
Abstract
Grassland birds are the most endangered bird group in Uruguay, mainly due to habitat loss. Livestock farming is the most widespread agricultural activity in Uruguay. Grazing practices modify grassland plant structure. The most pervasive grazing systems in the country are continuous and rotational grazing, although with high diversity in their application. We studied the plant structure and the grassland bird community in 40 cattle ranches in eastern Uruguay, which include continuous grazing systems (CGSs) and rotational grazing systems (RGSs). RGS had a higher average grass height and greater habitat heterogeneity than CGSs, with a difference of 3.7 cm in grass height, 3.1 cm more heterogeneity within paddocks, and 2.1 cm more heterogeneity between paddocks. Also, RGS had a greater richness and an effective number of species per farm than CGS, with averages of 2.8 more species and 7.7 more effective number of species in RGS in relation to CGS. The presence of four species was significantly greater in RGS and one in CGS. However, there were no exclusive species to any grazing system. Our study shows a discreet positive effect of RGS on the ranch’s grassland bird community, probably related to the structural diversity generated by these systems. Contrary to other works, this is the first work that reports a difference in species richness between rotational and continuous grazing systems. We, therefore, conclude that the grazing system is a relevant factor when managing grasslands to enhance bird biodiversity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Grassland birds respond to vegetation structure (e.g., height and leaf density) (MacArthur and MacArthur 1961; Norment et al. 1999; Isacch et al. 2004; Goijman and Zaccagnini 2008; Fisher and Davis 2010; Isacch and Cardoni 2011; Dias et al. 2014). They occupy diverse ecological niches, there being tall grasslands specialists, short grasslands specialists, and others that use a wide range of heights (Azpiroz et al. 2012). Therefore, many studies have concluded the need for conservation practices that create a mosaic of vegetation structure (Fuhlendorf et al. 2006; Davis et al. 2020; Sliwinski et al. 2020).
South American southern cone grasslands are one of the few and largest temperate prairies and savannahs in the world (Soriano 1992). Since the sixteenth century, cattle introduction has considerably transformed these grasslands (Modernel et al. 2016). Cattle grazing has generated a shortening and homogenization of the vegetation structure, concentrating it between the first 10 cm of height (Soriano 1992; Altesor et al. 1998, 2006; Rodríguez et al. 2003). These modifications to vegetation’s structure have resulted in habitat loss for grassland birds, mainly tall grasslands specialists, and their subsequent population decline (Comparatore et al. 1996; Isacch and Martínez 2001; Isacch et al. 2003; Azpiroz and Blake 2009; Dias et al. 2017).
Traditional continuous grazing systems (CGSs) and rotational grazing systems (RGSs) are the prevalent grazing methods in the region. Traditional CGS consists of permanent grazing throughout the year in every paddock. In RGS, cattle are concentrated in a few groups and moved periodically, alternating periods of occupation and rest in every paddock (Briske et al. 2008). In CGS, cattle can graze selectively for preferred species as they can access to the entire grazing area. In RGS, cattle have less area since they are more concentrated than in CGS and therefore graze more intensively and homogenously while other paddocks are resting (Bailey and Brown 2011). There is no scientific consensus on whether any of these systems are productively superior to the other (Briske et al. 2008; Barnes et al. 2008; Briske et al. 2008). However, many traditional CGSs are associated with grassland degradation and overgrazing due to selective grazing and minimal management decision (Kothmann 2009). The response of birds to different grazing systems as habitat generators for a diverse bird assemblage has been poorly evaluated and with different results (Isacch and Cardoni 2011; Murray et al. 2016; Pipher et al. 2016; M. Sliwinski et al. 2019; Milligan et al. 2020; Codesido and Bilenca 2021). Cozzani and Zalba (2009) propose to conserve patches of tall tussock as bird refuges, but this management was not incorporated into the overall management of the farms. Therefore, its viability in commercial terms has not been evaluated. Knowing the viability of a biodiversity management action in commercial farms is necessary for farmers to apply them (Neilly et al. 2018; Aldabe et al. 2019).
