
Abstract
In different plant community types, carbon, nitrogen, and phosphorus allocation is strongly influenced by environmental factors and differences in community composition. On the horizontal water gradient from west to east in the northern Tibetan Plateau, differences in water gradients alter community composition and nutrient distribution. In this study, the effects of community differences on nutrient cycling under different water conditions were analyzed in the northern Tibetan Plateau. The effects of community changes on element distribution under different precipitation gradients were analyzed in 19 plots along horizontal gradients from west to east. Plant community coverage increased from 2.4 to 95.6%; there was no significant difference in biomass among different communities. α-Diversity increased gradually from west to east community diversity, and soil water content was significantly positively correlated. The carbon content in the community decreased with the increase of water gradient from west to east, whereas community nitrogen and phosphorus contents increased. As a result, there was a significant negative correlation between C/N (carbon/nitrogen) and C/P (carbon/phosphorus) in the community and soil water content. Soil organic carbon varied significantly among different community types, and soil total phosphorus varied among geographical regions. The difference of soil carbon and phosphorus resulted in the increase of soil C/P (carbon/phosphorus) but decrease of community C/P (carbon/phosphorus). Therefore, phosphorus limitation on plant growth was most prominent in the east of northern Tibetan Plateau. In the northern Tibetan Plateau, changes along a water gradient directly affected plant community composition and distribution and thus changed distributions of carbon, nitrogen, and phosphorus contents between plants and soil.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Water is an important factor affecting the succession of terrestrial ecosystems (Allen et al. 2010). Precipitation can promote plant growth (Zhu et al. 2023a) and improve soil microorganisms and animal activities (Chen et al. 2010), thus affecting soil development. Previous studies have shown that global warming has accelerated the water cycle processes of the global ecosystem, and global total precipitation has shown an increasing trend in the past 100 years (Zhang et al. 2018a; Folland et al. 2001). The average annual precipitation will increase in high- and middle-latitude moist areas, whereas decreases in annual precipitation are expected in mid-latitude dry areas, resulting in an increase in extreme weather events such as extreme heat, drought, and flooding (Unger and Jongen 2015). Moreover, the increased rainfall in extreme weather leads to the reduction of water available to plants (Trenberth 2011). Therefore, the future changes in precipitation patterns of terrestrial ecosystems caused by global climate change are particularly important, especially in water-sensitive areas. The northern part of the Qinghai-Tibet Plateau is a typical alpine desert to grassland transition zone with precipitation gradient from 50 to 200 mm. Thus, water is an important factor affecting community diversity and element cycling in different vegetation types in this region.
Community diversity is an important index reflecting community characteristics and composition. Most previous studies have suggested that increasing precipitation can increase community productivity and species richness, where decreases can decrease community productivity (Yang et al. 2015). Differences in precipitation directly affect the number of species in grassland plant communities in China, but the effects of differences in precipitation on plant community productivity and diversity are different in different areas (Kigel et al. 2012). Changes in precipitation can alter diversity and relative abundances of species contributing to community biomass and thus may also affect biomass stability (Wilcox et al. 2016). In the Tibetan Plateau, the increase of precipitation in alpine grassland promoted the increase of species diversity but had no effect on biomass (Ma et al. 2017). Increased precipitation in annual grassland ecosystems promotes plant diversity yield and abundance, but has little effect on grass functional groups (Ganjurjav et al. 2018; Zavaleta et al. 2003). Increased precipitation can also delay the yellowing stage of meadow steppe to a certain extent, prolong the growing season, and thus improve plant net productivity (Hsu et al. 2012). In northern mid-temperate grasslands, drought during the growing season favors the growth of non-grasses, reduces species evenness, and promotes the establishment of drought-tolerant species (Grant et al. 2017). The impact of precipitation on plant communities is also indirect through changing soil water content. Compared with warming, the impact on plant individuals has a certain lag effect and cumulative effect (Niu and Wan 2008). Soil buffering capacity can also alleviate the impact of rainfall pattern change to a certain extent (Knapp et al. 2008). Simultaneously, changes in community diversity also affect decomposition and C, N, and P cycling between plants and soil (Reich et al. 2012).
