
Abstract
Photoperiodism has been shown to be an important synchronizer of seasonal reproduction in many rodent species in the wild; it is a reliable cue as in the southern hemisphere it coincides with the onset of rainfall and hence the availability of food resources for maximal reproductive success. The photoperiodic effect on the reproductive status of the Namibian gerbil, Gerbilliscus cf. leucogaster from central Namibia was investigated. Twenty adult males were exposed to a long-day length (16L:8D), while further 20 adult males were subjected to a short-day length (SD:8L:16D); all for a period of 3 months. Testicular mass per gram body mass, testicular volume and seminiferous tubule diameters were used to assess the effect of photoperiod on gonadal development. Body mass did not significantly differ between the two photoperiodic regimes. The testicular mass per gram of body mass was significantly heavier for the males maintained on a long photoperiod compared to those on a short photoperiod. Similarly, testicular volume and seminiferous tubule diameter were greater in males maintained on a long-day cycle compared to those on the short-day cycle. These findings suggest that G. cf. leucogaster is photoresponsive to day length changes. Photoperiodic changes in the semi-arid habitats can be used to herald the onset of reproduction as it often acts in concert with other proximate cues in desert rodents, but is a constant environmental cue that does not change from year to year, unlike rainfall patterns.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Photoperiod, rainfall and the sudden flush of green vegetation are important factors for the onset of reproduction in rodents inhabiting arid and semi-arid habitats. These cues do not act separately, but in concert with one another for the onset and maintenance of reproduction and maximizing reproductive success (Ben-Zaken et al. 2013). Photoperiod plays a very important role as a proximate factor, triggering reproductive events in terrestrial organisms and in particular those occurring at higher latitudes (Nelson et al. 1992; Rani and Kumar 2014). Rainfall, although sporadic in the semi-arid region of central Namibia, occurs during the spring and summer months when day length is longer. Gerbilliscus cf. leucogaster, is studied for the first time to determine the effects that photoperiod has on the reproductive system in relation to the other well-documented species in the region.
Changes in day length are perceived by the brain through changes in the melatonin signal that is secreted by the pineal gland (Prendergast et al. 2009; Rani and Kumar 2014). The effect of photoperiod in adults is manifested by the recrudescence and subsequent maintenance of reproductive function on long photoperiods and regression of reproductive structures and cessation of reproduction on short photoperiods in long-day animals and the opposite effects in short-day organisms (Yu et al. 1993). The pineal gland and its hormone melatonin have been implicated in the photoperiodic regulation of reproduction in all mammals studied to date (Yu et al. 1993; Rani and Kumar 2014).
Mammals that are responsive to long-day length cease to breed as the day length becomes shorter towards the end of summer and with the onset of autumn (Muteka et al. 2006; Prendergast et al. 2001). Effects of photoperiod on reproduction have been investigated in several mammalian species such as the golden spiny mouse, Acomys russatus (Ben-Zaken et al. 2013), the California vole, Microtus californicus (Nelson et al. 1983), the vole, Microtus agrestis (Spears and Clarke 1986), black rat, Rattus norvegicus (Heideman and Sylvester 1997; Heidemann et al. 1998), the California vole, the four-striped field mouse, Rhabdomys pumilio (Jackson and Bernard 1999), and the white-footed mouse, Peromyscus leucopus (Young et al. 2000). Increasing day length coincides with the beginning of spring and reproductive recrudescence in seasonally breeding animals begins during this period to maximize the survival of the offspring (Jameson 1988; Flowerdew 1987). Spring and autumn are crucial to the survival of many animals in the natural environment as these periods are characterized by the onset of food resources that result from rainfall (Flowerdew 1987; Meheretu et al. 2015). Thus, these periods maximize survival and growth rates of juveniles with the available nutritional supplies (Meheretu et al. 2015; Mutze 2007; Nilsson 2001; Tinney et al. 2001), and favourable ambient temperatures (Benson and Morris 1971; Bronson and Prayor 1983; Bronson 1989), conditions which are associated with spring and summer months of the year.
