
Abstract
Antioxidant complex enzymes have a significant role in cellular homeostasis control in plants, and they inhibit the toxic action of reactive oxygen species when they are in excess. There are many antioxidant enzymes executing this role; among these, superoxide dismutase, catalase, and ascorbate peroxidase are reported as the most studied in this process, as they prevent free radicals from becoming more reactive and toxic to cells. Thus, this research was conducted to evaluate antioxidant enzyme expression in response to hydric stress at the reproductive stage in upland rice genotypes. Three genotypes from the upland rice breeding program on agreement between UFLA, EPAMIG, and EMBRAPA, CMG2093, CMG2172, and BRSMG Relâmpago, were used as controls. Genotypes were grown under field conditions with supplementary irrigation during the whole crop cycle, and hydric stress was induced in the reproductive phase before panicle emission. Seedlings were used in enzyme analyses from the emergence test and IVE on substrate (soil + sand at a 2:1 rate) at 70% and 10% field capacity. Significant differences were observed among genotypes for vigor tests. In biochemical tests, the CMG2093 genotype had lower damage on hydric deficit, with the best performance under hydric restriction conditions, being considered tolerant for this stress type.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rice is a food for more than 50% of the world’s population. According to the FAO (2019), Brazil is a main rice producer in the world, being the only country out of Asia among the ten highest production countries. Rice is also essential for combating hunger and is rich in energy and nutrients.
It is a crop with large potential for Brazil, mainly if analyzed uplands rice, which is little exploited on the country despite the large area under its cultivation. According to CONAB (2022), upland rice represents < 20% of national production. However, there is a need for research that involves savannah-adapted cultivar exploitation, as rice is a crop with high water absorption needs, and regions where upland rice cultivation fields have drought periods, which may result in low productivity in relation to irrigated rice cultivation.
Rice is drought-sensitive during flowering and grain filling periods, in which damage can be irreversible, resulting in large production losses. Seeds are also highly affected by hydric stress, in terms of both formation and initial culture development. Fields with drought incidence produce seeds with lower quality and quantity once water deficit has a direct effect on reserve accumulation and no pollination process (Taiz and Zeiger 2013; You and Chan 2015).
The damage caused in culture is caused by an increase in reactive oxygen species (ROS) in plants, resulting in an imbalance in metabolic activities and oxidative stress (Sies and Cadenas 1985). Research has shown that due to an increase in water stress, plants tend to increase ROS production due to changes in metabolism caused by water restriction (Asada 2006; Jaleel et al. 2009; Czarnocka and Karpiński 2018). However, these substances are natural for any aerobic organism and have many metabolic roles (Basra et al. 2006; Czarnocka and Karpiński 2018). Antioxidant complex enzymes are responsible for maintaining balance and avoiding the oxidative activity of those molecules; in general, those enzymes can transform free radicals into nontoxic products.
The antioxidant complex is composed mainly of the enzymes superoxide dismutase (SOD) and catalase (CAT) and peroxidases such as ascorbate peroxidase (APX). SOD is the first defense against ROS by dismutation of superoxide radicals to hydrogen peroxide, which is a less toxic molecule (Fridavich 1995). Catalase is the main enzyme responsible for neutralizing hydrogen peroxide when it is in excess in cell compartments (Sharma et al. 2012). APX enzymes are more efficient at ROS removal, with high affinity for molecules in many cell compartments (Wang et al. 1999). Many studies have reported a correlation between enzyme activity and hydric stress in rice plants (Mittler et al. 2004; Zandalinas et al. 2018). Thus, this research was carried out to evaluate enzymes from the antioxidant complex in relation to the response to hydric stress during the reproductive phase in upland rice genotypes.
Materials and methods
We used seeds from the upland rice breeding program from the agreement between the Federal University of Lavras, EPAMIG, and EMBRAPA Rice and Bean. Seeds from harvest 2018/19 produced at Porangatu, Goias, Brazil, were used. We used three lineages: BRSMG Relâmpago as the hydric stress tolerant (witness) (De Castro et al. 2014) and lineages CMG2172 and CMG2093. Those seeds were produced under two conditions, the first with irrigation fields and the second on fields with no irrigation, resulting in two samples for each lineage: irrigated (irrigated fields) and nonirrigated (nonirrigated fields).
After seed receipt, we measured the water content according to the method suggested by Brasil (2009). We used duplicates for each genotype, with 25 seeds each, placed in an oven with forced air circulation at 105 °C for 24 h.
For the emergence test, we used four replicates of 25 seeds each on acrylic trays filled with substrate (soil + sand at a 2:1 ratio) moistened under two water restriction conditions: 10 and 70% field capacity. Trays were kept in a plant growing chamber at a constant temperature of 25 °C with daily irrigation aiming to maintain water retention according to tray weight every 24 h. Fourteen days after sowing, we measured seedling emergence (hypocotyl totally emerged from substrate). The results are expressed as percentages. The emergence speed index was measured by daily counting after daily emergence counting, and the index was calculated according to the method proposed by Maguire (1962).