According to MacArthur and MacArthur (1961), there is a greater diversity of birds in an ecosystem when there is greater structural diversity. Compared to traditional CGS, RGSs are expected to have greater richness and diversity of birds given their greater structural diversity associated with more diversity of ecological niches, as a result of the greater temporal-spatial variation in grazing intensity between paddocks. In this paper, we evaluate the vegetation structure and grassland bird communities in RGS and traditional CGS in eastern Uruguay.
Methods
Study area
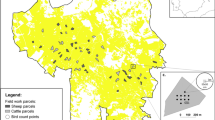
The present study was conducted on 40 cattle ranches in the states of Maldonado and Lavalleja in eastern Uruguay (34°12′10″ S; 54°45′48″ W) (Fig. 1). The ranches were located in the geomorphological unit of Sierras del Este in a range of heights from 0 to 500 m a.s.l. (Brazeiro et al. 2012), where plains and hills (sierras) are prevalent (Baeza et al. 2019). Ranches were equally distributed between hills and plains. The grasslands of the hills are located on shallow rocky soils, with moderate slopes and the presence of trees, given their proximity to rocky woodlands (bosque serrano). The grasslands of the plains are located on deep and fertile soils where the grasses develop with greater density and less presence of trees (Lezama et al. 2011).

Ranch’s location where we conducted the study during spring of 2015 and the spring of 2016 to the summer of 2017
The average ranch area was 250 hectares, with a maximum of 1290 hectares and a minimum of 40 hectares. Of the 40 ranches sampled the first year, 24 were CGS, and 16 were RGS. In the second year, there were 8 CGS and 32 RGS since several ranches changed their grazing system that year. Grazing systems were classified in conjunction with managers. The RGS varied between ranches, with an average of 10 paddocks, and at least a third were resting at any moment. The RGS had more than one herd of cattle. Paddock occupation time varied between 5 and 25 days, while resting time ranged from 20 days to 6 months. On the other hand, CGS had an average of 9 paddocks, and only a few had a single paddock without grazing, although it occupied a small proportion of the ranch area compared to RGS. Seventy-five percent of the sampled paddocks corresponded to natural grasslands, and the remaining were artificial pastures. RGS had 74% of their paddocks with natural grasslands, while CGSs had 78% with natural grasslands.
Bird sampling
Two samples from each ranch were recorded in two different seasons: the first from October to December 2015 and the second from December 2016 to January 2017, taking this time for the higher activity of birds and the presence of summer visitors and migrants. Due to the typical inter-annual climate variability present in the country, the first sampling was during a drought season, while the second was rainy. Bird surveys consisted of two observers simultaneously counting all visible birds along six transects per ranch (300 m long), trying to locate each transect in a different paddock. Transects were located far from fences and neighboring woodlands to avoid measuring their effects on the presence or absence of some species (birds use fences as perches, and forests may attract other species unrelated to grasslands). On CGS, six transects were located in grazed paddocks. On RGS, three transects were located in grazed paddocks and three in resting paddocks. The latter were identified with the help of managers or by evaluating grasses’ state and absence of fresh dung. We counted all birds observed or heard along the transect that used the paddock. We conducted the surveys between 07:00 and 10:00 and 16:00 and 19:00 when the birds are known to be most active.
Environmental variables sampling
At each transect, we sampled grass height and percentage of tree cover. Grass height was measured every 50 m with Robel’s method, recording six measurements, and then averaged. The method measures the visual obstruction caused by vegetation on a 1.5 m high pole divided into 10 cm segments (Robel et al. 1970). The observer records the lowest visible segment on the pole from a 4 m distance and 1 m height. The percentage of tree cover for the whole paddock was estimated visually during sampling. The percentage of artificial pasture cover was measured by satellite image.
Data modeling
Samples between years were assumed to be independent, given the marked climatic differences between years, which led to significant differences in grass height (see “Results”). Therefore, a sample of 80 observations was distributed in 40 ranches. Statistical analyses included (1.) modeling of plant structure and (2.) modeling of grassland birds’ community. All analyses were conducted with R 4.0.0 (R Core Team, 2020).