In natural ecosystems, populations of wild rodents and some domesticated animals rely on photoperiod as a major proximate cue to control reproduction (Muteka et al. 2006; Bronson 2009). However, some studies have shown that photoresponsive animals do not rely on photoperiod alone, but also on other changes in environmental conditions (Nelson et al. 1983; Sicard et al. 1993; Prendergast et al. 2001; Anand et al. 2002; Mutze 2007; Meheretu et al. 2015; Fabio-Braga and Klein 2018). These may include ultimate factors such as food availability and social factors (Anand et al. 2002; Meheretu et al. 2015). Some rodents such as the California vole, may opportunistically stimulate gonadal development or prevent gonadal regression on short days when food is adequate (Nelson et al. 1983; Hamid et al. 2012), in which case, the nutritional effects override the photoperiodic impacts on reproduction. Other environmental variables such as low humidity have also been found to stimulate gonadal development in Kusu rats, Arvicanthis niloticus, but have minimal or no effects in other rodent species (Nelson et al. 1983; Sicard et al. 1993). Thus, photoperiod is a major controlling factor in many terrestrial mammals, but may act in conjunction with other environmental factors (Muteka et al. 2006).
Some rodent species such as the pouched mouse, Saccostomus campestris, mainly breed during summer when reproduction is controlled by photoperiod (Bernard and Hall 1995). Reproduction during winter, however, appears to be inhibited by reduced food availability (Tinney et al. 2001). Similar observations have also been reported in the four-striped mouse, where the inhibition of reproduction during winter is prevented by factors other than photoperiod (Jackson and Bernard 1999). These studies confirm that although photoperiod is a major cue controlling reproduction, additional factors may be simultaneously influential, together with day length (Bronson 1989, 2009). Given that essential changes in environmental conditions vary widely, it is however, likely that their predictability may be less reliable than photoperiod (Muteka et al. 2006).
In arid and semi-arid regions, there is an urgent need to investigate the reproductive aspects of small mammal species, since the parameters controlling reproductive onset are not well-documented as is the case in temperate regions (Bronson 2009). Consequently, this study is the first aimed to determine the effect of photoperiod on male gonads of the Namibian gerbil, G. cf. leucogaster, from the Otjozondjupa and Khomas regions of central Namibia, under controlled laboratory conditions. This study has been undertaken to discern if photoperiod plays a role in the onset of reproductive activation in the male of this gerbil. Morphological and histological methods were used to investigate the reproductive changes in males G. cf. leucogaster. We, therefore, hypothesised that the Namibian gerbil, G. cf. leucogaster is a seasonal breeder that is responsive today length; or that the reproduction in the Namibian gerbils, is not confined to a specific season and is non responsive to changes in day length.
Materials and methods
Study areas
A total of 40 adult male specimens of G.cf. leucogaster were sampled from Otjinakwi Farm, in the Otjozondjupa region (− 20° 45′ 3.81″ + 17°, 1′ 21.80″) at 1440.24 m above sea level (a.s.l.) and Neudam Farm, in the Khomas Region (− 22° 30′ 16″ + 17° 22′ 9″) at 1656 m a.s.l. Both regions are situated in the central part of Namibia and are characterized by a semi-arid climate with an average annual temperature of 19.47 °C (Goddard Institute 1957–1987; Namibia Meteorological Service 2014).
The winter months (June–August), usually experience little or no rain and minimum temperatures range between − 5 and 18 °C. Nights are usually cool and very cold before dawn (Goddard Institute 1957–1987). Days are usually warm to hot, ranging from a maximum of 20 °C in July to 31 °C in January with mean annual rainfall around 360 mm (Goddard Institute 1957–1987). The natural vegetation of the area is scrub and steppe (savannah woodland) (Goddard Institute 1957–1987; Namibia Meteorological Service 2014).
Trapping and handling of animals
A mixture of peanut butter, syrup, oat meal, and fish oil was used as bait. The gerbils were trapped using Sherman live traps that were set prior to twilight and subsequently re-checked at dawn. Live animals were sampled and processed as approved by the Animal Ethics (Ethics clearance number EC066-17) Committee of the University of Pretoria, Pretoria, South Africa, with the necessary field collection permit issued by the Ministry of Environment and Tourism, Windhoek, Namibia. The animals were trapped during mid-January and were subjected to experimentation within 24 h after field capture to avoid acclimatization. The animals were kept in polyurethane cages with wood shavings provided as bedding. Forty male G. cf. leucogaster were selected based on body mass (grams) to discern the adults from the juveniles; of which 20 were subjected to a long-day (16L:8D), and twenty to a short-day (8L:16D) lighting scheme, all for a period of 3 months. Mice pellets (EPOL, Westville and Durban, South Africa) with a balanced carbohydrate, protein and fat content were given and water was provided ad libitum for the duration of the photoperiodic treatments.
