
Abstract
Human settlement and agricultural activities restrict increasingly the range of large mammals in many cases contributing to declining numbers of ungulates. Here, we studied home range size and habitat selection of seven adult female mountain nyalas in the northern end of the Bale Mountains National Park (BMNP) equipped with Global Positioning System (GPS) collars. Home range size was estimated using fixed kernel density and habitat selection was determined by resource selection functions. We found that female mountain nyalas have much smaller (5.7 km2) home ranges than the 19 km2 home range size predicted for a 170 kg, group-living species living in mixed habitats. Home ranges were 30% larger in the night than in the daytime. We suggest that the night extension beyond the park boundaries was caused by high livestock and other ungulates grazing pressure within the protected area which may cause forage-driven excursions out of the park. On the other hand, mountain nyalas are probably attracted by humans as shields against hyena predation. The resource selection index indicates that bushland and forest habitat are the most preferred habitat types, while agriculture and human settlements are the least preferred habitats. The small size of the northern part of the Bale Mountain National Park (31 km2) with the current high density of mountain nyala (24 individuals/km2) is too small for the predicted home range size of large ungulates. Hence, we suggest that protecting additional area may be needed for the long-term conservation of the endangered mountain nyala.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Home ranges and activity pattern of animals can be affected by several factors, such as the availability and quality of food and other resources, population densities, the presence of competitors, risk of predation, and climate (Willems and Hill 2009; Ochiai et al. 2010; Van Beest et al. 2011). Increasingly, human activities and human-caused landscape alteration and fragmentation also affect animal home range size, habitat use, activity, and movement patterns (Tucker et al. 2018; Knüsel et al. 2019).
As a rule of thumb, the size of a home range is expected to increase with the body size of the respective species (Noonan et al. 2020), mediated also by habitat quality and type (e.g., open, mixed, closed habitat) and the species’ social organization (e.g., solitary or group living: Mysterud et al. 2001; Ofstad et al. 2016). The influence of humans is expected to be higher for larger mammals because of their higher space requirements (Tucker et al. 2014). Furthermore, since humans are mainly diurnal, they interfere with wildlife more in the daytime than during the night, pushing many mammal species toward a more nocturnal activity (Gaynor et al. 2018).
A greater understanding of the human impact on activity and movement patterns, home ranges, and habitat use constitute important baseline data for conservation management, in particular the design of protected areas (Rhodes et al. 2005; Harless et al. 2009; Rechetelo et al. 2016). This is of particular importance for endangered species whose populations and ranges have already been reduced (Rechetelo et al. 2016).
Mountain nyalas, Tragelaphus buxtoni, are endangered antelopes endemic to the south-eastern highlands of Ethiopia. The largest mountain nyala population (n = 1100) still exists in the forest-dominated landscape of the Bale Mountains National Park (BMNP) (Atickem et al. 2011). The park was established in the 1970s primarily to protect mountain nyalas and another endemic species, for example the Ethiopian wolf, Canis simensis (Waltermire 1975). Currently, about 90% of the mountain nyala population of the park is confined to the park’s northern part, the Gaysay Area (Atickem et al. 2011). Human settlement and livestock grazing, however, are increasing outside and inside the park, resulting in competition for space and food with wild large herbivores, including the mountain nyala (Atickem and Loe 2013).
In this study, we equipped seven female mountain nyalas with GPS collars to investigate their activity and movement pattern, home range size, and habitat use. Given the strong human encroachment in the area surrounding the Gasay Area, we predict that the home ranges of the mountain nyalas are smaller than expected by theory and that diurnal habitat selection and movement patterns are influenced by human infrastructure. Based on our findings, we discuss whether the current size of the protected area and conservation measures are sufficient for the long-term conservation of mountain nyala.