Relation of the grazing system to the vegetation structure
We evaluated the differences in vegetation structure between ranches using multiple linear regression with the grazing system, season (rainy or dry), and topographic area (plains or hills) as explanatory variables. We used three variables as descriptors of vegetation structure: average grass height, structural heterogeneity between paddocks (HBP), and structural heterogeneity within paddocks (HWP). HBP corresponded to the variability in grass height between paddocks and was estimated as the standard deviation of the average grass height of the ranch paddocks. HWP corresponded to the variability in grass height within paddocks and was estimated as the average of the standard deviation of the grass height of each ranch paddock.
Relation of the grazing system to the bird community
We evaluated the grassland birds’ community using bird richness per ranch and the effective number of species per ranch (ENS) as response variables (Jost 2010). Bird richness corresponded to the number of grassland bird species observed on each ranch. We used ENS as an alternative to conventional biodiversity indices, which Jost (2010) defined as the exponential of Shannon’s diversity index (Shannon 1948). Conventional biodiversity indices are non-intuitive, non-linear parameters that are difficult to compare with each other (Jost 2010). We classified grassland birds based on Azpiroz et al. (2012) and personal observations.
We run statistical models with these two variables to assess the relationship between grassland birds with the following explanatory variables: grazing system, percentage of tree cover (average of transects data), percentage of natural cover, topographic zone (hills or plains), ranch area, and season (dry or rainy). A generalized linear model (GLM) with binomial error distribution evaluated the effect of the grazing systems on the probability of occurrence of each species. A GLM with Poisson error distribution evaluated the effect of the grazing systems and other environmental variables on species richness per ranch. Finally, a linear model (LM) with normal distribution evaluated the effect of the grazing systems and other environmental variables on the ENS.
In all analyses, we considered a significance value of (α) ≤ 0.05. We used a stepwise variable selection method to obtain the minimally adequate model, with Akaike information criteria (AIC) as selection criteria (Akaike 1974). For the LM, we verified the normality assumption with the Jarque and Bera (1987) test and logarithmic transformations were performed, if needed. We performed Breusch and Pagan (1979) tests to evaluate the homoscedasticity assumption. We evaluated outliers with Cook’s distance (Cook 1977). We used the visreg package for the graphical representation of the models (Breheny and Burchett 2017?), which plots how the expected values of the response variable change depending on the explanatory variable chosen, maintaining the other explanatory variables fixed. R2 and quasi R2 were calculated to assess the percentage of variability explained by the explanatory variables included in the models.
Before modeling, we standardized continuous explanatory variables (ranch area, percentage of tree cover, and percentage of natural cover) to allow comparison between model coefficients (Schielzeth 2010). In addition, we calculated the effect size and their confidence intervals, as proposed by Nakagawa and Cuthill (2007). The use of effect size is because hypothesis tests estimate the significance of explanatory variables but do not estimate the magnitude of the effect of interest or the precision of that estimate. We interpret effect size as small effect (between 0.2 and 0.5), medium effect (between 0.5 and 0.8), or large effect (more than 0.8) (Cohen 2013).
Results
Vegetation structure
Grass height ranged from 1.8 to 43.5 cm between ranches, averaging 11.4 ± 8.0 cm. The LM determined a significant effect of the grazing system and season over grass height (Table 1), with an adjusted R2 of 55% (Table 2). Based on the model coefficients, grass height was 3.68 cm higher in RGS than in CGS (difference between both treatments’ estimates). Also, grass height was 4.24 cm higher in the rainy season than in the dry season. Figure 2 graphs the model results between grazing treatments.