The animals were exposed to the two lighting regimes, under constant environment conditions in a laboratory. The long-day length (LD) was set at 16 h light and 8 h darkness (16L:8D), with lights on at 06h00 and off at 22h00. Short-day length (SD) was set at 8 h of light and 16 h darkness (8L:16D), with lights on at 07h00 and off at 15h00. The photoperiodic lengths were selected to simulate the day lengths experienced during natural summer and winter periods in the Khomas and the Otjozondjupa regions of central Namibia. Temperature was controlled at 26 ± 3 °C (n = 980) with relative humidity levels being maintained at 42 ± 5%. The light intensities in the rooms were 552 ± 37 lm/m2 (Canon light meter).
Processing of samples
After 90 days, the animals were sacrificed using an overdose of halothane and body mass was obtained using a Mettler digital balance (Ohaus Corp. Pine Brook, NY, USA). The testes were removed and weighed, the length and width of each testes was measured using a pair of digital callipers (Sylvac Opto RS 232, Ultra Praezision Messzeuge GmbH, Germany). Testicular volume was calculated using the formula for the volume of an ellipsoid described by Woodall and Skinner (1989) as follows:
where a = 1/2 maximum length and, b = 1/2 maximum breadth.
Gonads were then placed into Bouin’s fluid for a minimum of 24 h for fixation before being rinsed and stored in 70% alcohol. Voucher specimens were deposited in the Natural History Museum, Windhoek, Namibia.
Mitochondrial typing
DNA was extracted from a subset of tissue samples using the Roche High Pure Template Preparation kit (Roche). The cytochrome b gene region was amplified and sequenced as described by Bastos et al. (2011). The resulting sequences (deposited under accession numbers KT029850–KT029851) were used in nucleotide Blast searches against the Genbank database (www.ncbi.nlm.nih.gov/blast) to identify reference sequences with the highest levels of sequence identity to the gerbils sampled from central Namibia. The mitochondrial typing was done to establish which species of gerbil was being studied as there are several species that are morphologically indistinguishable.
Histology
All the testes were sectioned, mounted, and stained following the guidelines of Ross et al. (1995) and Leeson et al. (1985). Seminiferous tubule diameters were determined by selecting 100 testicular sections with circular tubules and photographs taken at 40 × magnification using a DMX 1200 Nikon digital camera and Image Tools software version 3.0 (Melville, New York, USA). Following Ross et al. (1995), all testicular sections were also examined for signs of spermatogenesis, spermatozoa and stages of testicular development.
Data analysis
Analysis of variance (ANOVA; McCullagh and Nelda 1989) was used to test for any body mass differences between males on long and short photoperiods as well as to test for differences in seminiferous tubule diameter, testicular mass and testicular volume. The data were analysed using algorithms in Microsoft Excel version 14 (McCullagh and Nelda 1989) and Statistical Application Software (SAS) version 9.1 (McDonald 2014a, b).
Results
Molecular typing
Nucleotide searches against the Genbank database, using full length cytochrome b gene sequences generated for specimens from both sampling sites in central Namibia confirmed that the two closest matches in this public database (AM40989-90) corresponded to Gerbilliscus leucogaster sampled from South Africa (92.6% sequence identity). In light of relatively high level of sequence divergence (7.4%), G. cf. leucogaster is used to denote the species evaluated in this study.
Body mass
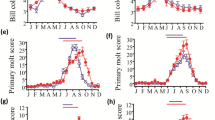
There was no statistically significant difference between the initial body mass of the G. cf. leucogaster males placed on a long-day length [ANOVA: body mass (LD) = 70.0 ± 2.1 g; n =20] and the initial body mass of the animals on a short-day length [ANOVA: body mass (SD) = 69.1 ± 2.6 g; n =20] (t test: n = 20; P = 0.43) at the start of the experiment (Fig. 1a).

