
Abstract
Several studies on the social behavior of birds have shown two main benefits of social groups: optimization of foraging and decreasing the risk of predation. We tested whether the association between rufous hornero (Furnarius rufus) and the chalk-browed mockingbird (Mimus saturninus) affects the foraging and alert/sentinel behaviors of both species. We also evaluated the influence of group size on these behaviors. During 60 h of observations on each species, the association was studied in a periurban area in Brazil with scan samplings. The presence of F. rufus increased the frequency of foraging while reducing the time that M. saturninus dedicated to alert/sentinel behaviors. For F. rufus, however, the presence of M. saturninus did not affect the studied behaviors, although a different study pointed out that this species could also benefit from the association with M. saturninus, indicating that both species may occasionally be positively affected by the association with each other.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Several studies on the social behavior of birds have shown two main benefits of social groups: optimization of foraging by scaring prey away, which minimizes the foraging effort by facilitating the location and capture of prey (Morse 1977), and decreasing the risk of predation (Morrell and James 2007) due to the confusion or dilution effect (Lehtonen and Jaatien 2016), selfish-herd effect (i.e., the protection accrued by individuals foraging at the center of the group; Dias 2006), rapid dissemination of information (e.g., Amorim and Dias 2021), and anticipated detection of predator resulting from shared vigilance (Dias 2006). On the other hand, the spread of diseases/parasites (Brown and Brown 2004) and the increased competition for resources are among the costs associated with group living (Beauchamp 2010). For both conspecific and heterospecific groups, these costs and benefits may be present.
In the case of heterospecific groups, most studies in the Neotropical region involve mixed species flocks (e.g., Alves and Cavalcanti 1996; Ragusa-Netto 2002; Tubelis 2006; Reis 2011). Associations involving only two species, unlike the typical mixed flocks, are still poorly studied, even when they involve common and widely distributed birds. For instance, the black-and-white monjita (Xolmis dominicanus) and the saffron-cowled blackbird (Xanthopsar flavus), although recorded in mixed flocks in the southeastern South American grasslands (Fraga et al. 1988; Fraga 2003; Fraga 2005), frequently associate separately as one species takes advantage of the sentinel behavior of the other (Dias and Maurício 2002; Krüger and Petry 2010).
The chalk-browed mockingbird (Mimus saturninus) lives in flocks of 6 to 12 individuals, always with the presence of a sentinel, who occupies an elevated position while other individuals of the group forage (Sigrist 2005). It is an omnivorous species that feeds mainly on the ground (Argel-de-Oliveira 1989) and inhabits almost all of Brazil, except forested regions of the Amazon (Sick 1997). According to Sigrist (2005), M. saturninus can forage alongside other birds of open areas such as guira cuckoo (Guira guira), smooth-billed ani (Crotophaga ani), white woodpecker (Melanerpes candidus), rufous hornero (Furnarius rufus), and campo flicker (Colaptes campestris). Furnarius rufus, well known for its oven-shaped clay nests, is almost always seen in pairs and has a predominantly terrestrial habit, foraging on the ground, turning over leaves and scavenging fallen trunks, sometimes followed by other ground-foraging species, such as the C. campestris and M. saturninus (Sigrist 2005). According to Sabino (2015), the average home range size of F. rufus is 0.74 ± 0.48 ha. Information on the home range size of M. saturninus is not available but is likely larger than F. rufus. Both species, M. saturninus and F. rufus, are among the most common urban birds in Brazil (Sick 1997; Sigrist 2005).
Argel-de-Oliveira (1989) provided the first evidence of an association between the two species, noting that F. rufus was the bird most often recorded close to M. saturninus during foraging and roosting. Ragusa-Netto (1997) carried out the first work focused on this association, showing that individuals of F. rufus had higher foraging efficiency when associated with M. saturninus. However, there is no information quantifying the effects of this association for M. saturninus. Thus, the objective of this study was to test if this association affects both species. We hypothesized that the association would increase the foraging frequency for both species, while decreasing the alert/vigilance frequency, expressed by the number of individuals engaged in each behavior in instantaneous samplings. In addition, we hypothesized that group size also had a similar effect, allowing for a lower alert/vigilance frequency and a higher foraging frequency.
