
Abstract
In response to the ongoing expansion of agriculture and land use changes in the Amazon region, new studies are necessary to understand the impacts of agricultural practices on Amazonian soils. The objective was to assess the changes induced in soil horizons by different management practices. The research was conducted in the National Forest of Caxiuanã, located in the municipality of Melgaço, Pará State, Brazil. A completely randomized 3 × 4 factorial design was employed, incorporating three types of vegetation (20-year-old secondary forest—SoilFS20, 5-year-old secondary forest—SoilFS5, and coivara—cutting and burning of native vegetation—SoilLTM) and four effective soil depths (0–5 cm, 5–10 cm, 10–20 cm, and 20–30 cm), with nine replicates for each combination. To enhance accuracy of the required assessment of the modifications induced in the physical and chemical attributes of the soil, isotopic composition of δ13C and δ15N was utilized. In areas subjected to clearing (SoilFS5 and SoilLTM), significant concentrations of carbon (C) and nitrogen (N) were observed in the 20–30 cm depth due to soil disturbance, which incorporated the litter into the deeper horizons. However, no significant differences in the C/N ratio were detected among the studied areas at the effective soil depth. Regardless of soil depth, SoilFS20 exhibited lower acidity, δ13C, δ15N, sandy and clay content, as well as iron (Fe) content, while displaying higher levels of SOM, zinc (Zn), calcium (Ca), manganese (Mn), potassium (K), silt, sodium (Na), copper (Cu), and phosphorus (P) content compared to the other sites. The results showed significant variations in the chemical and physical properties of the soil among the study sites, with SoilFS20 standing out in several characteristics. The increase in carbon stock in SoilFS20 indicated positive responses to specific management practices. Analyses of carbon, nitrogen concentrations, and stable isotopes highlighted SoilFS5 as distinctly different from the others. These findings contribute to the understanding of soil dynamics under different management practices, providing valuable insights for sustainable natural resource management in the region.
Similar content being viewed by others
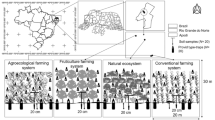

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Deforestation resulting from the expansion of agricultural areas and the practice of clearing land for timber production, agriculture, or charcoal represents a global challenge with significant implications for the terrestrial carbon cycle (Ray et al. 2023; Reichenbach et al. 2023). Land use plays a crucial role in climate change, significantly influencing the dynamics of soil carbon (C) and nitrogen (N) in terrestrial ecosystems (Batlle-Bayer et al. 2010; Cotrufo et al. 2011; Lu et al. 2014). Human activities have caused substantial environmental damage, resulting in rapid losses of C and N from the soil (Dou et al. 2016). Land use changes, including deforestation, agricultural crop management, and animal husbandry, directly impact the dynamics of C and N. The carbon cycle has drawn the interest of society as a whole due to the growing concern regarding ongoing climate changes (Mantovani et al. 2024).
In recent years, the expansion of agriculture in the Amazon region has accelerated, leading to deforestation of native areas and alterations in land use, thereby causing environmental damage to the dynamics of C and N and disrupting biological biodiversity (Almeida et al. 2016; Arvor et al. 2017). Oxisols and Ultisols, which account for 80% of the Brazilian Amazon's territory, are acidic soils with low natural chemical fertility, deep profiles, and color ranging from red to yellow (Souza et al. 2018). Additionally, historical agricultural practices, such as itinerant cultivation (burning, cultivating, harvesting, and abandoning), have contributed to the rapid anthropization of these soils (Montoya et al. 2020). Studies have shown that the outcomes of soil modifications in early agriculture can range from improvement to degradation, depending on various factors such as agricultural methods, cultivation intensity, and environmental sensitivity (Araujo et al. 2023; Satdichanh et al. 2023).
Tropical soils play a vital role in biological diversity and the global carbon cycle (Wu et al. 2024), representing 13% of all carbon storage in the world's soil (Post et al. 1982). In addition, Amazon C storage represents 62% of the total C of Brazilian soils (Cerri et al. 2008), with approximately half of it estimated to be stored in the topsoil (up to 20 cm) (Moraes et al. 1995). However, agricultural expansion, driving deforestation rates in the Amazon, exacerbates climate change and leads to poor soil quality (Christina et al. 2017). Forest-to-agricultural land conversion has been identified as the primary cause of organic carbon loss in tropical regions (Don et al. 2011). Deforestation and biomass burning accelerate the decomposition of soil organic matter (SOM), resulting in increased CO2 emissions, amplifying the greenhouse effect and contributing to global warming (Batjes 2006). Furthermore, these activities eradicate microbial biomass on the soil surface, affecting soil biochemical processes (Cerri et al. 2008). Consequently, rapid changes in the biogeochemical cycle significantly impact carbon storage (Wu et al. 2024), underscoring the need for extensive scientific research on soil carbon storage in the Brazilian Amazon (Araujo et al. 2023).
SOM undergoes changes in response to climate and land use alterations, as well as soil types, influencing carbon input and output rates and soil decomposition rates (Falloon et al. 2007). Lower microbial activity in deeper soil horizons leads to reduced decomposition of root biomass and, consequently, lower organic carbon levels (Lal 2008). However, soils with higher clay content exhibit higher carbon storage levels at greater depths due to their enhanced capacity to translocate and store elements (Zinn et al. 2002). Nevertheless, most studies focus primarily on surface horizons when analyzing organic carbon, as accumulation predominantly occurs in the topsoil (Wiesmeier et al. 2012). Therefore, it remains unclear whether carbon sinks also occur at greater depths (Kalbitz and Kaiser 2008).
The management practices historically employed by farmers in the Caxiuanã National Forest, a Conservation Unit, differ from the traditional slash-and-burn practices found in the tropics. The practice known as "slash and char" involves controlled biomass burning, resulting in the production of charcoal instead of ash. This incomplete combustion of biomass has given rise to the Amazonian Dark Earths, locally known as "Terra Preta de Índio" (SOILFS20), which are fertile anthropogenic soils found in the Amazon region. These soils are characterized by high levels of macro and micronutrients, cation exchange capacity (CEC), pH value, base saturation, and up to 70 times more carbon content compared to Oxisols (Glaser et al. 2000).
Considering that these sites often exhibit past agricultural activities that contribute to the formation of these anthropogenic soils, the use of isotopic techniques for the analysis of essential elements such as carbon and nitrogen becomes an interesting alternative. Therefore, understanding the influence of these soil management practices is of particular importance, especially in archaeological sites where SOILFS20 is present. Such studies are relevant not only for comprehending the process of formation of these anthropogenic soils but also for providing insights that enable better conservation practices through sustainable agriculture.