The decrease of precipitation will reduce soil water content, affect the absorption and circulation of soil nutrients by plants, and have a strong impact on plant growth and metabolism (Zhang et al. 2022). Reduced soil water content can reduce nutrient supply and affect nutrient cycling between plants and soil by reducing soil mineralization rate, soil microbial activity, soil mass flow, and nutrient diffusion (Borken and Matzner 2009; Sanaullah et al. 2012; Zhang et al. 2021). Previous studies have shown that plant C content is an important factor affecting community productivity, so water content not only affects community biomass but also changes the C content in the community (Xu et al. 2021; Yi and Zhou 2011). A meta-analysis showed that N content of herbage decreases nonlinearly with an increase in water availability, whereas the opposite is observed with plant structural C (Dumont et al. 2015), indicating that the change of water also had a significant impact on plant N and P content. By contrast, under drought conditions, decreases in soil moisture led to decreases in desorption, dissolution, and diffusion rate of soil-available P, resulting in decreases in plant P absorption and thus P content (Dijkstra et al. 2012). Drought also reduces plant N uptake because of decreases in mineralization, but compared with P content, plant N content is less sensitive to drought (Borken and Matzner 2009; Long et al. 2020). Therefore, drought causes an increase in plant N/P and also makes the plant P limit gradually prominent (He and Dijkstra 2014). In high-altitude and high-latitude areas, because of low temperatures and freeze-thaw, soil generally presents the characteristics of N-restricted. The nutrient limit of alpine grassland gradually changed from N-limiting to P-limiting in the northern Tibet and eastern Qinghai-Tibet Plateau (Zhang et al. 2018b; Zhao et al. 2017). The results showed that different hydrothermal conditions could affect the content and change of C, N, and P in soil. Long-term drought and low rainfall can reduce soil nutrient availability and inhibit soil weathering, resulting in slow P release and increased N loss (Belnap 2011). With an increase in precipitation, the input of C from plant sources to soil gradually increases. An increase of soil water content also promotes microbial activity and increases mineralization of soil C. Therefore, precipitation increases organic C turnover and soil C content in topsoil (Davidson and Janssens 2006; Widdig et al. 2020).
In grassland ecosystem, the composition of plant community affects the circulation of elements and the species alternation and are also restricted by N or P elements. Especially in extreme environments, the relationship between community species characteristics and element content was closer (Sabine 2010). In the northern part of the Qinghai-Tibet Plateau, the extremely arid alpine Gobi region, which is unique to China, forms vegetation types different from other ecosystems. The precipitation in the study area varies greatly, and it is divided into extreme arid region from west to east (< 50 mm), arid region (50–199 mm), and semi-arid region (200–499 mm). Therefore, the plateau arid areas are highly sensitive to changes in precipitation patterns and the resulting water gradient, increasing ecosystem vulnerability (Yao et al. 2017). Therefore, samples were taken from west to east along the precipitation gradient at an altitude of 3000 m in the northern part of the Qinghai-Tibet Plateau. Therefore, we ask two questions: (1) Under different water gradients, the contents of C, N, and P were significantly different and the changes were inconsistent in plant communities and soil and (2) community diversity significantly affected C, N, and P element cycling, under different water gradients.
2 Methods and Materials
2.1 Study Site and Design
The Altun-Qilian Mountains transect is located in the north of Qinghai-Tibet Plateau (87°10′E–103°45′E, 36°00′N–39°43′N). The study area belongs to plateau climate, which is dry and cold, and the sub-humid mountain climate of continental cold zone is controlled by westerly circulation. The annual average temperature is about 0 °C, the annual rainfall is about 30–700 mm, the sunshine intensity is high, and the temperature difference between day and night is large. The grassland ecosystems included meadow grassland, typical grassland, desert grassland, and alpine grassland by different precipitation gradients and dominant species types. The soil types are chestnut soil of mountain steppe, gray brown soil of mountain forest, and sub-alpine shrub meadow soil (Li et al. 2021). The main species include Kobresia robusta, Stipa purpurea, Carex kunlumsannsis, Koeleria cristata, and Oxytropis falcata (Wu et al. 2013).