Linear models for vegetation structure variables. They show the positive effect of rotational systems (RGSs) over grass height (left), structural heterogeneity between paddocks (center), and structural heterogeneity within paddocks (right). Solid lines represent the fitted equation, and gray-shaded areas represent the 95% confidence intervals
The HBP ranged from 1.0 to 44.7 cm between ranches, with an average of 6.4 ± 5.7 cm. The LM determined a significant effect of the grazing system and season over HBP (Table 1), with an adjusted R2 of 39% (Table 3). Based on the model coefficients, HBP was 2.08 cm higher in RGS than in CGS (difference between both treatments’ estimates). Also, HBP was 1.95 cm higher in the rainy season than in the dry season.
The structural HWP ranged from 1.0 to 27.9 cm between ranches with an average of 7.6 ± 4.8 cm. The LM determined a significant effect of the grazing system and season over HWP (Table 1), with an adjusted R2 of 31% (Table 4). Based on the model coefficients, HWP was 3.06 cm higher in RGS than in CGS (difference between both treatments’ estimates). Also, HBP was 1.56 cm higher in the rainy season than in the dry season. The topographic zone effect was included in the model but was not significant.
Bird community
Relation of the type of grazing and presence of different species of birds
We recorded 53 grassland bird species, of which 12 were grassland obligate species, and three were globally threatened species (Table 5). The five most common species were Furnarius rufus, Molothrus bonariensis, Nothura maculosa, Tyrannus savana, and Vanellus chilensis. The GLM to assess the effect of grazing systems on the probability of occurrence of each species found significant effects for five species. According to the models, the occurrence of four species was significantly higher in RGS: Embernagra platensis, Phacellodomus striaticollis, Progne tapera, and Sicalis flaveola. On the other hand, the occurrence of one species was significantly higher in CGS: Nengetus cinereus. Only one grassland obligate species (E. platensis) responded to the grazing system. Both species responded positively to the RGS. However, no significant effect of the grazing system on the threatened species was detected (Table 5).
Relation of grazing system with species richness and ENS per ranch
The species richness per ranch ranged from 10 to 29 species, with an average of 20.4 ± 4.08 species. The GLM determined a significant effect of the grazing system on species richness (Table 6), with an adjusted quasi R2 of 11% (Table 7). The model detected a significant difference in species richness between the different grazing systems, with an estimated mean of 21.5 species and 18.7 species in the RGS and CGS, respectively (Fig. 3). The other explanatory variables showed no significant effect (percentage of tree cover, percentage of natural cover, topographic area, ranch area, and season). Cohen’s d estimated a medium effect size of grazing system over species richness (d = 0.48, lower limit = 0.46, upper limit = 1.41).

Generalized linear model (left) and linear model (right) for bird community variables. They show the positive effect of rotational systems (RGSs) on species richness (left) and the effective number of species (right). Solid lines represent the fitted equation, and gray-shaded areas represent the 95% confidence intervals
The ENS per ranch ranged from 86 to six species, averaging 45.7 ± 16.5 effective number of species. The LM determined a significant effect of the grazing system over ENS (Table 8), with an adjusted R2 of 4% (Table 9). The model detected a significant difference in ENS between the different grazing systems, with an estimated mean of 48.8 species and 41.1 species in the RGS and CGS, respectively. The other explanatory variables showed no significant effect (percentage of tree cover, percentage of natural cover, topographic area, ranch area, and season). Cohen’s d estimated a medium effect size of grazing system over ENS (d = 0.55, lower limit IC = 0.46, upper limit IC = 1.56).
Discussion
Grassland birds are Uruguay’s most endangered bird group (Aldabe et al. 2013). Knowing the effect of different management practices is necessary for their conservation. We hypothesized that rotational grazing systems have more diverse bird communities than continuous grazing systems because the former have more structurally heterogeneous grasslands. Our results support this hypothesis. Our models found significantly higher heterogeneity between and within paddocks in RGS than CGS. In addition, contrary to other works in the Rio de la Plata Grasslands (Isacch and Martínez 2001; Codesido and Bilenca 2021), we found significantly more diverse grassland bird communities in RGS compared to CGS. Our success in detecting such an effect might be explained by the much larger number of ranches assessed here. The positive effect of structural heterogeneity on the grassland bird community is consistent with other studies (Fuhlendorf et al. 2006; Davis et al. 2020; Sliwinski et al. 2020). However, the magnitude of the treatment’s effect on the bird community was moderate, considering the low explained variability of our models (R2). This may be due to the homogenization of treatments as a result of drought during the first year of sampling.