SOM is considered a valuable indicator of changes in land use and soil quality (Zhang et al. 2024). A positive correlation in the soil organic pool indicates appropriate and sustainable land use (Obalum et al. 2017). Furthermore, the isotopic composition of SOM exhibits minimal variation, making it possible to use natural abundances of carbon-13 (δ13C) and nitrogen-15 (δ15N) to identify the source of organic matter before deforestation (Vitorello et al. 1989). Moreover, δ15N values tend to increase with depth in soils as inorganic nitrogen storage becomes relatively greater compared to organic nitrogen.
In this context, it is crucial to investigate whether agricultural practices have the potential to induce significant changes in the dynamics of soil organic matter (SOM) and nutrients. Our assumptions suggest that a short resting period (~ 5 years) for ecological regeneration may not be sufficient to enhance SOM, necessitating longer periods (~ 20 years) to effectively restore SOM formation and improve soil fertility. Thus, there is a need for studies to understand how the management practices in the Caxiuanã National Forest impact the global carbon cycle. By examining physical and chemical attributes, stable isotopic composition (δ13C and δ15N), carbon content and storage, carbon-to-nitrogen (C/N) ratio, this study aims to evaluate the level of anthropization and the origin of organic matter in soil horizons of three sites designated for agricultural use in the Brazilian Amazon.
2 Material and Methods
2.1 Study Area
The study was conducted in the Caxiuanã National Forest, a Conservation Unit located in Pará State, Brazil. The study site covers an area of 324 hectares, divided proportionally among the three study conditions: SOILFS20, SOILFS5, and SOILLTM. Each area represents a specific soil type and vegetation cover. Additionally, soil samples were collected from four different horizons of soil, including 0–5 cm, 5–10 cm, 10–20 cm, and 20–30 cm depths.
This study focused on analyzing soil sites located within the Caxiuanã National Forest, characterized by distinct soil types and land use histories. The first site, labeled as SoilFS20 (01º40′43.5" S and 51º 20′07.1" W), is an anthropogenic hypereutrophic yellow Ultisol with a sandy/medium texture. It was previously utilized for agriculture but has since been abandoned for 20 years, allowing for natural regeneration under a secondary forest. The second site, SoilFS5 (01º47′06.5" S e 51º28′36.0"W), shares similar characteristics with SoilFS20, but has a shorter period of abandonment, with only five years of secondary forest regeneration after cassava cultivation. Lastly, the third site, RCSoilLTM (01º47′06.5" S and 51º28′36.0" W), represents a typical hypereutrophic yellow Ultisol with a clayey/medium texture. This soil is subjected to temporary planting of various crops such as cassava, pumpkin, gherkin, corn, and watermelon, utilizing the "coivara" system, which involves slash and burn practices and the use of ashes as fertilizer. In this kind of cultivation de coivara, the preparing of area consists of slash and burn practice, followed by crops rotation and usage of the ashes as a fertilize.
2.2 Treatments and Soil Sampling
The experimental design followed a completely randomized block design with three plots per site (each measuring 3 × 4 m) and three sampling points per plot, resulting in a total of nine replications. The treatments included the three soil sites (SOILFS20, SOILFS5, and SOILLTM) and four soil horizons (0–5 cm, 5–10 cm, 10–20 cm, and 20–30 cm depths). This design yielded a total of 108 soil samples. The soil samples were air dried in a forced circulation oven at 60 °C until a constant mass was achieved and then sieved to a particle size of less than 2 mm.
2.3 Soil Chemical and Physical Analysis
Soil chemical and physical analyses were conducted following the methods described by Raij et al. (1997) at the Laboratory of Soil and Tissue Analysis in Belem, Pará State. The pH of the soil was determined using a 0.01 mol CaCl2 solution with a soil-to-solution ratio of 1:2.5 (w/v). Various elements, including P, K, Mn, Fe, Zn, Cu, and Na, were extracted using the Mehlich 1 method. The extraction of Ca, Mg, and Al was performed using 1 M KCl solution. The H + Al extraction was carried out using a 0.5 M calcium acetate solution, while B extraction utilized hot water and S extraction employed monocalcium phosphate in acetic acid. Based on the obtained results, the sum of bases (SB), cation exchange capacity (CEC at pH 7.0), base saturation (V), and organic matter (C = Org C × 1.724) were calculated.
To determine the clay, silt, and sand content in the soils, the technique of granulometric analysis was used. Initially, representative soil samples were collected at each study site. The samples were air-dried to remove moisture and then passed through a coarse sieve to remove debris and larger fragments. Subsequently, approximately 100 g of each sample were weighed and placed in dispersion containers along with an appropriate dispersing solution. These containers were vigorously agitated for a specific period to disperse the soil particles. After dispersion, the containers were filled with water to the brim and left undisturbed for a specific time to allow particle sedimentation. During this period, separation of particles occurs based on their size and density. After the settling time, aliquots of suspended water from the top of the containers, containing the clay, silt, and sand fractions, were extracted. These aliquots were transferred to evaporation containers and placed in an oven at a controlled temperature for complete drying. Once dried, the fractions of clay, silt, and sand were weighed, and the percentage content of each fraction was calculated relative to the initial sample mass. Standardized formulas based on the relative proportions of the obtained fractions were used to calculate the clay, silt, and sand content.
2.4 Determination of Soil Organic Matter (SOM)
The determination of soil organic matter (SOM) amount was based on the principle of measuring the easily oxidizable or decomposable SOM in the soil (Walkley and Black 1934). With the determination of the soil organic carbon (SOC) content, the conversion to SOM was performed by the Van Bemmlen factor (1.724), considering that the SOM of the soil contains 58% of organic C. The percentage of organic C in the soil is given by the Eq. (1).
SOC = soil organic carbon (g kg−1); V = volume of potassium bichromate used; Va = volume of ammoniacal ferrous sulfate used in sample titration; f = 40/volume of ammoniacal ferrous sulfate spent in the blank titration; 0.06 = correction factor; “f” = correction factor. The amount of SOM in the sample is calculated by the expression: SOM (g kg−1) = SOC × 1.724.
2.5 C Stock
The C stock (Mg ha−1) of each area was estimated using the SOC (g kg−1), soil density (g m−3) and depth (m), using the Eq. (2) in Bernoux et al. (1998):
S = Soil C stock (Mg ha−1); Ds = Soil bulk density (g m−3); h = 0.05 (m); C = Soil C content (g kg−1).
The soil density was determined by collecting undisturbed soil samples (100 cm3) in volumetric rings with the aid of an undisturbed soil sampler. After collecting the soil samples, they were dried in an oven (105 °C) to determine the dry mass. The soil density was obtained by the ratio of dry mass to the collected volume (100 cm3).