Materials and methods
Study area
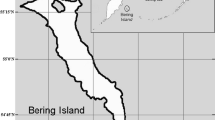
The study was carried out in the Gaysay Area (31 km2) of the BMNP and its surroundings outside of the national park. Unlike large parts of the BMNP, which are dominated by human settlements and herds of livestock, the Gaysay Area is patrolled by rangers as protection from livestock grazing, but the scale of illegal grazing is still extensive (Atickem and Loe 2013). The altitude of the area ranges from 3000 m to 3600 m, and the climate is characterized by a 4-month dry season (November to February) and an 8-month rainy season (March to October). We used spatial data from 2008, because the threat situation for the nyalas and the degree of human activities did not further degrade over the last 16 years due to the permanent presence of park rangers in the area. We defined our study area as the 100% minimum convex polygon containing all locations (fixes) of all collared female mountain nyalas (number fixes = 31,649), corresponding to an area of about 133 km2 (Fig. 1).

Study area in the northern part of the Bale mountains national park
Mountain nyala GPS-data
Since our budget allowed to buy only a limited number of collars, we decided to equip females with collars, since information on their spatial behavior is more relevant for the management and survival of the population. We equipped seven female mountain nyalas with Global Positioning System (GPS) collars (Followit Holding AB, Sweden) between 25/05/2008 and 29/05/2008. The females were immobilized using a remote injection system and collared (Atickem et al. 2016). The GPS units were programmed to take fixes every other hour, i.e., 12 fixes per day. We downloaded data from the GPS collars remotely by an external telemetry receiver and transferred respective location data onto a computer with a Tellus Project Manager. The collars lasted on average 402 days before failure (SD = 215, range 212–681 days). The immobilization and handling of the animals were permitted by the Ethiopian Wildlife Conservation Authority and followed their Ethical guidelines (Atickem et al. 2016). Our original plan was to immobilize the collared mountain nyala after our study to remove the collars. However, the experts at the Ethiopian Wildlife Conservation Authority decided that the collars cause less disturbance to the animal than capturing them again. Therefore, we were asked not to remove the collars.
Home range estimation
We conducted home range analyses using fixed kernel density estimation. The bandwidth h was determined with the rule-based ad hoc approach (Kie 2013). Home range contours were calculated at the 95% isopleth level. The most intensively used portion of a home range was regarded as the core area, i.e., the area with the 50% isopleth level (Bingham et al. 1997; Seaman et al. 1999). In addition, we calculated minimum convex polygons (MCP) at the 100% isopleth level for the habitat utilization analysis. Home ranges and core areas were estimated with the R package “rhr” (Signer and Balkenhol 2015).
Predicted home range size
Female mountain nyalas weigh between 150 and 200 kg, and live in groups with an average of nine animals in mixed habitats (Sillero-Zubiri 2013). To calculate their predicted home range size, we applied the equation given by Ofstad et al. (2016) HR = exp(− 10.28 + 1.44*log(BM) + 0.3*log(group size))/100, where HR refers to home range size, BM to body mass (measured in grams, here an average of 170,000 g), and group size (number of individuals, here 9). Note that the constant (− 10.28) is specific for species living in mixed habitats. Accordingly, for a female mountain nyala, the predicted HR size would be 19 km2.
Habitat classification
We defined six habitat classes: grassland, forest, heath (dominated by Erica sp.), bushland, agriculture, and human settlement. We classified these habitats from SPOT images (2 m resolution) by a supervised classification (ERDAS Imagine) based on 220 ground-truthing locations (ERDAS 1994; Dean and Smith 2003). Elevation and slope values were derived from a 90 m Digital Elevation Model (DEM; Jarvis et al. 2008). Geographic positions of settlements adjacent to the Gaysay Area were determined with handheld GPS (Atickem et al. 2014, for more details on habitat classification).
Habitat preference
To infer habitat preferences of the female mountain nyalas within our study area, we applied Generalized Linear Models (GLMs) with a use-availability design in R (Guisan and Zimmermann 2000; Johnson et al. 2008; R Development Core Team 2011). We used the animal locations as the sample unit in all analyses. Each location where a female was actually recorded (used location) was paired with one random (available) location in the following way: the random location copied all the non-spatial variables from the used point (female ID, season, daytime/night) before adding new spatial covariates (distance to humans, habitat class) from the random location (van Beest et al. 2010). The response variable was binomially distributed and consisted of real points (GPS points of respective mountain nyalas, coded 1) and randomly sampled points (coded 0).