As for structural heterogeneity, RGS had higher HBP than CGS. This was because RGS had two types of paddocks: rested and grazed. Grass height in the rested paddocks was 67% greater than in the grazed ones. Contrary to what we expected, RGS also had higher structural HWP. This was not expected given the more intensive and homogeneous grazing of paddocks that theoretically occurs in RGS (Bailey and Brown 2011). This may be because the RGS included in this study had medium-sized paddocks, which avoids intensive grazing and, thus, the homogenization of the grass structure within the paddock.
As for the bird community, on average, 13% more species per ranch were observed on RGS, a relatively small difference of 2.8 species. Potentially, a more considerable difference could be expected given the wide range of tall grass species that could be found on the ranches, but no such species were registered (Azpiroz et al. 2012). Based on our statistical models, the grazing system explained 4% and 11% of the variability in the richness and the effective number of species present on the ranches, respectively. This indicates that there are other important variables responsible for the bird community’s variability that we did not consider. Additionally, the drought period may have also affected both treatments. This climatic event might explain the lower structural heterogeneity shown by our models and resulted in all paddocks being heavily grazed because of food shortages. This may partially explain the moderate effect of treatments on the bird community. Furthermore, other studies showed that grazing systems alone do not have an effect on the grassland bird community and that longer rest periods are needed (Sliwinski et al. 2019) or even land sparing (Dotta et al. 2016). Another possible explanation is that in most CGS, there was at least one patch of tall tussock (mainly of Paspalum quadrifolium and Erianthus angustifolius) or paddock under deferment, which could replace the role of resting paddocks in RGS, and therefore homogenizing the effect of the treatments.
No species were recorded exclusively for any of the treatments. The moderate effect of the grazing system on the bird community could be due to the species identified in our probability of occurrence models. Embernagra platensis, Phacellodomus striaticollis, Progne tapera, and Sicalis flaveola were mostly benefited by RGS. These ranches had a higher grass height than CGS and corresponded with the habitat requirements of these species, which prefer medium and tall grassland sites (Azpiroz and Blake 2009; Dias et al. 2014). Likewise, the presence of P. tapera, considering that it is insectivorous, may be due to the higher abundance of insects in taller grasslands, given the higher biomass (Kruess and Tscharntke 2002).
As for the weaknesses of our study, it was complex to classify the ranches according to the grazing system. This was because several of the sampled CGS were in the process of transitioning to rotational systems. Moreover, we surveyed equal amounts of grazed and resting paddocks in the RGS, which did not necessarily reflect reality. In addition, we did not consider landscape features that might explain local species richness differences among ranches.
Our results suggest that RGS can have more grassland bird species than CGS if CGSs have a high grazing intensity that prevents the paddocks from developing HWP. If the grazing intensity is low, which generates HWP, we cannot discard that CGSs have the same or more grassland bird species than RGS. In this sense, it is necessary to include grazing intensity data and stocking levels (number of livestock and kg) to compare the grassland bird community between grazing systems.
Conclusions
The rotational grazing systems studied had higher and more heterogeneous grasslands than continuous grazing systems, possibly due to the variation in the temporal and special grazing pressure of the RGS.
The effect of the grazing system on the grassland birds’ community was low, possibly due to the water deficit that occurred during the samplings that limited grass growth and to the influence of tall tussocks as substitutes for tall grasses. In this study, we showed that the way ranchers manage their livestock grazing influences species richness. Considering the high percentage of grassland area on private hands in the Rio de la Plata grasslands, this result suggests the importance of grazing systems on grassland bird conservation. However, to confirm if rotational grazing systems promote higher bird species richness, future efforts should be focused on assessing differences between continuous and rotational grazing systems with more contrasting vegetation structures, considering grazing intensity and landscape effects.
Data availability
Not applicable.