2.6 Isotopic Analysis of C and N
The preparation and analysis of the samples were performed by weighing in tin capsules, containing 6 mg of soil from the depths 0–5 cm and 10–20 cm and 10 mg of soil from the depths 5–10 cm and 20–30 cm. These samples were introduced into an elemental analyzer (Carlo Erba EA 1110 CHNS, CE Instruments). Determination of the N and C concentrations was performed through the combustion of samples, where the gas from the combustion is put in a flow of helium through a chromatographic column and introduced directly into a mass spectrometer for isotopic ratios (Delta Plus, Thermo Quest – Finnigan). The natural abundance of 15N and 13C is expressed as deviations per thousand (‰) using the Eq. 3 (1999) (Martinelli et al. 1999).
The determination of δ15N was achieved through the Eq. (3) below.
RSample: sample isotopic ration 15N/14N; Ratm: pattern isotopic ration 15N/14N. Pattern is the abundance of 15N in the atmosphere.
The determination of δ13C was performed through the Eq. (4) below:
RSample: sample isotopic ration 13C/12C; \({R}_{pdb}\): pattern isotopic ration 13C/12C. The pattern is the limestone from the Pee Dee Belemite – PDB formation.
2.7 Data Analysis
The collected data were subjected to analysis of variance (ANOVA) using the F test at a significance level of p < 0.05. Significant differences among means were determined using the Tukey test at a significance level of p < 0.05. Pearson correlation analyses were conducted to evaluate the relationships between soil attributes. Additionally, the data were analyzed using principal component analysis (PCA) and cluster analysis to characterize the variables within each site and soil depth and group them based on similarity. The statistical analyses were performed using the SAS version package, and the graphical representation of the data was generated using SigmaPlot 11.0.
3 Results
3.1 Soil Organic Matter and Carbon stock
The carbon (C) stock and soil organic matter (SOM) content increased in accordance with the lower land use intensity, with SoilLTM > SoilFS5 > SoilFS20. Notably, there was a significant increase in the C stock of SoilFS20 compared to SoilFS5 and SoilLTM (Fig. 1). Specifically, the C stock in SoilFS20 showed a 40% and 120% increase relative to SoilFS5 and SoilLTM, respectively (Fig. 1a). Moreover, the SOM content in the SoilFS20 site exhibited a 25% and 83% increase compared to SoilFS5 and SoilLTM, respectively. The SOM content decreased with increasing soil depth, reaching the lowest values in the 20–30 cm horizon.

Carbon storage (A) and soil organic matter at different soil depth (B) in SoilFS20 (CP), SoilFS5 (TPI) and SoilLTM (RC) sites at Caxiuanã National Forest—Brazilian Amazon. Lowercase letters differ among agricultural practices (CP, TPI, and RC) by the Tukey test (p < 0.05)
3.2 Total Content of C and N, Stable Isotopes Composition (δ13C and δ15N) and C/N Ratio
A significant increase was observed in the total C content (Fig. 2a), N content (Fig. 2b), δ13C (Fig. 2c), and δ15N (Fig. 2d) of SoilFS5 compared to SoilFS20 and SoilLTM. However, there were no significant differences between SoilFS20 and SoilLTM (p > 0.05). The mean total C and N content of SoilFS5 were 5% and 12% higher than SoilFS20 and 10% and 20% higher than SoilLTM, respectively. Additionally, the δ13C values were highest in SoilFS5 (-26.06 ‰). When considering soil depth (0–30 cm), differences were observed in the 20–30 cm horizon, which exhibited the highest total C content and δ13C values in the SoilFS5 site (5.03 and -24.42 ‰, respectively).

Carbon (A) and Nitrogen (B) (C and N) content, stable isotopes composition of C (C) and N (D) and C/N ratio (E) in soil horizons of SoilFS20 (CP), SoilFS5 (TPI) and SoilLTM (RC) sites at Caxiuanã National Forest—Brazilian Amazon. Lowercase letters differ among agricultural practices (CP, TPI, and RC) by the Tukey test (p < 0.05)
The N content and δ15N values followed the order SoilFS20 > SoilLTM > SoilFS5, with SoilFS5 showing the highest mean N content (8.43 g kg−1). Concerning soil depth, the δ15N values decreased with increasing depth, reaching the lowest values in the 20–30 cm horizon. There were no significant differences in the C/N ratio between the studied areas within the effective soil depth. However, the C/N ratio differed between the 0–5 cm horizon of SoilFS5 and SoilLTM, as well as the 20–30 cm horizon of SoilFS20 and SoilFS5, with a mean value of 15.3.
3.3 Characterization among Sites and Depths
The specific correlation among micro and macronutrients and other variables can be observed in Fig. 1. We found a positive Pearson’s correlation between δ13C and N content, and δ15N with C content and SOM. Furthermore, the clay was negatively correlated to δ15N, C content, SOM and silt. The total variance was explained by 73% (Principal component 1 + 2), with PC1 being explained by δ13C, silt, K, Na, Ca, Cu, Mn and Zn, while PC2 was explained by δ15N, Mg and Fe. The parameters N content, C content, C/N ratio, SOM, sandy, clay, P and B contributed with average values to explain the variance in PC1 and PC2. Under SoilFS20, regardless soil depths, the samples were characterized by higher values of SOM, Zn, Ca, Mn, K, silt, Na, Cu and P and lower values of sandy, clay, Fe, δ13C and δ15N. Soil samples under lower SoilFS5 sites showed the opposite of SoilFS20, which means higher sandy, clay, Fe, δ13C and δ15N. The SoilLTM site showed average values among the SoilFS20 and SOILFS5.
The cluster analysis indicates the formation of two groups among soil sites: the group 1 is formed by SoilFS20 site, and group 2 that contains SoilFS5 and SoilLTM sites, regardless soil depths.
4 Discussion
Soil attributes play a crucial role in unraveling the complexities of nutrient cycling and fertility dynamics. Particularly, the values of sum of bases (SB) and carbon (C) in the surface horizon (0–30 cm) are indicative of enhanced nutrient cycling, facilitated by root activity and litter decomposition (Fig. 1). However, the inherent low chemical fertility of tropical soils, attributed to the prevalence of low-reactivity silicate minerals and limited cation exchange capacity (CEC) (Lotse Tedontsah et al. 2023; Nel et al. 2023), poses a challenge. Fortunately, the formation of humus in Amazonian soils introduces negative charges within the CEC, enhancing the soil's ability to retain base cations (Aprile 2014; Chertov and Nadporozhskaya 2018). SoilFS20, among the investigated soil types, demonstrates the most favorable conditions for soil organic matter (SOM) formation, owing to the continuous influx of organic residues maintaining a balance between humus formation and decomposition.
