NHS Foundation Trust, Department of Clinical Biochemistry.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on International Journal of Obesity website
Rights and permissions
About this article

Cite this article
Clifton, E., Day, F., De Lucia Rolfe, E. et al. Associations between body mass index-related genetic variants and adult body composition: The Fenland cohort study. Int J Obes 41, 613–619 (2017). https://doi.org/10.1038/ijo.2017.11
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ijo.2017.11
- Springer Nature Limited
This article is cited by
-
Opportunities and challenges for the use of common controls in sequencing studies
Nature Reviews Genetics (2022)
-
Independent and combined associations between fast-food outlet exposure and genetic risk for obesity: a population-based, cross-sectional study in the UK
BMC Medicine (2021)
-
Convergence between biological, behavioural and genetic determinants of obesity
Nature Reviews Genetics (2017)