Mean ± 1 standard error (SE) of male Namibia gerbils, Gerbilliscus cf. leucogaster from the Otjozondjupa and Khomas Regions of central Namibia showing: a initial and final body mass; b specific testicular mass (mg/g); c testicular volume (mm3); and d seminiferous tubule diameter (µm) after being subjected to long (LD) and short (SD) photoperiodic conditions
At the end of the experiment, the animals that were subjected to the long photoperiod (LD: 72.9 ± 1.01 g; F(2,20)=0.42; n = 20; P = 0.02) significantly gained 2.9 g of body mass, whereas those maintained on a short-day length (SD: 65.2 ± 1.5 g; F(2,20) = 1.16; n = 20; P = 0.001 showed a significant drop of 3.9 g in body mass (SD: (Fig. 1a).
Testicular mass and volume, and seminiferous tubule diameter
Testicular mass expressed as a function of a gram of body mass in G. cf. leucogaster was significantly higher for males maintained on LD (19.0 ± 2.0 mg/g; n = 2 0) than those males on SD (SD: 12.0 ± 3.00 mg/g; n =20) (two-way ANOVA: F(2,20) = 6.98; n = 20; P =0.01) (Fig. 1b). Similarly, testicular volume was significantly greater for animals on LD (LD = 635 ± 83.36 mm3; n =20) than those on SD (356.0 ± 51.12 mm3; n = 20) (two-way ANOVA: F(2,20) = 2.68; n = 20; P =0.01) (Fig. 1c). Mean seminiferous tubule diameter was significantly larger in males on LD (105.6 ± 4.6 μm; n =20) than on SD (81.24 ± 3.4 μm; n =20) (two-way ANOVA: F(2,20) = 5.93; n = 20; P < 0.05) (Fig. 1d).
Discussion
A number of studies have shown that photoperiod is a major environmental cue for controlling reproduction in the wild (Ims 1990; Sangeeta and Vinod 2014). The findings in the present study demonstrate that individual male G. cf. leucogaster respond to a long-day photoperiod with variable reproductive responses. From a physiological perspective, the obtained data indicate that long days serve as an environmental signal stimulating reproduction in G. cf. leucogaster from central Namibia. Research has shown that as a result of unpredictable climatic conditions in deserts, reproduction in desert species may not be successful in any given year, such that desert rodents breed opportunistically depending on relatively short-term climatic and nutritional conditions relying more on a nutritional resource base other than photoperiod (Kenagy and Bartholomew 1985; Rani and Kumar 2014).
Seasonal changes in the environment can affect a wide range of physiological and behavioural processes in most tropical and sub-tropical mammalian species. Food availability, rainfall, temperature, and day length (i.e., photoperiod) are among the environmental factors that affect the time of onset and the duration of the reproductive period in mammals (Nilsson 2001). A variety of other seasonal physiological responses often accompany the photoperiodically induced alterations in reproductive status and may include changes in pelage colouration or thickness (Duncan and Goldman 1984; Tavolaro et al. 2015), body fat deposition or utilization (Bartness and Wade 1985), and thermogenesis (Heldmaier et al. 1981). These seasonal responses also are typically regulated by changes in photoperiod (Bartness and Goldman 1989). The cessation of all breeding activity for several months during autumn and winter, therefore, represents a significant period in the life of an individual small mammal during which all opportunity for reproduction ceases (Muteka et al. 2006; Sangeeta and Vinod 2014).
Bronson (1985) and Forger and Zucker (1985) and Tavolaro et al. (2015) suggested that long photoperiod may promote somatic growth. The present study found animals on long-day length to have gained body mass, whereas those on short-day length displayed a significant dropped in body mass. This observation confirms that photoperiod does have effects on somatic growth as has been reported in Rattus norvegicus retrospectively by Bronson (1985), Forger and Zucker (1985), Heidemann et al. (1998). On the other hand, somatic effects may be more apparent in growing juveniles than that in adults, as all the animals used in the present study were adults.