The increase in SOM content significantly impacts other soil chemical attributes, influencing the soil's capacity to retain basic cations and facilitating Al3+ complexation (Singh et al. 2018). Additionally, the rise in SOM can lead to reduced H+ activity through the release of metallic cations, organic nitrogen mineralization, denitrification, and decarboxylation of organic acids (Angst et al. 2021). The extended regeneration period in our study area directly influences SOM accumulation, altering SB, Al3+, and base saturation (V%).
Accumulation of SOM is contingent upon the rate of organic residue deposition and decomposition. The elevated carbon content in SoilFS20 is attributed to the presence of primary and dense forests in flat terrain (Araújo et al. 2011). Despite occasional waterlogging in the fallow SoilFS20 site, sedimentation and organic matter decomposition augment carbon and phosphorus contents. In contrast, higher aluminum (Al) values observed in SoilFSS5 and SoilLTM (Fig. 3) may result from forested areas conversion to agricultural fields, leading to higher pH values (Araújo et al. 2011). The elevated Al and pH values confirm incomplete biomass combustion, resulting in the production of charcoal instead of ash.

Pearson correlation in soil horizons of SoilFS20 (CP), SoilFS5 (TPI) and SoilLTM (RC) sites at Caxiuanã National Forest—Brazilian Amazon. δ15N: stable nitrogen isotopes; δ13C: stable carbon isotopes; SOM: Soil organic matter. N: nitrogen concentration; C: carbon concentration; P: phosphorus concentration; Na: sodium concentration; Ca: calcium concentration; Mg: magnesium concentration; B: boron concentration; Cu: copper concentration; Fe: iron concentration; Mn; manganese concentration; Zn: zinc concentration. A correlation index ranging from 1 to -1, indicating a positive relationship at 1 and a negative relationship at -1 between the variables. *Significant correlation by F-test (p < 0.05)
The high silt content in SoilFS20 is attributed to the sedimentary origin of the parent material (claystone and siltstone) and past arid climatic conditions, with contributions from gypsum veins and carbonate materials gypsum veins and carbonate materials (Araújo et al. 2011). It is important to note that soil use does not directly alter soil granulometry, as it primarily depends on the original soil formation material. However, these attributes significantly impact SOM dynamics, influencing pH, SB, and V%. Soil granulometry directly affects SOM stability due to the role of clay in aggregate formation, subsequently impacting microbial accessibility, enzymatic activity, and O2 diffusion within the micropores (Cotrufo and Lavallee 2022).
Consequently, our results underscore the pivotal role of SOM in modulating the dynamics of soil chemical attributes, including SB, pH, Al3+, and V%. They emphasize the significance of SOM in the regeneration of anthropogenic dark earth, even in soils with inherently favorable chemical fertility. However, anthropogenic soil use can jeopardize these beneficial attributes. Therefore, it is imperative to implement practices that promote SOM formation and simultaneously reduce its degradation. Our study highlights that SOM content increases with the duration of the regeneration cycle, leading to significant alterations in both the chemical and granulometric attributes of the soil.
These findings emphasize the crucial importance of prioritizing and enhancing SOM in soil management strategies. The sustainable regeneration of anthropogenic dark earth relies on practices that promote the formation of SOM while minimizing its depletion. By focusing on SOM enrichment, soil fertility can be effectively restored and maintained, even in soils with initially favorable chemical properties.
Our study provides valuable insights into the complex interplay between soil attributes and SOM dynamics. It underscores the need for a comprehensive understanding of the factors influencing SOM accumulation and its impact on soil chemistry. These findings have practical implications for land managers, farmers, and policymakers involved in soil conservation and sustainable agriculture.
The content of SOM obtained in this study ranged from 10.38 to 46.44 g kg−1 (Fig. 1b), indicating medium levels of SOM. Similar results were reported by Lima et al. (2012) in the same areas, with values ranging from 22.24 to 25.79 g kg−1in tropical conditions in the Amazon (Lima et al. 2012). In contrast, Silva et al. (2012) found different SOM contents in anthropic soil in the south of Pará State, with values as high as 124.12 g kg−1, and a reduction ranging from 118.95 to 18.96 g kg−1 in the surface and 5 cm deep horizons (Silva et al. 2012).
The variations in SOM contents among the different areas indicate the significant influence of management systems and time on SOM production in the soil (Tyopine et al. 2022; Uzoh et al. 2020). Higher levels of SOM contribute to increased stability of soil aggregates, enhanced soil water retention and infiltration capacity, as well as reduced erosion and leaching (Obalum et al. 2012; Tyopine et al. 2020). Therefore, the content of SOM is of utmost importance in assessing soil quality and management practices. In this regard, the SoilFS20 site demonstrated better soil quality and management practices compared to SoilFS5 and SoilLTM.
Despite the highest accumulation of SOM in SoilFS20 (Fig. 3), this value is considered average due to the soil granulometry, as the studied soil is characterized by a medium sandy texture (Ramalho et al. 2022). The low clay content in the soil directly affects the protective mechanism of SOM. Soils with higher clay proportions provide more surfaces for C retention, resulting in increased soil C stock (Churchman et al. 2020).
According to Araujo et al. (2011), forests, such as the SoilFS20 site, exhibit higher C stocks in the 0–40 cm horizon compared to annual agriculture crops. This is attributed to the higher decomposition of root biomass and microbiome activity, which enhance the biogeochemical cycle and organic C levels (Araujo et al. 2011; Boddey et al. 2015). The C stock in agriculture crops is directly influenced by the time elapsed since the conversion from forest through slash and burn processes. Pastures with a history of 10 years showed higher C stocks compared to those with only three years, reflecting the lower C input during the initial stages of crop establishment (Araujo et al. 2011). Thus, the C stock serves as an indicator of sustainable land use and soil quality, as observed in the SoilFS20 site along with its physical and chemical soil attributes (Fig. 3).
The principal component analysis (PCA) revealed a positive association between SOM and nutrients in the surface soil horizons (0–0.5 m) through component 1 (Fig. 4). This correlation can be explained by the generation of negative charges on SOM from the dissociation of hydrogen in hydroxyl groups (Cotrufo and Lavallee 2022). The Pearson correlation analysis (Fig. 3) further confirmed the positive correlation between SOM and nutrients such as phosphorus (P), potassium (K), calcium (Ca), manganese (Mn), and zinc (Zn). These results emphasize the role of SOM in nutrient cycling, which in turn promotes increased biomass production in plants and the entry of organic waste through plant senescence at the end of their life cycle.