Biochemical analyses were carried out by using dry seeds (before germination tests) and seedlings from emergence tests. The samples were kept in a deep freezer (− 80 °C) until the test procedures were performed according to the following method. We measured SOD (EC 1.15.1.1), CAT (E.C. 1.11.1.6), and POX (EC. 1.11.1.7) through electrophoresis. Enzyme extracts were obtained by homogenizing 100 mg macerated samples on 250 µL buffer (0.2 M Tris HCl pH 8.0; 0.1% beta-mercaptoethanol). The solution was homogenized on a vortex and kept for 12 h in a refrigerator, followed by centrifugation at 14,000 rpm for 30 min at 4 °C.
Electrophoresis was executed in discontinuous acrylamide gels at 7.5% (separator) and 4.5% (concentrator). The system gel/electrode was Tris–glycine pH 8.9. We applied 60 µL of extract to a gel, and electrophoresis was carried out at 120 V for 5 h under refrigeration at 4 °C. After processing, gels were revealed according to Alfenas (2006). For each enzyme, we used three gels, representing biological replicates. Each replicate was used for visual analysis of band intensity using ImageJ® software at mm2 units.
The total protein concentration was determined by using the Coomassie Brilliant Blue link method proposed by Bradford (1976), using bovine serum albumin (BSA) as the standard and a wavelength of 595 mm. Aliquots were added to 1 mL of Bradford reagent, mixed by vortexing, and reacted for 15 min.
Lipidic peroxidation was carried out by malondialdehyde (MDA) produced by an extract reaction with thiobarbituric acid (TBA) according to Dhindsa et al. (1981), with adaptations. A sample was macerated and homogenized with 1.25 mL of trichloroacetic acid (0.1%) and 1% sodium dodecyl sulfate (SDS). The homogenate was centrifuged at 12,000 rpm for 15 min. An aliquot of 300 µL of supernatant was mixed with 1 mL of 20% trichloroacetic acid and transferred to a recipient with 0.5% thiobarbituric acid. The mix was heated at 95 °C for 30 min and placed on fast cooling in an ice bath. Absorbance was measured at 532 nm, and MDA concentration was calculated using an extinction coefficient of 155 mM−1 cm−1 (Buege and Aust 1978).
For antioxidant system enzyme measurement, 200 mg samples were macerated in liquid nitrogen and homogenized in 1.5 mL of extraction buffer [100 mM potassium phosphate, 0.1 mM EDTA, 10 mM ascorbic acid]. Samples were homogenized and centrifuged at 13,000 rpm for 30 min at 4 °C, and the supernatant was collected for enzymatic activity analyses (Biemelt et al. 1998).
SOD was determined by measuring the ability to inhibit the photochemical reduction of nitro blue tetrazolium (NBT), as proposed by Giannopolitis and Ries (Giannopolits and Ries 1977a, b). Samples were added to a mix composed of 50 nM potassium phosphate, 13 mM methionine, 0.1 µM EDTA, 75 µM NBT, and 2 µM riboflavin. Samples and control (mix + water instead of extracts) were kept on 20 W fluorescent light at room temperature for 7 min. The absorbance at 560 nm was measured. Each SOD unit is defined by the enzyme necessary to inhibit 50% of NBT reduction.
CAT activity was determined by the decrease in hydrogen peroxide concentration measured by a 240 nM absorbance decrease. For this, 9 µL of samples (or water instead of those) was applied in previously heated at 30 °C reaction buffer [100 mM potassium phosphate], and 9 µL hydrogen peroxide (12.5 mM) was applied. Absorbance was measured at 240 nm every 15 s for 3 min (Havir and McHale 1987). CAT activity is defined by the amount of enzyme necessary to decompose 1 µmol/min of H2O2.
APX was determined by reduction of ascorbate absorbance at 290 nm every 15 s for 3 min according to Nakano and Asada (1981). For this, 9 µL of extract (or water on witness) was added to 162 µL of reaction buffer [100 mM potassium phosphate pH 7.0 + 0.5 mM ascorbic acid] previously heated at 30 °C. To the mixed sample + reaction buffer, 9 µL hydrogen peroxide (2 mM) was added. The absorbance was measured on ELISA equipment.
For emergence and IVE tests, we used a completely random design on a factorial scheme or 3 (lineages) × 4 (water conditions, i.e., 2 field capacities combined with 2 irrigation schemes). The results were analyzed through ANOVA, and if observed differences among treatments were analyzed with the F test, averages were compared through Tukey’s test at 5% probability. All analyses were executed in R for Windows software (R Core Team 2021). For biochemical tests, we used averages among biological replicates and standard deviations.
Results
Both selected genotypes for molecular analysis were submitted to emergence percentage and speed (ESI) in simulated environments with optimal 70% of field capacity and one with water restriction (10% of field capacity). For the emergence test (Table 1), independent of the imposed conditions, CMG2093 was superior to CMG2172 and was not significantly different from BRMG Relâmpago. Thus, CMG2093, in addition to having more vigorous seeds, was able to develop better hydric deficit conditions.