Materials and methods
Study area
The study was carried out in the campus of the Universidade Estadual Paulista “Júlio de Mesquita Filho” (22°23′45.7″S; 47°32′38.3″W), a 111.46-ha area located at 626.5 m a.s.l. in the municipality of Rio Claro, São Paulo, Brazil (Carvalho 2020). The campus is a typical periurban area composed by vegetation resulting from a landscaping project or established by natural dispersal, with little of the original semidecidual forest (Potascheff et al. 2010).
Data collection
We conducted 120 h of focal observations of M. saturninus and F. rufus (60 h for each species) from April to July 2019, which corresponds to the non-breeding season for both species. Observations were performed with binoculars in 1-h sessions (two to eight sessions a day) between 7:00 and 11:00 h, and from 14:00 and 18:00 h. The order of the groups to be observed was randomly drawn. Each group was observed only once a day, and if the focal group moved out of sight before the end of a given observation session, that particular session was cancelled and repeated the next day.
We observed five groups of M. saturninus, totalling 30 individuals (group size: 6.2 ± 2.4 individuals per group; mean ± standard deviation), 14 of which were marked with metal and colored rings (Supplementary material Table S1). Based on the location of these groups, five pairs or familiar groups (2.4 ± 0.5 individuals per group) of F. rufus were selected for sampling, totalling 12 individuals, five of which were individually marked with metal and colored rings (Supplementary Table S1). Birds were mist-netted and marked according to the Brazilian regulations (SISBIO license n° 45434-5, CEMAVE authorization n° 3362, CEUA permit n° 030/2012).
At each observation session, individuals were followed at a minimum distance of 10 m and maximum of 40 m (adapted from Argel-de-Oliveira 1989), taking into consideration that the birds in the campus were habituated to human proximity. Scan samples (Altmann 1974) were taken every 5 min for which the following behaviors were recorded for all individuals of the group: alert/sentinel (individuals who stay on high and exposed perches, interrupting foraging activities to look around for potential predators; adapted from Argel-de-Oliveira 1989), and foraging. At each scan, the number of individuals engaging in alert/sentinel or foraging behavior was divided by the number of individuals observed at that scan sample, resulting in the relative frequency of occurrence of each behavior in a given scan. At each scan, we also recorded if the observed species was associated with the other species, i.e., if individuals of the observed group were less than 10 m apart from the individuals of the other species.
Statistical analyses
We tested whether the number of scans in which each species was associated to the other differed between species using a chi-square test, using the number of scans with or without heterospecific association as a frequency count for each species.
To test if a particular group was more inclined to perform alert/vigilance or foraging behaviors, we used generalized linear models (GLMs) with binomial error distribution, setting the relative frequency of each behavior as response variables and group ID as the explanatory variable. As there was no difference in the frequencies of each behavior between the groups of each species, we pooled the data from all groups of each species in the following analyses.
We evaluated whether the frequency of certain behaviors was affected by heterospecific association by creating generalized linear mixed models (GLMMs) with binomial distribution of errors, with one model for each behavior category for each species. The response variable for each of the models was the relative frequency of a given behavior in each of the scans, as described above. As independent variables, we used group size that refers to a constant value for each group that represents the maximum number of individuals observed in each group during the study, and the presence of heterospecific association as a binary variable. Since the number of F. rufus individuals associated with M. saturninus groups was greater than 3 in only 4.3% of scans, these occurrences were pooled in a single category of 4 or more individuals associated. When the heterospecific association affected the relative frequency of a particular behavior, we performed a new GLMM to test whether the frequency of that behavior (dependent variable) also depended on the number of heterospecific individuals (continuous independent variable) associated with the focal species. The focal group identity was included as a random factor in all models. GLMs and GLMMs were built in R software (R Core Team 2019), using the packages lme4 (Bates et al. 2015) and lmerTest (Kuznetsova et al. 2017).