Principal component analysis in soil horizons of the SoilFS20 (CP), SoilFS5 (TPI), and SoilLTM (RC) sites at three depths (0–5, 5–10, 10–20, and 20–30 cm) in the Caxiuanã National Forest—Brazilian Amazon. δ15N: stable nitrogen isotopes; δ13C: stable carbon isotopes; SOM: Soil organic matter. N: nitrogen concentration; C: carbon concentration; P: phosphorus concentration; Na: sodium concentration; Ca: calcium concentration; Mg: magnesium concentration; B: boron concentration; Cu: copper concentration; Fe: iron concentration; Mn; manganese concentration; Zn: zinc concentration
The formation of SOM in anthropogenic dark earth is influenced by land use, indicating that agricultural cultivation practices disrupt the balance between SOM formation and destruction. Furthermore, the findings of this study suggest that a period of five years of regeneration can alter SOM dynamics, leading to increased accumulation. However, the 20-year period showed even higher rates of SOM accumulation. These results indicate that the equilibrium of SOM in the ecosystem has not yet been reached, highlighting the need for further studies with longer observation periods to fully understand the long-term impact of agriculture on SOM dynamics (Fig. 5).

Cluster analysis in soil horizons of the SoilFS20 (CP), SoilFS5 (TPI), and SoilLTM (RC) sites at three depths (0–5, 5–10, 10–20, and 20–30 cm) in the Caxiuanã National Forest—Brazilian Amazon. The red line indicates the formation of two groups from hierarchical clustering analysis
Moreover, our study underscores the importance of regeneration processes in restoring the balance of SOM in ecosystems. However, it is important to note that the restoration of this balance is a slow and continuous process, with direct implications for the sustainability of ecosystems. Therefore, careful and sustainable land management practices are crucial to ensure the preservation and enhancement of SOM levels, which in turn support soil health and ecosystem functioning.
The SoilFS5 site exhibited remarkable increases in total C content, N content, δ13C, and δ15N. Notably, neither limestone nor fertilizers were applied to the sites, emphasizing that management practices employed since the conversion of forest to agriculture, such as slash and char techniques, as well as the fallow period, directly influenced the soil organic matter (SOM) (Raij et al. 2001). The significantly higher δ15N observed in the SoilFS20 site suggests intense microbial activity and decomposition, as indicated by the elevated total C and N contents, thus indicating superior soil quality.
Our findings demonstrate a correlation between δ15N and total C and SOM content (Fig. 3), underscoring the significance of nitrogen in the microorganisms' process of SOM formation. This observation is due to the fact that 95% of the nitrogen in the soil is associated with SOM, primarily in protein materials (40%), heterocyclic compounds (35%), NH4+ (19%), and amino sugars (5 to 6%) (Witzgall et al. 2021).
The total C content and δ13C values align with those observed in anthropized soils (Costa Junior et al. 2011). The enrichment of 15N in deeper soil horizons occurs due to the mineralization process, as decomposition is faster in molecules with lower weight (14N), leading to an increase in SOM δ15N (Costa Junior et al. 2011). In natural ecosystems unaffected by anthropic actions, SOM becomes a decisive factor in enhancing soil fertility and subsequently nutrient supply for plants. Conversely, plants play a crucial role in the introduction of organic residues for SOM formation, establishing a dependent relationship between SOM and plants. Plants facilitate the entry of carbon into the soil through photosynthesis products. This entry is linked to the contributions from aboveground and belowground plant biomass, root exudates, leaching of soluble plant components by rainwater, and the transformation of these substances by soil organisms (Angst et al. 2021; Cotrufo and Lavallee 2022).
The carbon-to-nitrogen (C/N) ratio indicates the conversion process from organic to inorganic compounds, influenced by climate conditions, microbial activity, land use, and management practices (Araujo et al. 2011). Araujo et al. (2011) further stated that in pastures, the C/N ratio increases with soil depth due to C inputs from root decomposition, which intensify over time. However, in forests, the C/N ratio tends to be higher in superficial horizons (0–5 cm) (Wiesmeier et al. 2012).
Consistent with our current research, Moreira et al. (2009) (Moreira et al. 2009), examined three SoilFS20 sites in the Amazon and found C/N ratio values ranging from 8.73 to 25.39 at a depth of 0 to 10 cm, while at 10 to 20 cm, the variation was from 14.80 to 24.08. This disparity could be attributed to the presence of easily decomposable organic materials, promoting energy availability for microorganisms. However, these microorganisms also require other nutrients, implying that if the organic residue has a high C/N ratio, nitrogen becomes the limiting factor for microbial population growth.
Hence, if the C/N ratio is less than 20/1, the conversion from organic to inorganic forms will occur due to microbial decomposition, resulting in short-term nitrogen availability for plants (Ayalew et al. 2022) Intriguingly, in our study, despite all C/N ratio levels being below 20/1, SoilFS5 still remains the system providing more nitrogen to plants, in contrast to the results observed in SoilFS20 and SoilLTM, which exhibited lower C/N ratio levels.
Our study highlights the significant influence of management practices, such as slash and char techniques, and the fallow period on SOM dynamics. The SoilFS5 site demonstrated higher total C content, N content, δ13C, and δ15N, indicating better soil quality compared to SoilFS20. The elevated δ15N in SoilFS20 suggests intense microbial activity and decomposition, further supported by higher total C and N contents. Our findings emphasize the importance of nitrogen in the process of SOM formation by microorganisms, as a large portion of soil nitrogen is associated with SOM. Additionally, the total C content and δ13C values align with observations in anthropized soils, while the enrichment of δ15N in deeper soil horizons can be attributed to the mineralization process.
The study also underscores the mutual dependence between SOM and plants. Plants play a crucial role in introducing organic residues into the soil through photosynthesis products, root biomass decomposition, root exudates, and leaching of soluble plant components. In turn, SOM enhances soil fertility and nutrient supply for plants. The C/N ratio serves as an indicator of the conversion process from organic to inorganic compounds, influenced by various factors such as climate conditions, microbial activity, and land use. The C/N ratio tends to increase with soil depth in pastures, but in forests, it is higher in superficial horizons.
Comparisons with previous research by Moreira et al. (2009) corroborate our findings, highlighting variations in the C/N ratio within different soil depths. Higher C/N ratios in the superficial horizons of forests can be attributed to easily decomposable organic materials, while nitrogen availability becomes a limiting factor for microbial population growth in high C/N ratio environments. Overall, understanding the dynamics of soil organic matter and its relationship with plants and management practices is crucial for optimizing soil fertility and sustainable agriculture. Further research should delve deeper into the specific mechanisms underlying microbial activity and decomposition processes, shedding more light on the intricate interactions within soil ecosystems.The findings of our study shed light on the fundamental differences in SOM dynamics among the three study sites, emphasizing its crucial role in shaping soil chemical attributes. However, our results indicate that these benefits are predominantly confined to the superficial horizons of the soil (0–5 cm), showing a decline in magnitude with increasing soil depth. This observation can be attributed to the direct influence of plant residues on the soil surface, which leads to a decrease in C content throughout the soil profile (Fig. 1).