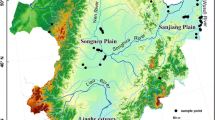
In August 2021, 19 samples (named L1–L19) were randomly located along a horizontal gradient from west to east at the southern foot of the Altun-Qilian Mountains (Fig. 1). Information such as geographical location and geomorphic features were recorded at each sample site. For each plot, we recorded its geographical location, including its altitude and longitude, altitude, and slope. Five quadrats (100 cm × 100 cm) were arranged in each site, and coverage was recorded in each quadrat. The respective coverage of different species composing the community were summed up by the per species coverage values. We determined the relative coverage for each species and the relative coverage for the community as a whole (i.e., coverage percentage) by estimating coverage for each species as a vertical projection on the ground. Then, all plants were cut from the ground in each quadrat, and fresh weights of each species in a quadrat were measured. Then, samples were oven-dried to constant weight. In addition, in each quadrat, five soil cores at 0–20 cm (auger diameter = 7.0 cm) were collected and then composited as one sample per quadrat.

Distribution of sample points
2.2 Measured Variables
Soil moisture content was determined by drying method in which soil sample was dried at 105 °C until a constant weight is achieved. Soil pH was determined using a conductor meter (1:1 soil-water suspension) and acidimeter (1:5 soil-water suspension). Soil organic carbon (SOC) was determined using the K2Cr2O7 oxidation. Soil total nitrogen (TN) was determined by using Vario Macro Cube-Elemental Analyzer. Soil total phosphorus (TP) concentrations were determined by using H2SO4-HClO4 digestion methods. After drying the plant samples at 75 °C for 48 h, the samples of each quadrat were mixed and the contents of plant C, N, and P were determined by the same method as the soil samples (Lu et al. 2015). Plant carbon (plant C) was determined using the K2Cr2O7 oxidation. Plant nitrogen (plant N) was determined by using Vario Macro Cube-Elemental Analyzer (Ele-mentar Analysensysteme GmbH, German). Plant phosphorus (plant P) concentrations were determined by using H2SO4-HClO4 digestion methods (Tang et al. 2018; Viciedo et al. 2021).
2.3 Data Processing
2.3.1 α-Diversity Index
In order to reflect plant community changes, the diversity index was used to represent species richness, evenness, dominance, community structure, and spatial heterogeneity (Zhang et al. 2018c; Whittaker and Niering 1965). Diversity indices were calculated using the data of each species obtained from a vegetation survey (Table 1), and the specific indices were determined as follows: Patrick richness index (R)
-
(1)
Shannon-Wiener index (H′)
$${H}^{\prime }=-\sum {\textrm{P}}_{\textrm{i}}\ln {\textrm{P}}_{\textrm{i}}$$ -
(2)
Simpson index (D)
$$D=1-\sum {P}_i^2$$ -
(3)
Pielou evenness index (Jsw)
$$Jsw=H/\ln S$$
In these expressions, S represents the total number of species in a plot, while Pi indicates the relative IV of the ith species. And S' is the average species number of the quadrat of the plot (Hill 1973).
2.3.2 β-Diversity Index
The β-diversity refers to the divergence of species composition between different habitat communities along environmental gradients or the rate of species replacement along environmental gradients. Measurements of β-diversity included community dissimilarity based on species composition and species replacement based on the distribution boundary (Carvalho et al. 2012). In this study, β-diversity indices were calculated using the data of each species obtained from a vegetation survey, the specific indicators are as follows:
-
(1)
Whittaker index (βw)
$$\upbeta \textrm{w}=\textrm{S}/\textrm{m}\upalpha -1$$ -
(2)
Cody index (βc)
$$\upbeta \textrm{c}=\left[\textrm{g}\left(\textrm{H}\right)+\textrm{I}\left(\textrm{H}\right)\right]/2$$ -
(3)
SØrensen index (SI)
$$\textrm{SI}=2\textrm{c}/\left(\textrm{a}+\textrm{b}\right)$$
In the expressions, S is the total number of species in all plots, mα is the average species number of each plot, and g(H) is the number of species increased along the habitat gradient H, I(H) is the number of species lost along the habitat gradient H. a and b are the number of species of the two sides, while c indicates the shared number of species in two plots (Baselga and Orme 2012; Tuomisto 2010).