To account for temporal autocorrelation among observations, we estimated robust standard errors using the approach of Forester et al. (2009), i.e., we clustered the data based on the lag of the temporal autocorrelation and placed every other cluster in a second data set. The robust standard errors were calculated by averaging the covariance matrices for both subsets of data, while the parameter estimates represent fitted values from the full-data set.
Sample sizes differed among individuals. To prevent sample size bias, we weighted the contribution of each data point by a vector W = the lowest sample size across all individuals/sample size of the focal animal in the GLM using the weights argument. Parameter estimates of this analysis are log odds of use relative to reference categories of categorical variables and zero values of continuous variables.
Results
Home range size
A total of 31,649 GPS fixes (Table 1) were obtained from the seven females with an average of 4,521 GPS fixes [SD = 2368] per female. The GPS collars recorded fixes with an overall success rate of 92.1% (range between 82.8% and 96.3%). On average, females were significantly more often located outside the protected area during nights (30% ± 33%) than during daytime (13% ± 18%, V = 2, n = 7, p = 0.021). However, there was a large inter-individual variation (2.3 to 99.4%; Table 1).
The resulting average home range size of a female was 5.7 ± 4.4 km2 and the mean core area size was 1.1 ± 0.79 km2 (Table 2). This is only about 1/3 of the 19 km2 home range size predicted for the females from the Ofstad et al.’s (2016) equation. On average, 70% (SE = 30%, range: 22–100%, Table 2, Fig. 1) of the home range of the females was located inside the protected area, but there were large individual differences.
Habitat use and habitat selection
The Gaysay Area consisted of grassland (45.5%), forest (22.9%), bushland (19.4%), heath (5.5%), and others including water bodies, roads, and settlements (6.8%). The area surrounding the Gaysay Area (buffer zone) was dominated by human settlement and crop fields.
Habitat selection of the female mountain nyalas differed between seasons and between night and day for some, but not all, habitat types. Heath was avoided both day and night in both seasons. Bushland and grassland were preferred daytime habitats, but were avoided during the night (except bushland in the wet season). Forest was strongly preferred during day time and moderately preferred during the night in the dry season, while the preference of forest was neutral both in day and night in the wet season (Fig. 2, Table 3). All seven females avoided human settlements and agriculture in the day time.

Habitat use of mountain nyala during day and night time
Discussion
The Gaysay Area at the northern end of the BMNP has been the stronghold of mountain nyalas for over 4 decades and is home to 90% of the mountain nyala population in the park. The Gaysay Area is, however, small in size (31 km2) with many herbivore wildlife species and is under livestock grazing pressure (Atickem and Loe 2013). With this study, we demonstrated that the home range size of the female mountain nyalas is by far less than the home range size predicted for group-living species living in mixed habitats. Bushland and forest are the most preferred habitat types by female mountain nyalas and these habitats combined are much smaller (13 km2) suggesting the need for an extension of the protected area free from the impact of humans and livestock grazing. Hence, we recommend extending the protection of the Gaysay Area habitat against human settlement and livestock grazing beyond the current 31 km2 area toward the southern range of the mountains. During 1986, this area was used intensively by mountain nyalas and used as a corridor to the eastern escarpments of the Bale Mountains (Afework et al. 2010). Since males roam farther and have usually larger home ranges than females (Hillman 1986; Sillero-Zubiri 2013), an extension of the Gaysay Area will most likely also benefit the males.
The mean 95% of the home range estimate (5.3 km2) was only 28% of the home range expected for a 170 kg, group-living species living in a mixed habitat. The comparatively small and largely overlapping home ranges of female mountain nyalas may be due to the dense human population surrounding the park. It is reported that human-dominated landscapes surrounded by a matrix exploited by agriculture or human settlement limit the ranging pattern and adversely affect the fitness of wildlife (Alfred et al. 2012; Knüsel et al. 2019). There is reason to expect a large cumulate impact on the suite of large herbivores in the Gaysay Area; in addition to the mountain nyala population which is estimated to be about 840 individuals (Atickem et al. 2011). The Gaysay Area is home to many other herbivores including about 400 Bohor reedbucks Redunca redunca (Afework et al. 2010), ca 600 common warthogs Phacochoerus africanus (Deribe et al. 2008), and an unknown number of Menelik’s bushbucks Tragelaphus scriptus and gray duikers Sylvicapra grimmia. Also, there is extensive illegal grazing of domestic cattle (Atickem and Loe 2013). The carrying capacity of the Gaysay Area may limit further population growth of the herbivore community including mountain nyalas.