Code availability
Not applicable.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723. https://doi.org/10.1109/TAC.1974.1100705
Aldabe J, Arballo E, Caballero-Sadi D et al (2013) Aves. In: Soutullo A, Clavijo C, Martínez-Lanfranco J (eds) Especies prioritarias para la conservación en Uruguay. SNAP/DINAMA/MVOTMA and DICUT/MEC, Montevideo, pp 149–173
Aldabe J, Lanctot RB, Blanco D, Rocca P, Inchausti P (2019) Managing grasslands to maximize migratory shorebird use and livestock production. Rangel Ecol Manag 72:150–159. https://doi.org/10.1016/j.rama.2018.08.001
Altesor A, Di Landro E, May H, Ezcurra E (1998) Long-term species change in a Uruguayan grassland. J Veg Sci 9:173–180. https://doi.org/10.2307/3237116
Altesor A, Piñeiro G, Lezama F et al (2006) Ecosystem changes associated with grazing in subhumid South American grasslands. J Veg Sci 17:323–332. https://doi.org/10.1111/j.1654-1103.2006.tb02452.x
Azpiroz AB, Blake JG (2009) Avian assemblages in altered and natural grasslands in the northern Campos of Uruguay. Condor 111:21–35. https://doi.org/10.1525/cond.2009.080111
Azpiroz AB, Isacch JP, Dias RA et al (2012) Ecology and conservation of grassland birds in southeastern South America: a review. J Field Ornithol 83:217–246. https://doi.org/10.1111/j.1557-9263.2012.00372.x
Baeza S, Rama G, Lezama F et al (2019) Cartografía de los pastizales naturales en las regiones geomorfológicas de Uruguay predominantemente ganaderas. Ampliación y actualización. In Altesor A, López-Mársico L, Paruelo JM (Eds) Bases ecológicas y tecnológicas para el manejo depastizales II. INIA, Montevideo, pp 27–47
Bailey DW, Brown JR (2011) Rotational grazing systems and livestock grazing behavior in shrub-dominated semi-arid and arid rangelands. Rangel Ecol Manag 64:1–9. https://doi.org/10.2111/REM-D-09-00184.1
Barnes MK, Norton BE, Maeno M, Malechek JC (2008) Paddock size and stocking density affect spatial heterogeneity of grazing. Rangel Ecol Manag 61:380–388. https://doi.org/10.2111/06-155.1
Brazeiro A, Panario D, Soutullo A et al (2012) Clasificación y delimitación de las eco-regiones de Uruguay. Informe Técnico. Convenio MGAP/PPR–Facultad de Ciencias/Vida Silvestre/Sociedad Zoológica del Uruguay/CIEDUR 40. https://doi.org/10.13140/2.1.1328.0328
Breheny P, Burchett W (2017) Visualization of regression models using visreg. R J 9:56–71
Breusch TS, Pagan AR (1979) A simple test for heteroscedasticity and random coefficient variation. Econometrica: J Econometric Soc 47:1287–1294. https://doi.org/10.2307/1911963
Briske DD, Derner JD, Brown JR et al (2008) Rotational grazing on rangelands: reconciliation of perception and experimental evidence. Rangel Ecol Manag 61:3–17. https://doi.org/10.2111/06-159R.1
Codesido M, Bilenca D (2021) Influencia de la intensidad de pastoreo sobre ensambles de aves en espartillares de la Bahía de Samborombon, Argentina. Hornero 36:21–30
Cohen J (2013) Statistical power analysis for the behavioral sciences. Hoboken. NJ: Taylor and Francis. https://doi.org/10.4324/9780203771587
Comparatore VM, Martínez MM, Vassallo AI et al (1996) Abundancia y relaciones con el hábitat de aves y mamíferos en pastizales de Paspalum quadrifarium (paja colorada) manejados con fuego (Prov. de Buenos Aires, Argentina). Interciencia 2:228–237
Cook RD (1977) Detection of influential observation in linear regression. Technometrics 19:15–18. https://doi.org/10.2307/1268249
Cozzani N, Zalba SM (2009) Estructura de la vegetación y selección de hábitats reproductivos en aves del pastizal pampeano. Ecol Austral 19:35–44