2.4 Data Analysis
To describe changes in plant community diversity and C, N, and P contents on the horizontal gradient, field survey data were statistically analyzed. Differences in diversity among regions and communities (L1–L19) were investigated by one-way ANOVA followed by Duncan’s test. Linear regression was used to analyze the relationship between SOC, TN, TP, plant C, plant N, plant P, and soil water content in horizontal gradient. In regression analysis, soil water content was fixed. Structural equation models (SEM) were used to analyze direct and indirect effects of plant nutrient community structure and soil physical and chemical factors at different positions on the horizontal gradient. SEM allows testing of multivariable hypotheses, some of which can be used as both predictive and response variables. To reduce the variables, soil moisture in all soil layers was treated equally. Principal component analysis (PCA) was performed for α-diversity index; plant C, N, and P content; and soil C, N, and P content, respectively. The first principal component (PC1) represents α-diversity (85.14%); plant C, N, and P (58.98%); and soil C, N, and P (78.01%). We adjusted the model according to the theoretical knowledge, removed the path with insignificant or weak correlation in the model, and got the final model. The maximum likelihood estimation method was used to fit the data. The fitting degree of the model was determined using χ2 test, comparative fit index (CFI), goodness of fit (GFI), and root square mean error of approximation (RMSEA). All statistical analyses were performed using SPSS 21.0 (SPSS, Inc., Chicago, IL, USA), R 3.4.1 (R Core Team 2017) and AMOS 22.0 (AMOS Development Co., Greene, Maine, USA) for SEM analysis.
3 Results
3.1 Community Characteristics at Different Sites on the Water Gradient
In this study area, vegetation types gradually changed from alpine desert grassland to alpine grassland from west to east, and the number of species increased significantly (Table 1). The community coverage changed significantly on the horizontal gradient, but biomass was not significantly affected because of large differences among samples. The community coverage increased significantly from west to east and was generally lower than 20% in the alpine desert grassland from L1 to L10, and higher than 70% in the alpine desert grassland from L13 to L19. The lowest community coverage was 2.4% in site L5, and the highest was 95.6% in site L 19. The biomass of some desert grassland ecosystems (L2, L4, L6, L7, L12) was higher than that of other grassland ecosystems. The lowest biomass was 5.5 g in L5, which was consistent with the coverage, and the highest biomass was 251 g in L2 (Table 1).
The Shannon-Wiener index ranged from 0 to 2.20, with the minimum value at the sample point L1 and L3 and the maximum value at the sample point L15. Simpson index is 0–0.85, and minimum and maximum values are the same as that of Shannon-Wiener index. Pielou index varied from 0 to 0.91, with the minimum value at sample point L1 and L3 and the maximum value at sample point L8. α-Diversity increased significantly from west to east (p < 0.05) (Fig. 2). The changes of different β-diversity indices in horizontal gradient were not consistent in the study area. Cody (βC) index increased significantly from west to east, while Whittaker (βW) index decreased significantly from west to east. In the same ecosystem (desert grassland and grassland), the variation of β-diversity index was smaller, but the variation of β-diversity index was more significant among different ecosystems (p < 0.05) (Fig. 3a, b, c).

Variation differences of α-diversity at various points, ** means significant difference, p<0.001

Variation of β-diversity at various points (p<0.05)
3.2 Characteristics of Plant and Soil C, N, and P Under Different Water Gradients
In the linear regression of plant community nutrients, C/N/P, and soil water content, it was found that most of the community nutrients and stoichiometry were highly correlated with water content (Fig. 4). The N and P contents of plant community were positively correlated with soil water content (p < 0.001); there was no significant correlation between C content of community and soil water content. The C/N and C/P were significantly negatively correlated with soil water content (p < 0.001); N/P had no significant correlation with soil water content (Fig. 4a, b, c, d, e, f).