Results
We obtained 258 scans for five groups of M. saturninus (51.6 ± 14.9 scans per group) and 160 scans for four groups of F. rufus (40.0 ± 28.2 scans per group). There was no shift of marked individuals of either species between the groups studied, and each group of M. saturninus always associated with the same individuals of F. rufus. In 57% (n = 258) of the scans focused on M. saturninus, they were associated with F. rufus, while in 42.5% (n = 160) of the scans in F. rufus, they were associated with M. saturninus. The number of scans where each species was associated with the other deviated from that expected by chance (χ2 = 8.286, P = 0.004), indicating that M. saturninus was significantly more frequently associated with F. rufus than the contrary.
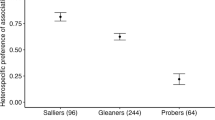
The presence of F. rufus affected the frequency of foraging and alert/sentinel behaviors performed by M. saturninus (Table 1). In the presence of F. rufus, M. saturninus increased the frequency of foraging, which nearly doubled from 21.8 to 42% (Fig. 1). Mimus saturninus also spent less time on alert/sentinel in the presence of F. rufus, with the frequency of this behavior dropping by half from 38.2 to 19.5%.

Influence of the presence of the rufous hornero (Furnarius rufus) on foraging and alert/sentinel behaviors in the chalk-browed mockingbird (Mimus saturninus). Numbers inside the bars correspond to the number of scans of the displayed data
There was an effect of M. saturninus group size on foraging frequency, which tended to increase with increasing group size (Table 1). Although the presence of F. rufus influenced foraging and alert/sentinel behaviors in M. saturninus, the number of F. rufus individuals did not influence (P = 0.386 and P = 0.465) the two behaviors.
Contrary to M. saturninus, the association did not influence the frequency of F. rufus behaviors, although the effect of the heterospecific grouping on the foraging of F. rufus was marginally significant (Table 1). The size of the M. saturninus group had no influence on the foraging and alert/sentinel behaviors of F. rufus (Table 1).
Discussion
When the association between M. saturninus and F. rufus was firstly described, it was hypothesized that F. rufus was the species that actively associated with M. saturninus, but no quantitative data regarding this association was provided (Argel-de-Oliveira 1989). We found, however, a greater preference for M. saturninus to associate with F. rufus (57% for M. saturninus and 42.5% for F. rufus). Given that M. saturninus is apparently more affected by the association, it is reasonable that it seeks F. rufus more often than the contrary. This is in line with the larger home range that M. saturninus likely has in relation to F. rufus (~2 ha; Sabino 2015; Amorim 2020) meaning that while a group of M. saturninus may find different pairs of familiar groups of F. rufus in their home ranges, the reverse is not true.
We provided evidence that the association between M. saturninus and F. rufus is potentially beneficial to the former but not to the latter species, which apparently differs from the results obtained by Ragusa-Netto (1997) showing a benefit also to F. rufus. It is important to note, however, that the two studies differ in the response evaluated. While we recorded the frequency of foraging as denoted by the proportion of individuals engaged in foraging, Ragusa-Netto (1997) focused on the rate of pecking during foraging. Moreover, the method used by Ragusa-Netto differs from ours in the following aspects: only one species was followed (F. rufus) during observations, which lasted from 1 to 2 min per individual and were performed in sequence; only pecking events on the substrate were recorded as foraging; for the analyses, each observation of an isolated bird was taken as a replicate, while for birds in groups, the average pecking rate for each group was used. Such differences in methodology may thus preclude a direct comparison with the present study. Anyway, the fact that we detected a marginally significant influence of the presence of M. saturninus on the foraging of F. rufus is suggestive that an increase in the foraging frequency of the former species is not unlikely.