Davis KP, Augustine DJ, Monroe AP, Derner JD, Aldridge CL (2020) Adaptive rangeland management benefits grassland birds utilizing opposing vegetation structure in the shortgrass steppe. Ecol Appl 30:e02020. https://doi.org/10.1002/eap.2020
Dias RA, Bastazini VA, Gianuca AT (2014) Bird-habitat associations in coastal rangelands of southern Brazil. Iheringia, Ser Zool 104:200–208. https://doi.org/10.1590/1678-476620141042200208
Dias RA, Gianuca AT, Vizentin-Bugoni J, Gonçalves MSS et al (2017) Livestock disturbance in Brazilian grasslands influences avian species diversity via turnover. Biodivers Conserv 26:2473–2490. https://doi.org/10.1007/s10531-017-1370-4
Dotta G, Phalan B, Silva TW et al (2016) Assessing strategies to reconcile agriculture and bird conservation in the temperate grasslands of South America. Conserv Biol 30:618–627. https://doi.org/10.1111/cobi.12635
Fisher RJ, Davis SK (2010) From Wiens to Robel: a review of grassland-bird habitat selection. J Wildl Manag 74:265–273. https://doi.org/10.2193/2009-020
Fuhlendorf SD, Harrell WC, Engle DM et al (2006) Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 16:1706–1716. https://doi.org/10.1890/1051-0761(2006)016[1706:shbtbf]2.0.co;2
Goijman AP, Zaccagnini ME (2008) The effects of habitat heterogeneity on avian density and richness in soybean fields in Entre Ríos, Argentina. Hornero 23:67–76
Isacch JP, Cardoni DA (2011) Different grazing strategies are necessary to conserve endangered grassland birds in short and tall salty grasslands of the flooding Pampas. Condor 113:724–734. https://doi.org/10.1525/cond.2011.100123
Isacch JP, Martínez MM (2001) Estacionalidad y relaciones con la estructura del hábitat de la comunidad de aves de pastizales de paja colorada (Paspalum quadrifarium) manejados con fuego en la provincia de Buenos Aires, Argentina. Ornitol Neotrop 12:345–354
Isacch JP, Bo MS, Maceira NO et al (2003) Composition and seasonal changes of the bird community in the west pampa grasslands of Argentina. J Field Ornithol 74:59–65. https://doi.org/10.1648/0273-8570-74.1.59
Isacch JP, Holz S, Ricci L, Martínez MM (2004) Post-fire vegetation change and bird use of a salt marsh in coastal Argentina. Wetlands 24:235–243. https://doi.org/10.1672/0277-5212(2004)024[0235:PVCABU]2.0.CO;2
IUCN (2022) The IUCN red list of threatened species. Version 2022–1. https://www.iucnredlist.org. Accessed 26 October 2022
Jarque CM, Bera AK (1987) A test for normality of observations and regression residuals. Int Stat Rev 55:163–172. https://doi.org/10.2307/1403192
Jost L (2010) The new synthesis of diversity indices and similarity measures. http://www.loujost.com/Statistics%20and%20Physics/Diversity%20and%20Similarity/DiversitySimilarityHome.htm. Accessed 26 October 2022
Kothmann M (2009) Grazing methods: a viewpoint. Rangelands 31:5–10. https://doi.org/10.2111/1551-501X-31.5.5
Kruess A, Tscharntke T (2002) Contrasting responses of plant and insect diversity to variation in grazing intensity. Biol Conserv 106:293–302. https://doi.org/10.1016/S0006-3207(01)00255-5
Lezama F, Altesor A, Pereira M, Paruelo J (2011) Descripción de la heterogeneidad florística en los pastizales naturales de las principales regiones geomorfológicas de Uruguay. In: Altesor A, Ayala W, Paruelo JM (eds) Bases ecológicas y tecnológicas para el manejo de pastizales. INIA, Montevideo, pp 15–32
MacArthur RH, MacArthur JW (1961) On bird species diversity. Ecology 42:594–598. https://doi.org/10.2307/1932254