Supporting our findings, Silva et al. (Silva et al. 2014), compared three land use systems and found that secondary forest exhibited double the carbon content compared to Brachiaria decumben and Mimosa caesalpineafolia. This disparity can be attributed to the significant role of forest canopy in mitigating erosion processes. Additionally, soil texture plays a crucial role in determining organic carbon levels and nutrient dynamics within the cation exchange capacity (CEC) framework (. Soils with higher clay contents tend to facilitate the interaction between organic matter and mineral nutrients, thereby promoting greater stability of SOM in the soil Lal 2020; Voltr et al. 2021). Consequently, SOM becomes intricately associated with nutrient availability and retention, including elements such as potassium (K), calcium (Ca), boron (B), copper (Cu), and zinc (Zn) (Fig. 3).
In light of these findings, our study underscores the influence of SOM content in the surface horizons on the chemical attributes of anthropogenic dark earth. However, the deeper horizons of the soil do not exhibit the optimal characteristics of soil fertility. Moreover, our results highlight that land use practices have the potential to alter soil fertility attributes, necessitating longer regeneration periods to restore the initial balance of SOM and, subsequently, desirable soil fertility characteristics.
Our study underscores the crucial role of SOM in shaping the chemical attributes of anthropogenic dark earth. While the surface horizons exhibit favorable characteristics of soil fertility, the deeper horizons do not share the same optimal attributes. Land use practices and the direct influence of plant residues significantly impact the distribution of SOM and its associated benefits, including nutrient availability and retention. Therefore, a thorough understanding of the dynamics of SOM and its interactions with soil nutrients is essential for sustainable land management and the restoration of desirable soil fertility characteristics. Further research and long-term monitoring are needed to explore effective strategies for managing SOM content and promoting soil fertility in different land use contexts.
5 Conclusions
This study provides valuable insights into the dynamics of soil attributes, with a specific focus on SOM and its impact on soil chemistry. Our study underscores the pivotal importance of prioritizing and enhancing SOM in soil management strategies. The SoilFS20 site demonstrated the most favorable conditions for SOM formation, emphasizing the significance of fallow periods and organic residue deposition in promoting soil fertility. In contrast, conversion from forest to agricultural fields in SoilFS5 and SoilLTM resulted in potential soil degradation. We have emphasized the strong association between SOM and nutrients, as well as the influence of soil granulometry on SOM dynamics. By prioritizing practices that foster SOM formation and reduce its degradation, we can effectively restore and maintain soil fertility, even in soils with initially favorable chemical properties. Understanding the dynamics of SOM and its interactions with soil nutrients is essential for sustainable land management and the restoration of desirable soil fertility characteristics. Our research contributes valuable insights for land managers, farmers, and policymakers involved in soil conservation and sustainable agriculture in the Amazon region.
Moving forward, further research is necessary to explore the long-term impacts of different management practices, land use changes, and climate conditions on SOM dynamics. Implementation of appropriate management strategies that prioritize SOM enrichment while minimizing degradation is crucial for the preservation and enhancement of soil health and ecosystem functioning. These efforts are essential for ensuring the long-term sustainability of agricultural systems and environmental health, especially in soils of the Amazon region.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
de Almeida CA, Coutinho AC, Esquerdo JCDM, Adami M, Venturieri A, Diniz CG, Dessay N, Durieux L, Gomes- AR (2016) High spatial resolution land use and land cover mapping of the Brazilian Legal Amazon in 2008 using Landsat-5/TM and MODIS data. Acta Amazon 46:291–302. https://doi.org/10.1590/1809-4392201505504
Angst G, Mueller KE, Nierop KGJ, Simpson MJ (2021) Plant- or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol Biochem 156:108189. https://doi.org/10.1016/j.soilbio.2021.108189
Aprile F (2014) Application of humus obtained from the anaerobic decomposition of aquatic plants in the soil fertilization of traditional cultures of the Amazon. Int J Plant Soil Sci 3:1031–1043. https://doi.org/10.9734/IJPSS/2014/11256
de Araújo EA, Ker JC, de Mendonça E, S, Silva IR da, Oliveira EK, (2011) Impacto da conversão floresta - pastagem nos estoques e na dinâmica do carbono e substâncias húmicas do solo no bioma Amazônico. Acta Amazon 41:103–114. https://doi.org/10.1590/S0044-59672011000100012
Araujo EA, Ker JC, Mendonça E de S, da Silva IR, Oliveira EK (2011) Impact of forest-pasture conversion on stocks and dynamics of soil carbon and humic substances in the Amazon. Acta Amazon 41:103–114. https://doi.org/10.1590/s0044-59672011000100012
Araujo ECG, Sanquetta CR, Dalla Corte AP, Pelissari AL, Orso GA, Silva TC (2023) Global review and state-of-the-art of biomass and carbon stock in the Amazon. J Environ Manage 331:117251. https://doi.org/10.1016/j.jenvman.2023.117251
Arvor D, Tritsch I, Barcellos C, Jégou N, Dubreuil V (2017) Land use sustainability on the South-Eastern Amazon agricultural frontier: Recent progress and the challenges ahead. Appl Geogr 80:86–97. https://doi.org/10.1016/j.apgeog.2017.02.003