Protected areas are the primary biodiversity conservation strategy used across the globe to avert biodiversity loss (Chape et al. 2005). The effectiveness of the protected areas size, however, becomes controversial as wildlife populations decline from several protected areas (Tucker et al. 2014; Butchart et al. 2010; Scholte 2011; Ogutu et al. 2016). Noonan et al. (2020) suggest the home range size of mammals on the bases of their body mass very much under estimated the home range size which possibly contributes for the sharp decline of large mammals. The decline is in particular severe for large mammals where Craigie et al. 2010 reported a 59% decline in large mammal population abundance in Africa’s Protected Areas in less than 40 years. Understanding the ecological requirement of large mammals and available resources in the protected area is key for reversing the currently observed decline of wildlife species (Gaston et al. 2008; Cattarino et al. 2016). During the night, mountain nyalas spend a relatively higher proportion of their time outside the park than in the daytime. During the wet season, mountain nyalas may move out of the park for foraging on the barley crop, but they also move toward human settlements to avoid hyena predation (Atickem et al. 2014), known as the human shield strategy (Berger 2007). While mountain nyalas are very shy in much of their range in the eastern escarpments of the Bale Mountains where legal and illegal hunting may occur, they are more tolerant of human presence in the protected Gaysay Area (Atickem et al. 2014). Therefore, range contraction due to human infrastructure may be expected to be stronger than in other areas inhabited by mountain nyalas.
For large herbivores, habitat selection is often a trade-off between foraging and avoidance of predators (Godvik et al. 2009). Many prey species resolve this by selecting predator-safe habitat during the time the predator is active and foraging habitat, while the predator is inactive (Lone et al. 2017). The mountain nyalas moved out of the park, in particular during the dry season when no crop for forage is available, to avoid hyena predation (Atickem et al. 2014).
The resource selection index indicates that bushland and forest habitat are the most preferred habitat types for mountain nyalas, while agriculture and human settlements are the least preferred habitats. Heath bushland, one of the most important habitats for mountain nyalas (Brown et al. 2012), is part of the Gaysay Area and was found to be among the least used and least preferred habitats during this study. From the geographic scale resource selection of mountain nyalas (Atickem et al. 2011), forest areas were found to be the most preferred habitat with the largest mountain nyala population, while mountain nyalas avoid human-influenced areas (agriculture and human settlement). The mountain nyala population of the Gaysay Area may be established due to its protected status rather than its habitat quality for forage in the first place. Mountain nyalas were not reported in the Gaysay Area during a survey by Brown in 1969 (Brown et al. 2012). Following the establishment of the park in 1970, the mountain nyala population dramatically increased from few individuals (Malcolm and Evangelista 2004) to 1100 in 1986 (Afework et al. 2010). While the population may have increased through births, the increase likely represents high emigration from the surrounding mountains due to pressure from the human population and attraction to the protected status of the area. Since 1991, humans and their livestock have encroached upon BMNP leaving the Gaysay Area as the last stand refuge for the mountain nyala population (Stephens et al. 2001).
Conclusion
Our study suggests that the currently protected area of the northern end of the Bale Mountains National Park, Gaysay area, is too small for the mountain nyala conservation. Given a large number of other herbivores and continued livestock grazing, the future long-term conservation of the endangered mountain nyala needs an additional protected area free from human activities and livestock grazing.
Availability of data and materials
Data are available at Dryad, Dataset, https://doi.org/10.5061/dryad.fttdz08pv.
Code availability
Not applicable.