The greater the number of individuals of M. saturninus in a group, the greater the amount of time they dedicated to foraging. Several studies on the effect of group size on feeding rate showed that this positive relationship results from the reduced vulnerability to predation provided by larger groups (see review by Beauchamp 1998). On the other hand, the food found needs to be shared, which may increase aggression and decrease the average food intake rate (Maruyama et al. 2009), a possibility that we did not investigate. Anyway, we recorded no agonistic encounters between the two species, which may indicate that there was not a fierce dispute over resources.
The increase in group size of M. saturninus did not affect the proportion of individuals engaged in alert/sentinel behaviors, contradicting several studies on ground-foraging birds that report an inverse relationship between these aspects (Sridhar et al. 2009). For F. rufus, group size was not related to any of the behaviors studied. One likely reason is that this species is territorial and non-gregarious, living mostly in pairs (Sick 1997), which suggests that they are unable to benefit from the proximity of conspecifics due to the several costs offered by unrelated conspecifics, for example, possibility of extra-pair copulation, usurpation of food and territory (Fraga 1980). It seems that for F. rufus the social behavior benefits could be achieved by associating with heterospecifics.
More studies could be carried out focusing on other behavioral aspects involved in the association, such as perching behavior, since apparently these two species choose to sleep in the same set of trees that we and Argel-de-Oliveira (1989) observed, indicating that the association goes beyond the realm of collecting. Also, as reproductive demands interfere with foraging time, reproductive aspects must be investigated. For example, M. saturninus relies on the cooperation of other individuals in the herd to feed the young (Argel-de-Oliveira 1989), which can provide more time for foraging, while F. rufus divides parental care into just two individuals (Fraga 1980), which can reduce the time available for foraging.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–267. https://doi.org/10.1163/156853974x00534
Alves MAS, Cavalcanti RB (1996) Sentinel behavior, seasonality, and the structure of bird flocks in a Brazilian savanna. Ornitol Neotrop 7:43–51
Amorim PSP (2020) Traços de vida e o efeito do querido inimigo em joão-de-barro (Furnariidae: Furnarius rufus). MSc. Dissertation, Universidade Federal de Juiz de Fora
Amorim PS, Dias RI (2021) A non-vocal alarm? Effects of wing trill playbacks on antipredator responses in the scaled dove. Acta Ethol 24:119–126. https://doi.org/10.1007/s10211-021-00368-9
Argel-de-Oliveira MM (1989) Ecoetologia do sabiá-do-campo Mimus saturninus no estado de São Paulo. MSc. Dissertation, Universidade Estadual de Campinas
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Beauchamp G (1998) The effect of group size on mean food intake rate in birds. Biol Rev 73:449–472. https://doi.org/10.1111/j.1469-185X.1998.tb00179.x
Beauchamp G (2010) A comparative analysis of vigilance in birds. Evol Ecol 24:1267–1276. https://doi.org/10.1007/s10682-010-9358-5
Brown CR, Brown MB (2004) Group size and ectoparasitism affect daily survival probability in a colonial bird. Behav Ecol Sociobiol 56:498–511. https://doi.org/10.1007/s00265-004-0813-6
Carvalho REWF (2020) Biologia reprodutiva do gaviãozinho Gampsonyx swainsonii (Accipitridae, Aves). Undergraduate Monograph, Universidade Estadual Paulista “Júlio de Mesquita Filho”
Dias RI (2006) Effects of position and flock size on vigilance and foraging behaviour of the scaled dove Columbina squammata. Behav Process 73:248–252. https://doi.org/10.1016/j.beproc.2006.06.002
Dias RA, Maurício GN (2002) Natural history notes and conservation of a saffron-cowled blackbird Xanthopsar flavus population in the southern coastal plain of Rio Grande do Sul, Brazil. Bird Conserv Int 12:255–268. https://doi.org/10.1017/S0959270902002162