Male G. cf. leucogaster showed a significant gonadal and somatic regression when subjected to short photoperiods in this study. Similar findings were obtained in adult Siberian hamsters, which exhibited gonadal regression on short-day lengths in both sexes (Schlatt et al. 1993; Wade and Bartness 1984; Alibhai 1986). The increase in the values of testicular mass, volume and seminiferous tubule diameters between males maintained under long photoperiod and those on short photoperiods suggests a positive response to the effects of long photoperiod and gonadal regression on short photoperiods. Comparing males of similar age and gonadal sizes in the natural population, males did not exhibit gonadal regression when exposed to a short photoperiod as observed in some mammals such as Syrian hamsters in which individuals subjected to short photoperiod had regressed testes relative to animals exposed to long photoperiods (Bartness and Wade 1985; Tavolaro et al. 2015).
A number of rodents have been reported to exhibit testicular regression during a short photoperiod to avoid adverse environmental effects on their offspring (Schlatt et al. 1993; Tavolaro et al. 2015). In the grasshopper mouse, Onychmys leucogaster, gonadal recrudescence is inhibited on a photoperiod of 10L:14D, but stimulated on a photoperiod of 14L:10D (Frost and Zucker 1983). A similar study on the desert pocket mouse, Perognathus formosus, showed that testicular development and recrudescence is stimulated when subjected to a 16L:8D photoperiod, but inhibited when subjected to 8L:16D day length (Kenagy and Bartholomew 1981). Muteka et al. (2006) showed that the Tete veld rat, Aethomys ineptus and the Namaqua rock mouse, Micaelamys namaquensis both utilize photoperiod to time the appropriate reproductive period in the wild.
In the Tete veld rat, the failure of male animals to undergo gonadal regression on a short-day length has been proposed to be adaptive as this enables reproductively mature males the potential to continue their reproductive efforts during winter (Kriegsfeld et al. 1999). This observation is further supported by the presence of spermatozoa in the epididymis and minimal spermatogenic activity in the seminiferous tubules of the males of G. cf. leucogaster during winter months. Although the offspring live under harsh conditions during winter, the possibility exists that a litter may be successfully reared if conditions for survival, such as adequate food resources or favourable microclimatic conditions such as temperature are suitable (Bukreeva and Lidzhi-Garyaeva 2018). The possibility of winter breeding in the Namibian gerbil could be due to one of several reasons: First, gerbils may have a photoperiodic system much like that of typical LD-breeding rodents, but the potential inhibitory effect of SDs is overridden by other environmental factors such as ambient temperature, rainfall, humidity, or food availability as observed in Microtus socialis (Bukreeva and Lidzhi-Garyaeva 2018); second, the gerbils may be reproductively nonresponsive to the photoperiod and instead use other predictors to time their reproduction; Thirdly, seasonality in these animals may be a consequence of an opportunistic strategy in which reproduction is restricted to the times when climatic and nutritional conditions are optimal without the use of any environmental predictors. This is even more so in winter breeding rodents such as M. socialis that breeds during the winter in the northern hemisphere (Bukreeva and Lidzhi-Garyaeva 2018). Thus, other cues may be more accurate than photoperiod that these may override the photoperiodic effects when available in abundance.
The significantly higher testicular mass, volume, and seminiferous tubule diameter in animals maintained on a long day compared to those on a short day, implies an important role for photoperiodism in the recrudescence and regression of reproductive activity in G. cf. leucogaster in central Namibia. It is reported that in some animals exposed to a short-day length their gonads do not completely regress. Rather there is a cessation of spermatogenic activity on a short-day length, as has been reported in the Egyptian spiny mouse, Acomys cahirinus, Anderson’s gerbil, Gerbillus andersoni (El-Bakry et al. 1998; Kerbeshian et al. 1994; Kerbeshian and Bronson 1996), the Deer mouse, Peromyscus maniculatus (Scheffer 1924; Whitestt and Miller 1982); the pouched mouse, Saccostomus campestris (Bernard and Hall 1995), and the Siberian hamster, Phodopus sungorus (Hoffmann 1973). In young male F344 rats, Heidemann et al. (1998) reported a inhibition of testicular regression on day length of 16L:8D, partial regression on 14L:10D, and a total regression on a photoperiod of less or equal to 10 h light/day In G. cf. leucogaster, it is possible that there is critical photoperiodic threshold required for complete testicular regression other than the two photoperiodic regimes to which the animals were exposed.