The ESI results corroborate those from the emergence at 70% soil field retention capacity, which leads to the conclusion that, under ideal conditions, CMG2093 is superior to CMG2172 and is not different when compared to witness in both seed production environments: with and without irrigation. However, under water restriction conditions at 10% field capacity, lineages CMG2093 and CMG2172 were not significantly different independently of the seed production environment, being superior to CMG2172 only for seeds produced on irrigated fields (Table 1).
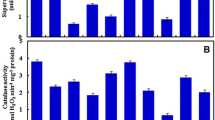
We found higher amounts of protein content in seeds, independent of the cultivar and condition, than in seedlings at both 70% and 10% field capacity. On seeds, independent of the condition imposed, CMG2172 was the cultivar with higher amounts of protein (Fig. 1) compared to the others. As seedlings were observed, CMG2172 had superior values for protein content compared to the others at 70% field capacity. However, at 10% field capacity, CMG2172 and CMG2093 had increases in those values (Fig. 1).

Total protein on rice seeds and seedlings, produced with and without irrigation and seedlings produced from these production systems, at conditions of 70 and 10% field capacity for cultivars CMG2172, BRSMG Relâmpago and CMG2093. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
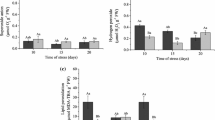
The results for hydrogen peroxide content are shown in Fig. 2. In this analysis, we found that higher values were observed on seedlings, independent of the condition, than on seeds. On seeds, higher values of H2O2 were found for CMG2093 than for the other cultivars as seedlings were irrigated, and under irrigation, BRSMG Relâmpago had a higher content of hydrogen peroxide (Fig. 2). Similarly, under 70% field capacity, BRSMG Relâmpago under both conditions (irrigated and not) had a higher content, with CMG2172 being statistically similar if not irrigated (Fig. 2). It must be mentioned that under 70% field capacity, higher values of H2O2 were found. Additionally, little or no differences among cultivars and conditions were found under 10% field capacity (Fig. 2).

H2–O2 in rice seeds and seedlings from three cultivars produced with and without irrigation on conditions of 70 and 10% field capacity. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
Lipid peroxidation was measured through the content of MDA in seeds and seedlings, and we found that higher contents were found in seeds and seedlings of BRSMG Relâmpago under most conditions (Fig. 3). On the other hand, CMG2172 was the cultivar with lower values of seeds and seedlings over 70% field capacity; however, this cultivar produced higher values of MDA under 10% field capacity.

MDA quantification on rice seeds and seedlings from three cultivars produced on conditions of 70 and 10% field capacity. On X axis, “seeds” refers to the enzyme activity for seeds before emergence, and 70% and 10% refer to seedlings growing under these field capacity
Regarding SOD activity (Fig. 4), lower concentrations of this enzyme were found in seeds of CMG2172, the cultivar with lower values for both conditions of field production. Under ideal cultivation conditions, such as 70% field capacity, the produced seedlings had comparable results for both seeds from irrigated and nonirrigated fields. CMG2093 had superior activity on both comparatives regarding the contrasting material.

Superoxide dismutase enzyme activity on rice seeds and seedlings produced with and without irrigation on conditions of 70 and 10% field capacity. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
Comparing the environments, we observed a decrease in activity in both genotypes, as seen in seeds from fields without irrigation that developed into seedlings with lower SOD activity. For seedlings developed at 10% field capacity, under both conditions of an environment with irrigation, lineages CMG2172 and CMG2093 were superior to the witness, and for an environment without irrigation, the witness and genotype CMG2093 had higher concentrations.
The three genotypes showed a balance in APX (Fig. 5) under irrigated field conditions, which indicates that, under ideal developing conditions, seeds from these lineages have similar responses regarding APX enzyme activity. When genotypes from production fields irrigated or not were analyzed, a stress incidence could be found once cultivar CMG2093 had higher activity compared to the others, independent of the evaluated material.

APX activity on rice seeds and seedlings from three cultivars on production systems of 70 and 10% field capacity. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
The results for APX corroborate those for CAT (Fig. 6) observed in this research; a higher presence of one was efficient in dismantling the hydrogen peroxide produced, which resulted in lower CAT activity for the studied genotypes.

Catalase (CAT) activity on rice seeds and seedlings from three cultivars produced with and without irrigation and in different field capacity. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
By observing the results obtained by the genotypes for CAT and Fig. 7, we found a balance in dry seeds for production fields with or without irrigation, showing that seed formation and stress induction were insufficient to cause a difference in genotype composition. It is possible to see by analyzing seedlings from the emergence test at 70% field capacity, which had more CAT activity for CMG2172 and lower for CMG2093. Once both cultivars were sown under ideal water conditions, this difference between the genotypes can be attributed to the lower physiological quality of CMG2172, which had lower performance in vigor tests than the other genotypes, probably because the seeds were in an intense deterioration process, resulting in higher ROS production and consequently higher CAT expression.

Isoenzymatic pattern for (A) and activity quantification (B) catalase on rice seeds and seedlings produced under hydric stress and seedlings under two irrigation conditions. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
We could not detect this enzyme in dry seeds (Fig. 8); similarly, the absence of these enzymes could not be observed in rice through electrophoresis. Regarding seedlings produced at 70% field capacity, we observed higher expression for both fields in CMG2172, as observed for CAT. Considering the results for emergence percentage and speed, this difference between genotypes can be justified due to CMG2172 physiological quality, which resulted in higher enzyme expression aiming to mitigate ROS-caused damage (Fig. 8).