Milligan MC, Berkeley LI, McNew LB (2020) Effects of rangeland management on the nesting ecology of sharp-tailed grouse. Rangel Ecol Manag 73:128–137. https://doi.org/10.1016/j.rama.2019.08.009
Modernel P, Rossing WAH, Corbeels M, Dogliotti S, Picasso V, Tittonell P (2016) Land use change and ecosystem service provision in Pampas and Campos grasslands of southern South America. Environ Res Lett 11:113002. https://doi.org/10.1088/1748-9326/11/11/113002
Murray C, Minderman J, Allison J, Calladine J (2016) Vegetation structure influences foraging decisions in a declining grassland bird: the importance of fine-scale habitat and grazing regime. Bird Stud 63:223–232. https://doi.org/10.1080/00063657.2016.1180342
Nakagawa S, Cuthill IC (2007) Effect size, confidence interval and statistical significance: a practical guide for biologists. Biol Rev 82:591–605. https://doi.org/10.1111/j.1469-185X.2007.00027.x
Neilly H, O’Reagain P, Vanderwal J, Schwarzkopf L (2018) Profitable and sustainable cattle grazing strategies support reptiles in tropical savanna rangeland. Rangel Ecol Manag 71:205–212. https://doi.org/10.1016/j.rama.2017.09.005
Norment CJ, Ardizzone CD, Hartman K (1999) Habitat relations and breeding biology of grassland birds in Western New York: management implications. Stud Avian Biol 19:112–121
Pipher EN, Curry CM, Koper N (2016) Cattle grazing intensity and duration have varied effects on songbird nest survival in mixed-grass prairies. Rangel Ecol Manag 69:437–443. https://doi.org/10.1016/j.rama.2016.07.001
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 26 Oct 2022
Robel RJ, Briggs JN, Dayton AD, Hulbert LC (1970) Relationships between visual obstruction measurements and weight of grassland vegetation. Rangel Ecol Manag 23:295–297. https://doi.org/10.2307/3896225
Rodríguez C, Leoni E, Lezama F, Altesor A (2003) Temporal trends in species composition and plant traits in natural grasslands of Uruguay. J Veg Sci 14:433–440. https://doi.org/10.1111/j.1654-1103.2003.tb02169.x
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113. https://doi.org/10.1111/j.2041-210X.2010.00012.x
Shannon CE (1948) A mathematical theory of communication. Bell Syst Techn J 27:379–423. https://doi.org/10.1002/j.1538-7305.1948.tb01338.x
Sliwinski M, Powell L, Schacht W (2019) Grazing systems do not affect bird habitat on a sandhills landscape. Rangel Ecol Manag 72:136–144. https://doi.org/10.1016/j.rama.2018.07.006
Sliwinski MS, Powell LA, Schacht WH (2020) Similar bird communities across grazing systems in the Nebraska Sandhills. J Wildl Manag 84:802–812. https://doi.org/10.1002/jwmg.21825
Soriano A (1992) Río de la Plata grasslands. Ecosyst World 8:367–407
Vickery PD, Tubaro PL, Silva JMC et al (1999) Conservation of grassland birds in the western hemisphere. Stud Avian Biol 19:2–26
Acknowledgements
This work was supported by the International Program, U.S. Forest Service, Alianza del Pastizal-BirdLife International, Aves Uruguay, and Centro Universitario Regional del Este, Universidad de la República, Uruguay.
Funding
International Programs, US Forest Service; Alianza del Pastizal-BirdLife International; Aves Uruguay.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by: Carla Fontana, Ph D. (Associated Editor)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Pírez, F., Aldabe, J. Comparison of the bird community in livestock farms with continuous and rotational grazing in eastern Uruguay. Ornithol. Res. 31, 41–50 (2023). https://doi.org/10.1007/s43388-022-00113-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43388-022-00113-1