The present study demonstrates that G. cf. leucogaster is responsive to changes in photoperiod. It should however, be mentioned that other environmental variables may act synergistically to promote breeding at a particular season. Although it was demonstrated that reproductive activity in male G. cf. leucogaster is heavily influenced by exposure to different photoperiods, other environmental factors such as rainfall, food availability, and ambient temperature may also play a subtle role in the timing of reproduction in these species within their environments. However, photoperiod is clearly an important proximate cue for controlling the onset of reproductive parameters in G. cf. leucogaster from central Namibia.
References
Alibhai SK (1986) Reproductive response of Gerbillus harwoodii to 6-MBOA in the Kora National Reserve, Kenya. J Trop Pediatr 2:377–379
Anand S, Losee-Olson S, Turek FW, Horton TH (2002) Differential regulation of luteinizing hormone and follicle-stimulating hormone in male Siberian hamster by exposure to females and photoperiod. Endocrinology 143:2178–2188
Bartness TJ, Goldman BD (1989) Mammalian pineal melatonin: a clock for all seasons. Experientia 45:939–945
Bartness TJ, Wade GN (1985) Seasonal body weight cycles in hamsters. Neurosci Biobehav Rev 9:599–612
Bastos ADS, Nair D, Taylor PJ, Brettschneider H, Kirsten F, Mostert ME, von Maltitz E, Lamb JM, van Hooft P, Belmain SR, Contrafatto G, Downs S, Chimimba CT (2011) Genetic monitoring detects an overlooked cryptic species and reveals the diversity and distribution of three invasive Rattus congeners in South Africa. BMC Genet 12:26
Benson GK, Morris LR (1971) Foetal growth and lactation in rats exposed to high temperatures during pregnancy. J Reprod Fertil 27:369–384
Ben-Zaken I, Haim A, Zubidat AE (2013) Long-day photoperiod interacts with vasopressin and food restriction to modulate reproductive status and vasopressin receptor expression of male golden spiny mice. J Exp Biol 216:3495–3503. https://doi.org/10.1242/jeb.089607
Bernard RTF, Hall J (1995) Failure of the oestrous cycle and spermatogenesis to respond to day length in subtropical African rodent, the pouched mouse (Saccostomus campestris). Biol Reprod 52:1291–1296
Bronson FH (1985) Mammalian reproduction: an ecological perspective. Biol Reprod 32:1–26
Bronson FH (1989) Regulation by social cues. In: Bronson FH (ed) Mammalian reproductive biology. University of Chicago Press, Chicago
Bronson FH (2009) Climate change and seasonal reproduction in mammals. Philos Trans R Soc Lond Biol 364:3331–3340
Bronson FH, Prayor S (1983) Ambient temperature and reproductive success in rodent living at different latitudes. Biol Reprod 29:72–80
Bukreeva OM, Lidzhi-Garyaeva GV (2018) Mass migration of social voles (Microtus socialis Pallas, 1773) in the Northwestern Caspian Region. Arid Ecosyst 8:147–151. https://doi.org/10.1134/S2079096118020038
Duncan MJ, Goldman BD (1984) Hormonal regulation of the annual pelage color cycle in the Djungarian hamster, Phodopus sungorus. II. Role of prolactin. J Exp Zool 230:97–103
El-Bakry HA, Zaharan WM, Bartness TJ (1998) Photoperiod responses of four wild trapped desert rodent species. Am J Physiol 275:2012–2022
Fabio-Braga AP, Klein W (2018) Temperature and circadian effects on metabolic rate of South American echimyid rodents, Trinomys setosus and Clyomys bishopi (Rodentia: Echimyidae). Zoologia 35:1–6
Flowerdew JR (1987) Effects of the environment on reproduction. In: Flowerdew JR (ed) Mammals, their reproductive biology and population ecology. Edward Arnold Publishers Ltd, London