Isoenzymatic pattern (A) and activity quantification (B) for peroxidase enzyme on rice seeds and seedlings produced under hydric stress and seedlings under two irrigation conditions. On X axis, “seeds” refers to the enzyme activity for seeds before emergence tests, and 70% and 10% refer to seedlings growing under these field capacity
Regarding POX, in seedlings, the results of the emergence test at 10% field capacity were not different from those of the field with irrigation. However, seedlings from both fields, CMG2172, had higher expression, corroborating the result observed for SOD. Thus, CMG2172 is more sensitive to hydric stress incidence than the other cultivars in terms of both seed production and germination. POX is an enzyme with higher affinity to hydrogen peroxide, with an essential role in precocious detoxication due to radical accumulation; thus, as CMG2172 has lower physiological quality considering emergence tests, higher peroxidase synthesis is a response to higher cell membrane damage, as could be observed by higher MDA production.
Discussion
As the population increases, the demand for food production, such as rice, follows. Thus, breeding has contributed to improving rice production. Additionally, together with breeding, the expansion of production areas increases; however, as with any crop, rice is cultivated worldwide and sometimes out of its center of origin (Bernier et al. 2008). As breeding has evolved, new cultivars are produced, and upland rice has been researched due to its potential for production under lower water availability compared to other crops. Thus, there are a considerable number of studies on new cultivars and their tolerance to drought. However, as rice breeding continues to generate new cultivars, their physiological characteristics must be tested, including drought tolerance.
According to Guedes et al. (2011), the use of emergence percentage and speed tests are important tools for seed vigor inference once they simulate field conditions. Additionally, as mentioned by Marcos-Filho (2015) and Nakagawa (1999), vigorous seeds tend to develop faster and have increased production. As described by de Castro et al. (2014), BRS Relâmpago is a cultivar reported as drought tolerant, being a model of comparison for the results from CMG2093 and CMG2172, which, according to our results on germination, had distinct values.
In our results, CMG2093 germinated at a statistically similar rate to BRS Relâmpago, and both cultivars were superior to CMG2172, highlighting the differences among tolerant and nontolerant cultivars. Although BRS Relâmpago is drought tolerant, the production of this cultivar was reported to be lower than that of CMG2093 and CMG2172 (Castro et al. 2019). This indicates that both cultivars are of interest if production is considered; however, the environmental conditions must be taken into consideration once CMG2172 does not tolerate drought.
Among the factors of importance for drought tolerance, the antioxidant system must be considered one of the most important. Under stress, the accumulation of reactive oxygen species (ROS) increases, and these ROS are toxic, resulting in damage to the cell structure and DNA molecule radicals (You and Chan 2015; Soares et al. 2019). In this case, a vigorous seed may be able to express and maintain the antioxidant system in response to stress (Atique-ur-Rehman et al. 2020; Zhu et al. 2020a).
The understanding of the mechanisms linked to stress tolerance, such as drought, has been carried out in many studies (Wang et al. 2003; Munns et al. 2010; Blum 2011; de Abreu et al. 2014; Bakhsh and Hussain 2015). The understanding of enzymatic parameters and their differences among cultivars may be of interest in the search for faster and more accurate methods to identify cultivars regarding stress tolerance (Catão et al. 2014, 2016; Oliveira et al. 2021; Wei et al. 2021). The use of antioxidant system enzymes stands among those methods, as reported by Cavasin et al. (2023).
In our results, independent of field conditions, CMG2172 had higher protein values than the other cultivars, highlighting that those differences may be found even on the same crop and not only on different species. As we compared the results on germination, we observed that this cultivar had lower vigor than the others, indicating that higher protein content may be related to seed vigor (Chen et al. 2017; Abdul-Baki 2022). This clue is highlighted, as we observed that both CMG2172 and CMG2093 seedlings under 10% field capacity had lower vigor and higher protein content, while BRS Relâmpago, a tolerant cultivar, had superior vigor and lower protein content. Sharma and Dubey (2012) reported these differences among cultivars, which may lead to the conclusion that protein content may be a marker for seed vigor and drought tolerance.
Under stress conditions, H2O2 may accumulate in cells, causing damage to their structure (Jaleel et al. 2009), as well as MDA content, which is the final product of lipid peroxidation, both resulting from oxidative stress (Waszczak et al. 2018). However, lipid peroxidation and H2O2 content could not be correlated with vigor results for cultivars and conditions of our experiments. Li et al. (2018) reported similar results, as MDA and H2O2 may be related to other factors, and the low content may be linked to higher expression of lipoxygenase enzyme production. It is important to highlight that H2O2 cannot be considered a “villain” to plant physiology. Low amounts of this molecule function in plant signaling, activating metabolism and seed germination (Waszczak et al. 2018; Kerchev and Van Breusegem 2022).