Ayalew T, Yoseph T, Högy P, Cadisch G (2022) Leaf growth, gas exchange and assimilation performance of cowpea varieties in response to Bradyrhizobium inoculation. Heliyon 8:e08746. https://doi.org/10.1016/j.heliyon.2022.e08746
Batjes NH (2006) Organic carbon stocks in the soils of Brazil. Soil Use Manag 21:22–24. https://doi.org/10.1111/j.1475-2743.2005.tb00102.x
Batlle-Bayer L, Batjes NH, Bindraban PS (2010) Changes in organic carbon stocks upon land use conversion in the Brazilian Cerrado: A review. Agric Ecosyst Environ 137:47–58. https://doi.org/10.1016/j.agee.2010.02.003
Bernoux M, Arrouays D, Cerri CC, Bourennane H (1998) Modelling vertical distribution of carbon in Oxisols of the Western Brazilian Amazon (Rondônia). Soil Sci 163:941–951
Boddey RM, Jantalia CP, Machado PLO de A, Madari BE, Milori DMBP, Urquiaga S (2015) Estoques de carbono nos solos do Brasil – quantidade e mecanismos de acumulo e preservação. In: Lima MA, Boddey RM, Alves BJR, Machado PLO de A, Urquiaga S (eds) Estoques de 18 carbono e emissões de gases de efeito estufa na agropecuária brasileira., Embrapa. Brasilia, pp 31–80
Cerri CEP, Feigl BJ, Cerri CC (2008) Dinâmica da matéria orgânica do solo na Amazônia. In: Santos G de A, Silva LS da, Canellas LP, Camargo FAO (eds) Fundamentos da matéria orgânica do solo: ecossistemas tropicais e subtropicais., Metropole. Porto Alegre
Chertov OG, Nadporozhskaya MA (2018) Humus forms in forest soils: Concepts and classifications. Eurasian Soil Sci 51:1142–1153. https://doi.org/10.1134/S1064229318100022
Christina M, Nouvellon Y, Laclau JP, Stape JL, Bouillet JP, Lambais GR, le Maire G (2017) Importance of deep water uptake in tropical eucalypt forest. Funct Ecol 31:509–519. https://doi.org/10.1111/1365-2435.12727
Churchman GJ, Singh M, Schapel A, Sarkar B, Bolan N (2020) Clay Minerals as the key to the sequestration of carbon in soils. Clays Clay Miner 68:135–143. https://doi.org/10.1007/s42860-020-00071-z
Costa Junior C, Piccolo M de C, Neto MS, de Camargo PB, Cerri CC, Bernoux M (2011) Carbono total e δ 13C em agregados do solo sob vegetação nativa e pastagem no bioma Cerrado. Rev Bras Cienc Solo 35:1241–1252. https://doi.org/10.1590/S0100-06832011000400017
Cotrufo MF, Conant RT, Paustian K (2011) Soil organic matter dynamics: land use, management and global change. Plant Soil 338:1–3. https://doi.org/10.1007/s11104-010-0617-6
Cotrufo MF, Lavallee JM (2022) Soil organic matter formation, persistence, and functioning: A synthesis of current understanding to inform its conservation and regeneration. Adv Agron 172:1–66. https://doi.org/10.1016/bs.agron.2021.11.002
Don A, Schumacher J, Freibauer A (2011) Impact of tropical land-use change on soil organic carbon stocks - a meta-analysis. Glob Chang Biol 17:1658–1670
Dou X, Xu X, Shu X, Zhang Q, Cheng X (2016) Shifts in soil organic carbon and nitrogen dynamics for afforestation in central China. Ecol Eng 87:263–270. https://doi.org/10.1016/j.ecoleng.2015.11.052
Falloon P, Jones CD, Cerri CE, Al-Adamat R, Kamoni P, Bhattacharyya T, Easter M, Paustian K, Killian K, Coleman K, Milne E (2007) Climate change and its impact on soil and vegetation carbon storage in Kenya, Jordan, India and Brazil. Agric Ecosyst Environ 122:114–124. https://doi.org/10.1016/j.agee.2007.01.013
Glaser B, Balashov E, Haumaier L, Guggenberger G, Zech W (2000) Black carbon in density fractions of anthropogenic soils of the Brazilian Amazon region. In: Organic Geochemistry. Pergamon, pp 669–678
Kalbitz K, Kaiser K (2008) Contribution of dissolved organic matter to carbon storage in forest mineral soils. J Plant Nutr Soil Sci 171:52–60
Lal R (2008) Carbon sequestration. Philos Trans Royal Soc b: Biol Sci 363:815–830
Lal R (2020) Soil organic matter and water retention. Agron J 112:3265–3277. https://doi.org/10.1002/agj2.20282
Lima HV, Barros KRM, Canellas LP, Kern DC (2012) Fracionamento químico da matéria orgânica e caracterização física de Terra Preta de Índio. Revista De Ciências Agrárias 55:44–51. https://doi.org/10.4322/rca.2012.037
Lotse Tedontsah VP, Mbog MB, Bitom-Mamdem L, Ngon Ngon GF, Edzoa RC, Tassongwa B, Bitom D, Etame J (2023) Spatial distribution and evolution of pH as a function of cation exchange capacity, sum of exchangeable bases, and organic matter and aluminum in the soils of Foumban. Appl Environ Soil Sci 2023:1–11. https://doi.org/10.1155/2023/5172804
Lu YH, Bauer JE, Canuel EA, Chambers RM, Yamashita Y, Jaffé R, Barrett A (2014) Effects of land use on sources and ages of inorganic and organic carbon in temperate headwater streams. Biogeochemistry 119:275–292. https://doi.org/10.1007/s10533-014-9965-2
Mantovani VA, Terra M de CNS, Rodrigues AF, Silva CA, Guo L, de Mello JM, de Mello CR (2024) Unprecedentedly high soil carbon stocks and their spatial variability in a seasonally dry Atlantic Forest in Brazil. Catena (Amst) 235:107696 https://doi.org/10.1016/j.catena.2023.107696
Martinelli LA, Piccolo MC, Townsend AR, Vitousek PM, Cuevas E, McDowell W, Robertson GP, Santos OC, Treseder K (1999) Nitrogen stable isotopic composition 20 of leaves and soil: Tropical versus temperate forests. Biogeochemistry 46:45–65
Montoya E, Lombardo U, Levis C, Aymard GA, Mayle FE (2020) Human Contribution to Amazonian Plant Diversity: Legacy of Pre-Columbian Land Use in Modern Plant Communities. pp 495–520
Moraes JL, Cerri CC, Melillo JM, Kicklighter D, Neill C, Skole DL, Steudler PA (1995) Soil Carbon Stocks of the Brazilian Amazon Basin. Soil Sci Soc Am J 59:244–247. https://doi.org/10.2136/sssaj1995.03615995005900010038x
Moreira A, Teixeira W, Martins G, Falcão NPS (2009) Métodos de caracterização química de amostras de horizontes antrópicos das terras pretas de índio. In: W. Teixeira D, Kern B, Madari H, Lima WI (eds) As Terras Pretas de Índio da Amazônia: Sua caracterização e uso deste conhecimento na criação de novas áreas.