References
Afework B, Bekele A, Balakrishnan M (2010) Population status, structure and activity patterns of the bohor reedbuck Redunca redunca in the north of the Bale Mountains National Park, Ethiopia. Afr J Ecol 48:502–510. https://doi.org/10.1111/j.1365-2028.2009.01139.x
Alfred R, Ahmad AH, Payne J, Williams C, Ambu LN, How PM, Goossens B (2012) Home range and ranging behaviour of Bornean elephant (Elephas maximus borneensis) females. PLoS ONE 7:e31400. https://doi.org/10.1371/journal.pone.0031400
Atickem A, Cooper DV, Kock R, Rueness EK, Fischer M, Bekele A, Loe LE, Stenseth NC (2016) Immobilization of mountain nyala (Tragelaphus buxtoni) in the Bale Mountains. Ethiopia Gnusletter 33:6–7
Atickem A, Loe LE (2013) Livestock-wildlife conflicts in the Ethiopian highlands: assessing the dietary and spatial overlap between mountain nyala and cattle. Afr J Ecol 52:343–351. https://doi.org/10.1111/aje.12126
Atickem A, Loe LE, Langangen Ø, Rueness EK, Bekele A, Stenseth NC (2011) Estimating population size and habitat suitability for mountain nyala in areas with different protection status. Anim Conserv 14:409–418. https://doi.org/10.1111/j.1469-1795.2011.00437.x
Atickem A, Loe LE, Stenseth NC (2014) Individual heterogeneity in use of human shields by mountain nyala. Ethology 7:715–725. https://doi.org/10.1111/eth.12242
Berger J (2007) Fear, human shields and the redistribution of prey and predators in protected areas. Biol Lett 3:620–623. https://doi.org/10.1098/rsbl.2007.0415
Bingham BB, Noon BR (1997) Mitigation of habitat “take”: application to habitat conservation planning. Conserv Biol 11:127–139. https://doi.org/10.1046/j.1523-1739.1997.95331.x
Brown CL, Hardy AR, Barber JR, Fristrup KM, Crooks KR, Angeloni LM (2012) The effect of human activities and their associated noise on ungulate behavior. PLoS ONE 7:e40505. https://doi.org/10.1371/journal.pone.0040505
Butchart SHM, Walpole M, Collen B, van Strien A, Scharlemann JPW et al (2010) Global biodiversity: indicators of recent declines. Science 328(5982):1164–1168. https://doi.org/10.1126/science.1187512
Cattarino L, McAlpine CA, Rhodes JR (2016) Spatial scale and movement behaviour traits control the impacts of habitat fragmentation on individual fitness. J Anim Ecol 85:168–177. https://doi.org/10.1111/1365-2656.12427
Chape S, Harrison J, Spalding M, Lysenko I (2005) Measuring the extent and effectiveness of protected areas as an indicator for meeting global biodiversity targets. Phil Trans R Soc Lond B Biol Sci 360:443–455. https://doi.org/10.1098/rstb.2004.1592
Craigie ID, Baillie JEM, Balmford A, Carbone C, Collen B, Green RE, Huttone JM (2010) Large mammal population declines in Africa’s protected areas. Biol Conserv 143:2221–2228. https://doi.org/10.1016/j.biocon.2010.06.007
Dean AM, Smith GM (2003) An evaluation of per-parcel land cover mapping using maximum likelihood class probabilities. Int J Remote Sens 24:2905–2920. https://doi.org/10.1080/01431160210155910
Deribe E, Bekele A, Balakrishnan M (2008) Population status and diurnal activity patterns of the common warthog (Phacochoerus africanus) in the Bale Mountains National Park, Ethiopia. Int J Ecol Environ Sci 34:91–97
ERDAS (1994) ERDAS Field Guide. Version 8.1. Atlanta, Georgia
Forester JD, Im HK, Rathouz PJ (2009) Accounting for animal movement in estimation of resource selection functions: sampling and data analysis. Ecology 90:3554–3565. https://doi.org/10.1890/08-0874.1
Gaston KJ, Jackson SF, Cantú-salazar L, Cruz-piñón G (2008) The ecological performance of protected areas. Annu Rev Ecol Evol Syst 39:93–113. https://doi.org/10.1146/annurev.ecolsys.39.110707.173529
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1235. https://doi.org/10.1126/science.aar7121