As CMG2172 was observed to have lower vigor, lower SOD activity was also observed compared to CMG2093 for both parameters. Higher SOD values for CMG2172 and CMG2093 were found compared to BRS Relâmpago. Interestingly, as observed in our results, lower SOD activity was found under extreme (10% field capacity) conditions, which does not corroborate results from other research, such as Sahitya et al. (2018). It is important to note that SOD, as an antioxidant enzyme, is considered a marker for stress tolerance, which usually increases under these conditions (Saibi and Brini 2018; Berwal and Ram 2019).
A correlation between SOD enzyme activity and genotype ability to develop and establish under water deficit conditions was observed. Once in the vigor tests, material with higher values for emergence speed and percentage had SOD enzyme activity under either ideal or water restriction conditions. Seedlings with higher SOD activity had higher cell protection (Mullineaux and Rausch 2005); once this enzyme is the first way to control the damage caused by free radicals, it has a role in initiating detoxification by reducing superoxide radicals on hydrogen peroxide (Das and Roychoudhury 2014). Thus, SOD activity in seedlings under water deficit conditions is a positive signal regarding the plant’s ability to tolerate environmental adversity; once at low levels, superoxide radicals result in less cell oxidative damage due to their lower toxicity compared to hydrogen peroxide (Li et al. 2013). The results from this research corroborate those of et al. (2020), who observed higher development and growth in rice seedlings with higher SOD activity.
Following the route started by SOD in the antioxidant system, APX and CAT act by converting H2O2 into water and oxygen (García-Caparrós et al. 2021). We found that both enzymes follow the same pattern of results, suggesting that both can be correlated in between and with SOD results. CAT is an intracellular enzyme that acts by transforming H2O2 into a nonreactive substance and is essential to reduce damage caused by oxidative stress (Marques et al. 2019). An increase in CAT expression in vigorous seeds indicates better performance of the antioxidant system (Lacerda et al. 2003). For other crops, this increase was observed on induced hydric restriction, as observed by Hendges et al. (2015) for corn and Bortolin et al. (2020).
Higher expression of CAT was observed in seedlings from the emergence test at 10% field capacity when CMG2093 and witness were analyzed compared to seedlings at sowing conditions with controlled moisture (70% field capacity), while CMG2172 was not different, evidencing the same expression standard. This increase in hydric stress conditions on CMG2093 can be seen as a positive signal once at this point, due to cellular metabolism changes, is for adaptation to dry conditions. CAT activity is necessary to avoid damage due to ROS accumulation, and if compared to 10% field capacity conditions, CMG2093 was superior to CMG2172, reforming the theory that this increase was enough to maintain seed quality for these genotypes. Additionally, MDA decreased in CMG2093 and BRSMG Relâmpago but increased in CMG2093.
CAT and APX are enzymes responsible for detoxifying the excess hydrogen peroxide produced in cells; they are found in different isoforms and have a role in transforming H2O2 into water and molecular O2 (Sharma et al. 2012). Thus, by observing the results obtained for the genotypes for catalase, we found a balance in dry seeds for production fields with or without irrigation, showing that seed formation and stress induction were insufficient to cause a difference in genotype composition. It is possible to see by analyzing seedlings from the emergence test at 70% field capacity, which had more CAT activity for CMG2172 and lower for CMG2093. Once both cultivars were sown under ideal water conditions, this difference between the genotypes can be attributed to the lower physiological quality of CMG2172, which had lower performance in vigor tests than the other genotypes, probably because the seeds were in an intense deterioration process, resulting in higher ROS production and consequently higher CAT expression.
POX is essential for ROS detoxication, has a high affinity for H2O2, and is widely distributed in cells (Wang et al. 1999). Its expression varies according to tissue and developmental stage, and its presence varies according to vegetative growth (Vieira et al. 2008; de Tunes et al. 2014). Sohag et al. (2020) found that rice seedlings under water deficit with high APX activity had higher tolerance to induced stress. This higher APX activity in seedlings suggests a signal of tolerance to water deficit once H2O2 accumulation in different cell compartments creates a stress defense barrier, avoiding oxidative damage to fundamental organelles such as chloroplasts (Zhang et al. 2013).
D’arcy-Lamenta et al. (2006) also found this relationship in cowpea leaves, observing higher expression of APX genes aiming to mitigate hydric restriction effects. Similarly, the same results were observed in rye seeds under water restriction (Pedó et al. 2016), in which an increase in peroxidases was related to low seed vigor, corroborating the results observed here. POX expression is related to water retention capacity by leaves (Mercado et al. 2004), which can justify the lower expression on CMG2093 once it had less aerial part development. The same results were found by Zhu et al. (2020b) on cassava seedlings under different hydric conditions, in which seedlings with lower decreases in POX activity were those with less water on the aerial part.
By analyzing CAT and POX expression in both genotypes for emergence tests together with hydrogen peroxide and MDA quantification, we observed a decrease in peroxidases under hydric deficit conditions, resulting in an increase in CAT expression for the genotypes with higher vigor (CMG2093 and BRSMG Relâmpago). This increase was enough to reduce H2O2 at the membrane level, which was not observed for CMG2172, which maintained the peroxide concentration and increased MDA at 10% field capacity. The same results were found by Wang et al. (2018), who observed higher lipid peroxidation in seeds with higher deterioration due to lower antioxidant complex enzyme efficiency.