Forger NG, Zucker I (1985) Photoperiodic regulation of reproductive development in male white-footed mice (Peromyscus leucopus) born at different phases of breeding season. J Reprod Fertil 73:271–278
Frost D, Zucker I (1983) Photoperiodic and melatonin influences seasonal gonadal cycles in the grasshopper mouse (Onychomys leucogaster). J Reprod Fertil 69:237–244
The Goddard Institute for Space Studies (1957–1988) Columbia University, New York City, USA
Hamid HY, Zakaria MZAB, Meng GY Haron, Abd W, Mustapha NM (2012) Effects of elevated ambient temperature on reproductive outcomes and offspring growth depend on exposure time. Sci World J 2012:1–6. https://doi.org/10.1100/2012/359134
Heideman PD, Sylvester CJ (1997) Reproductive photo responsiveness in unmanipulated male Fischer 344 laboratory rats. J Reprod 57(1):134–138
Heidemann PD, Deiler RW, York M (1998) Food and neonatal androgen interact with photoperiod to inhibit reproductive maturation in Fischer 344 rats. Biol Reprod 59:358–363
Heldmaier G, Steinlechner S, Rafael J, Vsiansky P (1981) Photoperiodic control and effects of melatonin on non-shivering thermogenesis and brown adipose tissue. Science 212:917–919
Hoffmann K (1973) The infl uence of photoperiod and melatonin on testis size, body weight, and pelage colour in the Djungarian hamster (Phodopus sungorus). J Comput Physiol A 85:267–282
Ims RA (1990) The ecology and evolution of reproductive synchrony. Trends Ecol Evol 5:135–140
Jackson C, Bernard RTF (1999) Short day length alone does not inhibit spermatogenesis in the seasonally breeding four-stripped field mouse (Rhabdomys pumilio). Biol Reprod 60:1320–1323
Jameson EW (1988) The patterns of seasonality, Ch. 13. In: Jameson EW (ed) Vertebrate reproduction. A Wiley Interscience publication. Wiley, New York
Kenagy J, Bartholomew A (1981) Effects of day length, temperature and green food on testicular development in a dessert mouse, Perognathus formosus. Physiol Zool 54:62–73
Kenagy GJ, Bartholomew GA (1985) Seasonal reproductive patterns in five coexisting California desert rodent species. Ecol Monogr 55:371–397
Kerbeshian MC, Bronson FH (1996) Running-induced testicular recrudescence in the meadow vole: role of the circadian system. Physiol Behav 60:165–170
Kerbeshian MC, Bronson FH, Bellis ED (1994) Variation in reproductive photo responsiveness in a wild population of meadow voles. Biol Reprod 50:7445–7450
Kriegsfeld LJ, Drazen DL, Nelson RJ (1999) Effects of photoperiod and reproductive responsiveness on pituitary sensitivity to GnRH in male prairie voles (Microtus ochrogaster). Gen Comp Endocrinol 116(2):221–228
Leeson CR, Leeson TS, Paparo AA (1985) Textbook of histology. WB Saunders Company, Philadelphia
McCullagh P, Nelda J (1989) General linear models, 2nd edn. Chapman and Hall, Boca Raton
McDonald JH (2014a) Using spreadsheets for statistics. In: McDonald JH (ed) Handbook of biological statistics. Sparky House Publishing, Baltimore, pp 266–273
McDonald JH (2014b) Getting started with SAS. In: McDonald JH (ed) Handbook of biological statistics. Sparky House Publishing, Baltimore, pp 285–292
Meheretu Y, Welegerima K, Sluydts V, Bauer H, Gebrehiwot K, Deckers J, Makundi R, Leirs H (2015) Reproduction and survival of rodents in crop fields: the effects of rainfall, crop stage and stone-bund density. Wildl Res 42(2):158–164. https://doi.org/10.1071/WR14121
Muteka SP, Chimimba CT, Bennett NC (2006) Reproductive photoresponsiveness in Aethomys ineptus and A. namaquensis (Rodentia: Muridae) from southern Africa. J Zool Lond 268:225–231
Mutze G (2007) Does high growth rate of juvenile house mice with prolonged access to ripening grain and water drive population outbreaks? N Z J Zool 4:195–202
Namibia Meteorological Service (2014) 12C Hugel Street, Windhoek, Namibia