Linear regression between plant community element and soil moisture; the solid line indicates a significant correlation (p < 0.05). C carbon, N nitrogen, P phosphorus
In the correlation analysis of different soil physical and chemical indexes and stoichiometric ratio with soil water content, it was found that all indexes were significantly correlated with soil water content (Fig. 5). SOC, TN, and TP were positively correlated with soil water content (p < 0.001). Soil C/P and N/P were positively correlated with soil water content (p < 0.001). Soil C/N and soil pH were negatively correlated with soil water content (p < 0.05), and the variation trend of soil C/N with soil water content was smaller than other indexes (Fig. 5a, b, c, d, e, f, g).

Linear regression between soil physical and chemical indexes and soil moisture; the solid line indicates a significant correlation (p < 0.05). SOC soil organic carbon, TN soil total nitrogen, TP soil total phosphorus
3.3 SEM Analysis of Community Characteristics and C, N, and P
According to the analysis results of structural equation model (SEM), the paths with low fitting degree were deleted (χ2=10.809, p=0.373) (Fig. 6). In the final model, soil moisture had a direct positive effect on species number, coverage, and soil C, N, and P and a direct negative effect on β-diversity (p < 0.05). Soil moisture had indirect positive effects on diversity and plant C, N, and P (p < 0.05). Species number, coverage, and α-diversity had direct or indirect positive effects on soil C, N, and P and plant C, N, and P (p < 0.05). But β-diversity had a direct negative effect on plant C, N, and P (p < 0.05). At the same time, α-diversity also had a direct negative effect on β-diversity and has no significant effect on soil C, N, and P and plant C, N, and P (p < 0.05). Soil moisture had a significant positive effect on community characteristic indices and C, N, and P element indices except β-diversity, but had a negative effect on β-diversity. In the same way, except for β-diversity, all community characteristics had significant positive effects on element indices, while β-diversity had negative effects on C, N, and P (Fig. 6, Table 2).

Structural equation model of soil water content for plant and soil nutrient community indicators in horizontal gradient. The models fit the data well: χ2=10.809, p=0.373, AIC=46.809, RMSEA=0.030, CFI=0.999, GFI=0.968. Principal component analysis (PCA) was performed for α-diversity index; plant carbon (C), nitrogen (N), and phosphorus (P) content; and soil carbon (C), nitrogen (N), and phosphorus (P) content, respectively. The first principal component (PC1) represents α-diversity, plant CNP, and soil CNP. Numbers adjacent to arrows are the standardized path coefficients (equivalent to correlation coefficients). Arrow thickness indicate the strength of the relationships. Solid arrows denote significant effects (p < 0.05), the direction of arrows represents top-down and bottom-up forces; R2 values associated with response variables indicate the variance accounted for by the mode. Non-significant paths are removed in the final model
4 Discussion
In this study, the water gradient significantly affected plant community species composition and diversity in different regions. The main vegetation types in the study area were alpine desert grassland and alpine grassland. With the increase in longitude along the horizontal gradient from west to east, precipitation gradually increased, and the vegetation type gradually changed from desert grassland to grassland (Table 1). The region also has relatively high altitudes and low temperatures, placing it in the cold plateau zone (Wu et al. 2013). In arid and semiarid regions, precipitation is the most important environmental factor affecting plant growth and community composition (Klein et al. 2004; Yang et al. 2011). Increases in water content contribute to plant community development and formation, and herbaceous plants in cold regions also have relatively high resistance to extremely cold conditions. Therefore, in other studies at the same altitude in the central Tibetan Plateau, grassland vegetation types are dominated by Gramineae (Bhatta et al. 2018; Niu et al. 2019). This observation is consistent with the results of vegetation types in the east of the horizontal gradient. With changes in community species composition, species diversity and species transfer rate between communities also changed. All α-diversity indices increased significantly from west to east on the horizontal gradient, but in the same ecosystem, variation in α-diversity was low (Fig. 2). This result is not consistent with those of previous studies that showed a decrease or no significant change with increasing longitude in other regions (La et al. 2014; Martínez-Hernández et al. 2017). The differences were mainly due to the extreme climate in the western part of the study area, where the cold and dry environments were not conducive to plant growth (Zhu et al. 2023b; Baruch 1984). With the increase in longitude, water conditions improved, and some cold-tolerant and water-loving grasses began to appear, increasing species richness (Liu et al. 2018). With the increase in species richness, evenness also increased, indicating that community stability and ability to resist external environmental changes increased and that community structure and composition tended to be complex (Harrison et al. 2020; White et al. 2014). In the study area, the improvement in water conditions also had positive effects on community construction and development. Alpha-diversity and β-diversity reflect differences in species composition within and between communities, respectively. Therefore, environmental disturbances and increases in α-diversity also affect β-diversity. Although β-diversity was not significantly different in the same ecosystem, it was significantly different among ecological types. Therefore, the same vegetation type within a similar range had strong homogeneity, whereas different vegetation types under different environmental conditions had strong heterogeneity (Kraft et al. 2011; Wang et al. 2014). Environmental heterogeneity also leads to niche diversity, thus providing space and opportunities for species to diversify (Yang et al. 2016). Overall, the environment in the study area varied greatly from west to east, resulting in high habitat diversity, and increases in community species also increased differences in microenvironments among communities.