Nel T, Bruneel Y, Smolders E (2023) Comparison of five methods to determine the cation exchange capacity of soil. J Plant Nutr Soil Sci 186:311–320. https://doi.org/10.1002/jpln.202200378
Obalum SE, Chibuike GU, Peth S, Ouyang Y (2017) Soil organic matter as sole indicator of soil degradation. Environ Monit Assess 189:1–19. https://doi.org/10.1007/s10661-017-5881-y
Obalum SE, Watanabe Y, Igwe CA, Obi ME, Wakatsuki T (2012) Carbon stock in the solum of some coarse-textured soils under secondary forest, grassland fallow, and bare footpath in the derived savanna of south-eastern Nigeria. Soil Research 50:157. https://doi.org/10.1071/SR11096
Post WM, Emanuel WR, Zinke PJ, Stangenberger AG (1982) Soil carbon pools and world life zones. Nature 298:156–159. https://doi.org/10.1038/298156a0
Raij BV, Cantarella H, Quaggio JA, Furlani AMC (1997) Recomendações de adubação e calagem para o estado de São Paulo. Boletim Técnico 100:285
Raij B Van, Andrade JC de, Cantarella H, Quaggio JA (2001) Análise química para avaliação da fertilidade de solos tropicais. Instituto Agronômico de Campinas, Campinas
Ramalho B, Dieckow J, de Freitas MV, Gardner Brown G, Luzia Simon P, Alves Ibarr M, Cunha L, Kille P (2022) Carbon and nitrogen storage and stability by mineral-organic association in physical fractions of anthropogenic dark earth and of reference soils in Amazonia. Catena (amst) 213:106185. https://doi.org/10.1016/j.catena.2022.106185
Ray T, Malasiya D, Verma S, Kumar T, Khan ML (2023) Impact of forest fire frequency on tree biomass and carbon stocks in the tropical dry deciduous forest of Panna Tiger Reserve, Central India. Trop Ecol 64:337–351. https://doi.org/10.1007/s42965-022-00248-8
Reichenbach M, Fiener P, Hoyt A, Trumbore S, Six J, Doetterl S (2023) Soil carbon stocks in stable tropical landforms are dominated by geochemical controls and not by land use. Glob Chang Biol 29:2591–2607. https://doi.org/10.1111/gcb.16622
Satdichanh M, Dossa GGO, Yan K, Tomlinson KW, Barton KE, Crow SE, Winowiecki L, Vågen T, Xu J, Harrison RD (2023) Drivers of soil organic carbon stock during tropical forest succession. J Ecol 111:1722–1734. https://doi.org/10.1111/1365-2745.14141
Silva AS, Silva I de F da, Bandeira LB, Dias B de O, Silva Neto L de F da (2014) Argila e matéria orgânica e seus efeitos na agregação em diferentes usos do solo. Ciência Rural 44:1783–1789https://doi.org/10.1590/0103-8478cr20130789
da Silva AKT, Guimarães JTF, Lemos VP, da Costa ML, Kern DC (2012) Mineralogia e geoquímica de perfis de solo com Terra Preta Arqueológica de Bom Jesus do Tocantins, sudeste da Amazônia. Acta Amazon 42:477–490. https://doi.org/10.1590/S0044-59672012000400005
Singh M, Sarkar B, Sarkar S, Churchman J, Bolan N, Mandal S, Menon M, Purakayastha TJ, Beerling DJ (2018) Stabilization of Soil Organic Carbon as Influenced by Clay Mineralogy. pp 33–84
de Souza ES, Fernandes AR, De Souza Braz AM, de Oliveira FJ, Alleoni LRF, Campos MCC (2018) Physical, chemical, and mineralogical attributes of a representative group of soils from the eastern Amazon region in Brazil. SOIL 4:195–212. https://doi.org/10.5194/soil-4-195-2018
Tyopine AA, Obalum S, Igwe CA, Okoye COB (2022) Spatial distribution and relative enrichment of some upper-group trace elements in rhizosphere of highly anthropized and rapidly developing tropical environment. Intl J Environ Qual 49:19–33
Tyopine AA, Sikakwe GU, Obalum SE, Okoye COB (2020) Relative distribution of rare-earth metals alongside alkaline earth and alkali metals in rhizosphere of agricultural soils in humid tropical environment. Environ Monit Assess 192:504. https://doi.org/10.1007/s10661-020-08437-5
Uzoh I, Igwenagu C, Obalum SE (2020) Effect of different organic composts and inorganic fertilizer on nodulation, nitrogen fixation and grain yields of soybean (Glycine max L.) under field conditions. Nigerian Journal of Soil Science 30:63–69. https://doi.org/10.36265/njss.2020.300108
Vitorello VA, Cerri CC, Andreux F, Feller C, Victória RL (1989) Organic Matter and Natural Carbon-13 Distribution in Forested and Cultivated Oxisols. Soil Sci Soc Am J 53:773–778. https://doi.org/10.2136/sssaj1989.03615995005300030024x
Voltr V, Menšík L, Hlisnikovský L, Hruška M, Pokorný E, Pospíšilová L (2021) The Soil Organic Matter in Connection with Soil Properties and Soil Inputs. Agronomy 11:779. https://doi.org/10.3390/agronomy11040779
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Wiesmeier M, Spörlein P, Geuß U, Hangen E, Haug S, Reischl A, Schilling B, Lützow M, Kögel-Knabner I (2012) Soil organic carbon stocks in southeast Germany (Bavaria) as affected by land use, soil type and sampling depth. Glob Chang Biol 18:2233–2245. https://doi.org/10.1111/j.1365-2486.2012.02699.x
Witzgall K, Vidal A, Schubert DI, Höschen C, Schweizer SA, Buegger F, Pouteau V, Chenu C, Mueller CW (2021) Particulate organic matter as a functional soil component for persistent soil organic carbon. Nat Commun 12:4115. https://doi.org/10.1038/s41467-021-24192-8
Wu G, Huang G, Lin S, Huang Z, cheng H, Su Y, (2024) Changes in soil organic carbon stocks and its physical fractions along an elevation in a subtropical mountain forest. J Environ Manage 351:119823. https://doi.org/10.1016/j.jenvman.2023.119823
Zhang H, Liu S, Yu J, Li J, Shangguan Z, Deng L (2024) Thinning increases forest ecosystem carbon stocks. For Ecol Manage 555:121702. https://doi.org/10.1016/j.foreco.2024.121702
Zinn YL, Resck DVS, Da Silva JE (2002) Soil organic carbon as affected by afforestation with Eucalyptus and Pinus in the Cerrado region of Brazil. For Ecol Manage 166:285–294. https://doi.org/10.1016/S0378-1127(01)00682-X
Acknowledgements
We thank to researchers Dirceu Maximino Fernandes (UNESP-FCA), Plínio Camargo (CENA-USP), Jamil Chaar El Husny, Sônia Botelho and Moacir Valente (EMBRAPA-CPATU) and José Raimundo Natividade Gama (UEMA) for assistance with measurements and technical support at analysis laboratories.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Correspondence and requests for materials should be addressed to M.G.C. The experimental research was carried out in compliance with relevant institutional, national and international guidelines and legislation.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Paes da Silva, R., de Souza Mateus, N., dos Santos, C.R.C. et al. Impact of Changes in Soil Attributes and Composition following Anthropization Related to Agricultural Practices in the Amazon Region. J Soil Sci Plant Nutr 24, 2898–2909 (2024). https://doi.org/10.1007/s42729-024-01716-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-024-01716-x