This relationship between CAT and a decrease in MDA was observed by Refli (2016) in rice seeds under hydric stress. Authors reported that higher enzyme expression and lower lipid peroxidation is due CAT origin place is peroxisomes and the negative relation between this enzyme and POX is natural.
Antioxidant complex enzymes are extremely important for the control of free radicals naturally produced in living organisms and become more relevant under extreme environmental conditions because they can detoxify ROS produced in plants (Jaleel et al. 2009). The correlation among the results on enzymatic parameters to vigor tests indicates that antioxidant system enzymes can be used as markers to indicate higher or lesser drought tolerance in rice, as was reported as interesting in another study (Vieira et al. 2008; Cavasin et al. 2023).
In conclusion, we observed lower damage by hydric stress in CMG2093, which resulted in its higher performance under stress. Finally, as distinct results on enzymatic activity could be found in cultivars, it is possible that antioxidant system activity may be used as a marker for rice drought tolerance.
Data availability
All data from this research can be made available upon request to the corresponding author.
References
Abdul-Baki AA (2022) Biochemical aspects of seed vigor. HortScience 15:765–771. https://doi.org/10.21273/hortsci.15.6.765
Alfenas AC (2006) Eletroforese e marcadores bioquímicos em plantas e microorganismos. UFV, Viçosa
Asada K (2006) Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 141:391–396
Atique-ur-Rehman KM, Afzal I (2020) Production and processing of quality cotton seed. In: Ahmad S, Hasanuzzaman M (eds) Cotton production and uses. Springer, Singapore, p 547
Bakhsh A, Hussain T (2015) Engineering crop plants against abiotic stress: current achievements and prospects. Emirates J Food Agric 27:24–39. https://doi.org/10.9755/ejfa.v27i1.17980
Basra SMA, Farooq M, Wahid A, Khan MB (2006) Rice seed invigoration by hormonal and vitamin priming. Seed Sci Technol 34:753–758. https://doi.org/10.15258/sst.2006.34.3.23
Bernier J, Atlin GN, Serraj R et al (2008) Breeding upland rice for drought resistance. J Sci Food Agric 88:927–939
Berwal MK, Ram C (2019) Superoxide dismutase: a stable biochemical marker for abiotic stress tolerance in higher plants. In: Abiotic and biotic stress in plants
Biemelt S, Keetman U, Albrecht G (1998) Re-aeration following hypoxia or anoxia leads to activation of the antioxidative defense system in roots of wheat seedlings. Plant Physiol 116:651–658. https://doi.org/10.1104/pp.116.2.651
Blum A (2011) Plant breeding for water-limited environments. Springer, New York
Bortolin GS, Salinas-Arcienega AJ, Galviz-Fajardo YC et al (2020) Seed germination and antioxidant enzyme activity in seedlings of diploid and tetraploid bahiagrass under water restriction. Ciência Rural 50:e20190382. https://doi.org/10.1590/0103-8478cr20190382
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1006/abio.1976.9999
Brasil (2009) Regras para análise de sementes. Ministério da Agricultura Pecuária e Abastecimento, Brasilia
Buege JA, Aust SD (1978) Microsomal lipid peroxidation. Methods Enzymol 52:302–310. https://doi.org/10.1016/S0076-6879(78)52032-6
Castro DG, Fernandes MCN, Félix MR et al (2019) Estimativas de associação entre caracteres agronômicos na seleção de genótipos de arroz de terras altas. Magistra 30:359–367
Catão HCRM, Gomes LAA, Dos Santos HO et al (2014) Aspectos fisiológicos e bioquímicos da germinação de sementes de alface em diferentes temperaturas. Pesqui Agropecu Bras 49:316–322. https://doi.org/10.1590/S0100-204X2014000400010
Catão HCRM, Gomes LAA, Guimarães RM et al (2016) Physiological and isozyme alterations in lettuce seeds under different conditions and storage periods. J Seed Sci 38:305–313. https://doi.org/10.1590/2317-1545v38n4163863
Cavasin PY, Dos Santos HO, Oliveira TF et al (2023) Endo-β-mannanase and superoxide dismutase as enzymatic markers for lettuce seeds thermotolerance. J Seed Sci 45:e202345008. https://doi.org/10.1590/2317-1545v45266112
Chen LT, Sun AQ, Yang M et al (2017) Relationships of wheat seed vigor with enzyme activities and gene expression related to seed germination under stress conditions. Chinese J Appl Ecol 28:609–619. https://doi.org/10.13287/j.1001-9332.201702.019
CONAB (2022) Acompanhamento da Safra Brasileira. Bol Da Safra 2021(9):60
Czarnocka W, Karpiński S (2018) Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic Biol Med 122:4–20. https://doi.org/10.1016/j.freeradbiomed.2018.01.011
D’Arcy-Lameta A, Ferrari-Iliou R, Contour-Ansel D et al (2006) Isolation and characterization of four ascorbate peroxidase cDNAs responsive to water deficit in cowpea leaves. Ann Bot 97:133–140. https://doi.org/10.1093/aob/mcj010