Godvik IMR, Loe LE, Vik JO, Veiberg V, Langvatn R, Mysterud A (2009) Temporal scales, trade- offs, and functional responses in red deer habitat selection. Ecology 90:699–710. https://doi.org/10.1890/08-0576.1
Guisan A, Zimmermann NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135:147–186. https://doi.org/10.1016/S0304-3800(00)00354-9
Harless ML, Walde AD, Delaney DK, Pater LL, Hayes WK (2009) Home range, spatial overlap, and burrow use of the desert tortoise in the West Mojave Desert. Copeia 2:378–389. https://doi.org/10.1643/CE-07-226
Hillman J (1986) Conservation in Bale Mountains National Park, Ethiopia. Oryx 20(2):89–94. https://doi.org/10.1017/S0030605300026314
Jarvis A, Reuter HI, Nelson A, Guevara E (2008) Hole-filled SRTM for the globe Version 4, available from the CGIAR-CSI SRTM 90m database. http://srtm.csi.cgiar.org. Accessed 12 Jan 2021
Johnson DS, Thomas DL, ver Hoef JM, Christ A, (2008) A general framework for the analysis of animal resource selection from telemetry data. Biometrics 64:968–976. https://doi.org/10.1111/j.1541-0420.2007.00943.x
Kie JG (2013) A rule-based ad hoc method for selecting a bandwidth in kernel home-range analyses. Anim Biotelem 1:13. https://doi.org/10.1186/2050-3385-1-13
Knüsel MA, Lee DE, König B, Bond ML (2019) Correlates of home range sizes of giraffes, Giraffa camelopardalis. Anim Behav 149:143–151. https://doi.org/10.1016/j.anbehav.2019.01.017
Lone K, Mysterud A, Gobakken T, Odden J, Linnell J, Loe LE (2017) Temporal variation in habitat selection breaks the catch-22 of spatially contrasting predation risk from multiple predators. Oikos 126:624–632. https://doi.org/10.1111/oik.03486
Malcolm J, Evangelista P (2004) The range and status of the mountain nyala. Technical Report to the Ethiopian Wildlife Conservation Department, Oromia Rural Land and Natural Resource Department
Mysterud A, Pérez-Barbería-Francisco FJ, Gordon IJ (2001) The effect of season, sex and feeding style on home range area versus body mass scaling in temperate ruminants. Oecologia 127:30–39. https://doi.org/10.1007/s004420000562
Noonan MJ, Fleming CH, Tucker MA, Kays R, Harrison AL, Crofoot MC, Abrahms B, Alberts SC, Ali AH et al (2020) Effects of body size on estimation of mammalian area requirements. Conserv Biol 34(4):1017–1028. https://doi.org/10.1111/cobi.13495
Ochiai K, Susaki K, Mochizuki T, Okasaka Y, Amada Y (2010) Relationships among habitat quality, home range size, reproductive performance and population density: comparison of three populations of the Japanese serow (Capricornis crispus). Mamm Study 35:265–276. https://doi.org/10.3106/041.035.0406
Ofstad EG, Herfindal I, Solberg EJ, Saether BE (2016) Home ranges, habitat and body mass: simple correlates of home range size in ungulates. Proc R Soc b: Biol Sci 283:20161234. https://doi.org/10.1098/rspb.2016.1234
Ogutu JO, Piepho HP, Said MY, Ojwang GO, Njino LW, Kifugo SC, Wargute PW (2016) Extreme wildlife declines and concurrent increase in livestock numbers in Kenya: what are the causes? PLoS ONE 11:e0163249. https://doi.org/10.1371/journal.pone.0163249
R Development Core Team (2011) A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rechetelo J, Grice A, Reside AE, Hardesty BD, Moloney J (2016) Movement patterns, home range size and habitat selection of an endangered resource tracking species, the black-throated finch (Poephila cincta cincta). PLoS ONE 11:e0167254. https://doi.org/10.1371/journal.pone.0167254
Rhodes JR, Mcalpine CA, Lunney D, Possingham HP (2005) A spatially explicit habitat selection model incorporating home range behavior. Ecology 86:1199–1205. https://doi.org/10.1890/04-0912