Nelson RJ, Dark J, Zucker I (1983) Influence of photoperiod, nutrition and water availability on the reproduction of male California vole (Microtus californicus). J Reprod Fertil 69:473–477
Nelson RJ, Kita M, Blom MC, Rhyme-Grey J (1992) Photoperiod influences the critical caloric intake necessary to maintain reproduction among male deer mice (Peromyscus maniculatus). Biol Reprod 46:226–232
Nilsson P (2001) The biological triangle of life. Interaction between nutrition, reproduction and longevity. Lakartidningen 98:1797–1800
Prendergast BJ, Kriegsfeld LJ, Nelson RJ (2001) Photoperiodic polymorphism in rodents: neuroendocrine mechanisms, costs, and functions. Q Rev Biol 76:293–325
Prendergast BJ, Zucker I, Nelson RJ (2009) Seasonal rhythms of mammalian behavioral neuroendocrinology. In: Pfaff D, Arnold A, Etgen A, Fahrbach S, Moss R, Rubin R (eds) Hormones, brain, and behavior, vol 1, 2nd edn. Academic Press, San Diego
Rani S, Kumar V (2014) Photoperiodic regulation of seasonal reproduction in higher vertebrates. Indian J Exp Biol 52:413–419
Ross MH, Romrell LJ, Kaye GI (1995) Histology: text and atlas. William & Wilkins, Baltimore
Sangeeta R, Vinod K (2014) Photoperiodic regulation of seasonal reproduction in higher vertebrates. Indian J Exp Biol 52(5):413–419
Scheffer TH (1924) Notes on the breeding of Peromyscus. J Mammal 5:258–260
Schlatt S, Niklowitz P, Hoffmann K, Nieschlag E (1993) Influence of short photoperiods on reproductive organs and estrous cycles of normal and pinealectomized female Djungarian hamsters, Phodopus sungorus. Biol Reprod 49:243–250
Sicard B, Fuminier F, Maurel D, Boissin J (1993) Temperature and water conditions mediate the effects of day length on the breeding cycle of a Sahelian rodent, Arvicanthis niloticus. Biol Reprod 49:716–722
Spears N, Clarke JR (1986) Effects of male presence and of photoperiod on the sexual maturation of the field vole (Microtus agrestis). J Reprod Fertil 78:231–238
Tavolaro FM, Thomson LM, Ross AW, Morgan PJ, Helfer G (2015) Photoperiodic effects on seasonal physiology, reproductive status and hypothalamic gene expression in young male F344 rats. J Neuroendocrinol 27:79–87
Tinney GM, Bernard RTF, White RM (2001) Influences of food quality and quantity on the male reproductive organs of a seasonally breeding rodent, the pouched mouse (Saccostomus campestris), from a seasonal but unpredictable environment. Afr Zool 36:23–30
Wade GN, Bartness TJ (1984) Effects of photoperiod and gonadectomy on food intake, body weight, and body composition in Siberian hamsters. Am J Physiol 246:R26–R30
Whitestt JM, Miller LL (1982) Photoperiod and reproduction in female deer mice. Biol Reprod 26:296–304
Woodall PF, Skinner JD (1989) Seasonality of reproduction in male rock shrews, Elephantulus myurus. J Zool Lond 217:203–212
Young KA, Zirkin BR, Nelson RJ (2000) Testicular regression in response to food restriction and short photoperiod in white-footed mice (Peromyscus leucopus) is mediated by Apoptosis. Biol Reprod 62:347–354
Yu H-S, Reiter RJ, Goldman BD, Nelson RJ (1993) Melatonin and seasonality in mammals. In: Yu H-S, Reiter RJ (eds) Melatonin—biosynthesis, physiological effects, and clinical applications. CRC Press, Boca Raton, pp 225–252
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Sabine Begall.
Rights and permissions
About this article

Cite this article
Muteka, S.P., Chimimba, C.T., Bastos, A.D.S. et al. Photoperiodic effects on the male gonads of the Namibian gerbil, Gerbilliscus cf. leucogaster from central Namibia. Mamm Biol 100, 165–171 (2020). https://doi.org/10.1007/s42991-020-00008-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-020-00008-y