Das K, Roychoudhury A (2014) Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front Environ Sci 2:1–13. https://doi.org/10.3389/fenvs.2014.00053
De Castro AP, De Morais OP, Breseghello F et al (2014) BRS Esmeralda: cultivar de arroz de terras altas com elevada produtividade e maior tolerância à seca. Embrapa Arroz e Feijão-Comunicado Técnico 2015:1–4
de Abreu VM, Von Pinho ÉV, de R, Von Pinho RG, et al (2014) Desempenho fisiológico e expressão de isoenzimas em sementes de milho submetidas ao estresse hídrico. J Seed Sci 36:40–47. https://doi.org/10.1590/S2317-15372014000100005
de Lacerda AL, de Sá ME, Valério Filho WV (2003) Armazenamento de sementes de soja dessecadas e avaliação da qualidade fisiológica, bioquímica e sanitária. Rev Bras Sementes 25:97–105. https://doi.org/10.1590/s0101-31222003000400014
de Oliveira DF, Cavasin PY, Silva S et al (2021) Genetic control of thermoinhibition tolerance in lettuce seeds. Pesqui Agropecuária Bras 56:e02337. https://doi.org/10.1590/s1678-3921.pab2021.v56.02337
de Tunes LVM, Fonseca DÂR, Meneghello GE et al (2014) Qualidade fisiológica, sanitária e enzimática de sementes de arroz irrigado recobertas com silício. Rev Ceres 61:675–685. https://doi.org/10.1590/0034-737X201461050011
Dhindsa RS, Plumb-dhindsa P, Thorpe TA (1981) Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:93–101. https://doi.org/10.1093/jxb/32.1.93
FAO (2019) FAOSTAT database: agriculture production. In: Food Agric Organ United Nations. http://www.fao.org/home/en/
Fridavich I (1995) Superoxide radical and superoxide dismutases. Annu Rev Biochem 64:97–112. https://doi.org/10.1146/annurev.bi.64.070195.000525
García-Caparrós P, De Filippis L, Gul A et al (2021) Oxidative stress and antioxidant metabolism under adverse environmental conditions: a review. Bot Rev 87:421–466. https://doi.org/10.1007/s12229-020-09231-1
Giannopolits CN, Ries SK (1977a) Superoxide dismutases: I. Occurr Higher Plants Plant Physiol 59:309–314. https://doi.org/10.1104/pp.59.2.309
Giannopolits CN, Ries SK (1977b) Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol 59:315–318. https://doi.org/10.1104/pp.59.2.315
Guedes RS, Alves EU, De Oliveira LSB et al (2011) Envelhecimento acelerado na avaliação da qualidade fisiológia de sementes de Dalbergia nigra (Vell.) Fr. All Semin Agrar 32:443–450. https://doi.org/10.5433/1679-0359.2011v32n2p443
Havir EA, McHale NA (1987) Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol 84:450–455. https://doi.org/10.1104/pp.84.2.450
Hendges FB, Rambo CR, Alcassa LP et al (2015) Avaliação enzimática e fisiológica de plântulas de milho submetidas à seca. Rev Bras Energias Renov. https://doi.org/10.5380/rber.v4i2.42287
Jaleel CA, Riadh K, Gopi R et al (2009) Antioxidant defense responses: physiological plasticity in higher plants under abiotic constraints. Acta Physiol Plant 31:427–436
Kerchev PI, Van Breusegem F (2022) Improving oxidative stress resilience in plants. Plant J 109:359–372. https://doi.org/10.1111/tpj.15493
Li CR, Liang DD, Li J et al (2013) Unraveling mitochondrial retrograde regulation in the abiotic stress induction of rice ALTERNATIVE OXIDASE 1 genes. Plant Cell Environ 36:775–788. https://doi.org/10.1111/pce.12013
Liu L, Xia W, Li H et al (2018) Salinity inhibits rice seed germination by reducing α-amylase activity via decreased bioactive gibberellin content. Front Plant Sci 9:275. https://doi.org/10.3389/fpls.2018.00275
Maguire JD (1962) Speed of germination—aid in selection and evaluation for seedling emergence and vigor. Crop Sci 2:176–177. https://doi.org/10.2135/cropsci1962.0011183x000200020033x
Marcos Filho J (2015) Fisiologia de sementes de plantas cultivadas
Marques TL, Von Pinho RG, De Von Pinho ÉVR, Dos Santos HO (2019) Expression of zmLEA3, AOX2 and zmPP2C genes in maize lines associated with tolerance to water deficit. Cienc e Agrotecnologia 43:e022519. https://doi.org/10.1590/1413-7054201943022519
Mercado JA, Matas AJ, Heredia A et al (2004) Changes in the water binding characteristics of the cell walls from transgenic Nicotiana tabacum leaves with enhanced levels of peroxidase activity. Physiol Plant 122:504–512. https://doi.org/10.1111/j.1399-3054.2004.00429.x
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9:490–498
Mullineaux PM, Rausch T (2005) Glutathione, photosynthesis and the redox regulation of stress-responsive gene expression. Photosynth Res 86:459–474. https://doi.org/10.1007/s11120-005-8811-8
Munns R, James RA, Sirault XRR et al (2010) New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J Exp Bot 61:3499–3507. https://doi.org/10.1093/jxb/erq199