Fraga RM (1980) The breeding of rufous horneros (Furnarius rufus). Condor 82:58–68. https://doi.org/10.2307/1366785
Fraga RM (2003) Distribution, natural history and conservation of the black-and-white monjita (Heteroxolmis dominicana) in Argentina, a species vulnerable to extinction. Ornitol Neotrop 14:145–156
Fraga RM (2005) Ecology, behavior and social organization of saffron-cowled blackbirds (Xanthopsar flavus). Ornitol Neotrop 16:15–29
Fraga RM, Casañas H, Pugnali G (1988) Natural history and conservation of the endangered saffron-cowled blackbird Xanthopsar flavus in Argentina. Bird Conserv Int 8:255–267. https://doi.org/10.1017/S095927090000191X
Krüger L, Petry MV (2010) Black-and-white monjita (Xolmis dominicanus) followed by the saffron-cowled blackbird (Xanthopsar flavus): statistical evidence. Ornitol Neotrop 21:299–303
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 82:1–26. https://doi.org/10.18637/jss.v082.i13
Lehtonen J, Jaatien K (2016) Safety in numbers: the dilution effect and other drivers of group life in the face of danger. Behav Ecol Sociobiol 70:449–458. https://doi.org/10.1007/s00265-016-2075-5
Maruyama PK, Cunha AF, Tizo-Pedrosos E, Del-Claro K (2009) Relation of group size and daily activity patterns to southern lapwing (Vanellus chilensis) behaviour. J Ethol 28:339–344. https://doi.org/10.1007/s10164-009-0193-5
Morrell LJ, James R (2007) Mechanisms for aggregation in animals: rule success depends on ecological variables. Behav Ecol 19:193–201. https://doi.org/10.1093/beheco/arm122
Morse DH (1977) Feeding behavior and predator avoidance in heterospecific groups. Biosci 27:332–339. https://doi.org/10.2307/1297632
Potascheff CM, Lombardi JA, Lorenzi H (2010) Angiospermas arbóreas e arbustivas do campus da Universidade Estadual Paulista Júlio de Mesquita, Rio Claro (SP). Bioikos 24:21–30
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ragusa-Netto J (1997) Evidence for the possible advantage of heterospecific social foraging in Furnarius rufus (Passeriformes: Furnariidae). Ararajuba 5:233–235
Ragusa-Netto J (2002) Vigilance towards raptors by nuclear species in bird mixed flocks in a Brazilian savannah. Stud Neotrop Fauna Environ 37:219–226. https://doi.org/10.1076/snfe.37.3.219.8573
Reis MG (2011) Ecologia de bandos mistos de aves na Estação Ecológica de Itirapina, estado de São Paulo. MSc. Dissertation, Universidade Federal de São Carlos
Sabino U (2015) Área de vida e comportamento de cuidado parental do Furnarius rufus (joão-de-barro). MSc. Dissertation, Universidade Federal do Mato Grosso
Sick H (1997) Ornitologia brasileira. Nova Fronteira, Rio de Janeiro
Sigrist T (2005) Aves do Brasil. Avis Brasilis, Vinhedo
Sridhar H, Beauchamp G, Shanker K (2009) Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim Behav 78:337–347. https://doi.org/10.1016/j.anbehav.2009.05.008
Tubelis DP (2006) Role of mixed-species flocks in the use of adjacent savannas by forest birds in the central Cerrado, Brazil. Austral Ecol 31:38–45. https://doi.org/10.1111/J.1442-9993.2006.01541.X
Acknowledgements
We thank Karl Mokross for his collaboration in the conception of the study and for his help in the field, colleagues Antônio Fontanella, André Luiz Bitencourt Moraes, and Ivan Provinciato for their help in the field, and two anonymous reviewers for their comments and suggestions on the manuscript. We also thank the Department of Biodiversity for the logistic support.
Funding
MAP receives a research fellowship from the Brazilian Research Council (CNPq #304742/2019-8).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by: Cristiano Azevedo
Supplementary Information
ESM 1
(PDF 78.8 kb)
Rights and permissions
About this article

Cite this article
Piassi, P.M., Batisteli, A.F., Gussoni, C.O.A. et al. Effects of the association between Mimus saturninus and Furnarius rufus on their foraging and alert behaviors. Ornithol. Res. 30, 99–103 (2022). https://doi.org/10.1007/s43388-022-00090-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43388-022-00090-5