Scholte P (2011) Towards understanding large mammal population declines in Africa’s protected areas: a west-central African perspective. Trop Conserv Sci 1:1–11. https://doi.org/10.1177/194008291100400102
Seaman DE, Millspaugh JJ, Kernohan BJ, Brundige GC, Raedeke KJ, Gitzen RA (1999) Effects of sample size on kernel home range estimates. J Wildl Manag 63:739–747. https://doi.org/10.2307/3802664
Signer J, Balkenhol N (2015) Reproducible home ranges (rhr): a new, user-friendly R package for analyses of wildlife telemetry data. Wildl Soc Bull 39(2):358–363. https://doi.org/10.1002/wsb.539
Sillero-Zubiri C (2013) Tragelaphus buxtoni mountain nyala (Gedemsa). In: Kingdon J, Hoffman M (eds) Mammals of Africa. Vol. VI pigs, hippopotamuses, chevrotain, giraffes, deer and bovids, vol 6. Bloomsbury, London, pp 159–162
Stephens PA, dSa CA, Sillero-Zubiri C, Leader-Williams N (2001) Impact of livestock and settlement on the large mammalian wildlife of the Bale Mountains National Park southern Ethiopia. Biol Conserv 100:307–322. https://doi.org/10.1016/S0006-3207(01)00035-0
Tucker MA, Böhning-Gaese K, Fagan WF, Fryxell JM, Van Moorter B, Alberts SC, Ali AH, Allen AM et al (2018) Moving in the Anthropocene: global reductions in terrestrial mammalian movements. Science 359:466. https://doi.org/10.1126/science.aam9712
Tucker MA, Ord TJ, Rogers TL (2014) Evolutionary predictors of mammalian home range size: body mass, diet and the environment. Global Ecol Biogeogr 23:1105–1114. https://doi.org/10.1111/geb.12194
van Beest FM, Mysterud A, Loe LE, Milner JM (2010) Forage quantity, quality and depletion as scale-dependent mechanisms driving habitat selection of a large browsing herbivore. J Anim Ecol 79:910–922. https://doi.org/10.1111/j.1365-2656.2010.01701.x
van Beest FM, Rivrud IM, Loe LE, Milner JM, Mysterud A (2011) What determines variation in home range size across spatiotemporal scales in a large browsing herbivore? J Anim Ecol 80:771–785. https://doi.org/10.1111/j.1365-2656.2011.01829.x
Waltermire R (1975) A national park in the Bale Mountains. Walia 6:20–23
Willems EP, Hill RA (2009) Predator-specific landscapes of fear and resource distribution: effects on spatial range use. Ecology 90:546–555. https://doi.org/10.1890/08-0765.1
Acknowledgements
We thank Saint Louis Zoo for financially supporting this project. We are very grateful to Dr. David V. Cooper for his help in immobilizing mountain nyalas and fixing GPS collars. We thank Ethiopian Wildlife Conservation Authority for giving the permit to carry out the project.
Funding
This work was supported by Saint Louis Zoo WildCare Institute provided to Dr. Anagaw Atickem.
Author information
Authors and Affiliations
Contributions
AA and LEL conceived the study; AAU and AA did the fieldwork; LEL and MK did the data analysis; AA, MF, and DZ wrote the manuscript. All authors contributed to improving the draft manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
This study has received ethical approval and full support from the Ethiopian Wildlife Conservation Authority.
Research involving human and animal participants
Additional declarations for articles in life science journals report the results of studies involving humans and/or animals. We did collaring of mountain nyala following all the ethical procedures requested by the Ethiopian Wildlife Conservation Authority.
Additional information
Handling editor: Vera Rduch.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Atickem, A., Klapproth, M., Fischer, M. et al. Home range and habitat selection of female mountain nyalas (Tragelaphus buxtoni) in the human-dominated landscape of the Ethiopian Highlands. Mamm Biol 102, 155–162 (2022). https://doi.org/10.1007/s42991-021-00216-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-021-00216-0