Nakagawa J (1999) Testes de vigor baseados no desempenho das plântulas. Vigor Sementes Conceitos e Testes Londrina Abrates 1:1–24
Pedó T, Koch F, Gazolla A et al (2016) Expressão isoenzimática e do vigor de sementes de centeio sob efeito da restrição hídrica. Pesqui Agropecuária Pernambucana 21:17–23. https://doi.org/10.12661/pap.2016.004
R_Core_Team (2021) R: A Language and Environment for Statistical Computing. RFoundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Refli YAP (2016) The response of antioxidant genes in rice (Oryza sativa L.) seedling Cv. Cempo Ireng under drought and salinity stresses. In: AIP conference proceedings. American Institute of Physics Inc., p 20063
Saibi W, Brini F (2018) Superoxide dismutase (SOD) and abiotic stress tolerance in plants: an overview. In: Superoxide dismutase: structure, synthesis and applications. pp 101–142
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:1–26. https://doi.org/10.1155/2012/217037
Sies H, Cadenas E (1985) Oxidative stress: damage to intact cells and organs. Philos Trans R Soc London B, Biol Sci 311:617–631
Soares CM, Ludwig MP, Soares Rother CM et al (2019) Seed quality and crop performance of soybeans submitted to different forms of treatment and seed size. J Seed Sci 41:86–96. https://doi.org/10.1590/2317-1545v41n1213725
Sohag AAM, Tahjib-Ul-Arif M, Brestič M et al (2020) Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil Environ 66:7–13. https://doi.org/10.17221/472/2019-PSE
Taiz L, Zeiger E (2013) Fisiologia vegetal. Porto Alegre
Vieira AR, Oliveira JA, Guimarães RM et al (2008) Marcador isoenzimático de dormência em sementes de arroz. Rev Bras Sementes 30:81–89. https://doi.org/10.1590/s0101-31222008000100011
Wang J, Zhang H, Allen RD (1999) Overexpression of an arabidopsis peroxisomal ascorbate peroxidase gene in tobacco increases protection against oxidative stress. Plant Cell Physiol 40:725–732
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: toward genetic engineering for stress tolerance. Planta 218:1–14
Wang W, He A, Peng S et al (2018) The effect of storage condition and duration on the deterioration of primed rice seeds. Front Plant Sci 9:1–17. https://doi.org/10.3389/fpls.2018.00172
Waszczak C, Carmody M, Kangasjärvi J (2018) Reactive oxygen species in plant signaling. Annu Rev Plant Biol 69:209–236
Wei S, Zhang L, Huo G et al (2021) Comparative transcriptomics and metabolomics analyses provide insights into thermal resistance in lettuce (Lactuca sativa L.). Sci Hortic (amsterdam) 289:110423. https://doi.org/10.1016/j.scienta.2021.110423
You J, Chan Z (2015) Ros regulation during abiotic stress responses in crop plants. Front Plant Sci 6:1092
Zandalinas SI, Mittler R, Balfagón D et al (2018) Plant adaptations to the combination of drought and high temperatures. Physiol Plant 162:2–12
Zhang Z, Zhang Q, Wu J et al (2013) Gene knockout study reveals that cytosolic ascorbate peroxidase 2(OsAPX2) plays a critical role in growth and reproduction in rice under drought. PLoS One, Salt and Cold Stresses. https://doi.org/10.1371/journal.pone.0057472
Zhu M, Yu X, Zhao G, Wang L (2020a) Effects of harvest time on seed vigor, enzyme activity and gene expression of conventional japonica rice. Arch Agron Soil Sci 11:214. https://doi.org/10.1080/03650340.2020.1840558
Zhu Y, Luo X, Nawaz G et al (2020b) Physiological and biochemical responses of four cassava cultivars to drought stress. Sci Rep. https://doi.org/10.1038/s41598-020-63809-8
Acknowledgements
The authors thank the research promotional agencies Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES – Brasil), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG – Brasil), and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (Process 426309/2018-9). HOS is a productivity fellow from CNPq (Process 310211/2021-2).
Funding
This research was supported by Brazilian research funding agencies Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG).
Author information
Authors and Affiliations
Contributions
GVT, HOS, and FBSB conceived the idea and designed the methodology. GVT, GJF, and ALR collected, analyzed, and interpreted the data. GVT, HOS, FBSB, and WVSP wrote and edited the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
To the knowledge of the authors, no competing interest, financial or nonfinancial, is issued on this research.
Additional information
Communicated by Mian Abdur Rehman Arif.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Tirelli, G.V., dos Santos, H.O., Frota, G.J. et al. Physiological and biochemical performance of hydric stress-produced rice seeds. CEREAL RESEARCH COMMUNICATIONS (2023). https://doi.org/10.1007/s42976-023-00460-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42976-023-00460-9