Grassland ecosystems are mostly limited by N or N and P. Drought reduces N and P contents in shrub communities and increases N and P contents in herbaceous communities (Van Dobben et al. 2017; Wang et al. 2019). In contrast to previous research, N and P contents of plant communities in this study generally decreased with the increase in degree of drought (Fig. 4). This result was mainly because the study area in the west (L1–L10) was desert grassland ecosystem, with plant community characteristics of nutrient absorption and utilization consistent with ecological strategies of small shrubs in dry environments. In addition, part of the opportunistic strategy of herbaceous species disappeared, which reduces nutrients in a community (Hetherington and Woodward 2003; Vander Yacht et al. 2017). Relations between stoichiometric ratios of plant communities and soil water content were different from those of plant nutrients (Fig. 4). The influence of soil water content on C content in communities was much greater than that on N and P contents, whereas the influence on N and P was the same. This result was mainly because in the alpine desert grassland ecosystem, small shrubs with relatively high xylem flow increased, leading to increases in C/N and C/P ratios (Yang and Liu 2019). However, with the increase in water supply, the alpine desert grassland ecosystem transitioned to an alpine grassland ecosystem, with the main vegetation type herbaceous plants. High N and P ensures rapid growth of those plants, and thus, C/N and C/P ratios are low (Matzek and Vitousek 2009). Differences in water use among different vegetation types also lead to differences in nutrient use in plant communities (Prieto et al. 2012; Wang et al. 2016). The community N/P ratio in the study area was generally less than 14, indicating that the study area was mainly limited by N or by N and P (Güsewell 2004). The results are consistent with those of previous research; that is, high-altitude and high-latitude areas are mainly limited by N (Fay et al. 2015). Therefore, there are significant differences in nutrient concentrations and stoichiometry of plant communities in different ecosystems. In this study, soil water content and soil C, N, and P contents were extremely significantly positively correlated (p < 0.001; Fig. 5). The results indicated that the increase in soil water content promoted changes in soil nutrient cycling from west to east. Differences in soil C and N contents among different ecosystems and P content among parent materials in different regions resulted in differences in soil stoichiometric ratios (Fig. 5). Soil N/P and C/P ratios were consistent with the variation in soil nutrients with water content, further indicating that environmental conditions had much greater effects on soil C/N ratio than on soil P content (Augusto et al. 2017; Mehmood et al. 2018). The decreases in soil N/P and C/P ratios also indicated that with the growth of plant communities and the increase in water content, the decomposition rate of organic matter in soil accelerated and the efficiency of N and P in soil increased (Chen and Chen 2021; McGroddy et al. 2004). However, the soil C/N ratio decreased slightly with the increase in soil water content, which was in contrast to other stoichiometric ratios. The result indicated that soils had high stoichiometric homeostasis and that soil C and N also maintained high coupling under different environmental conditions (Li et al. 2020). Such high coupling might be because organic matter in soil is composed of C and N at a fixed ratio (Pribyl 2010), which also reflects the consistency in spatial variation of C and N and indicates that the increase in soil water content promoted the coupling of soil C, N, and P (Chen et al. 2016; Luo et al. 2006).
In the study area, the changes in N and P contents in plant communities were consistent with the changes in soil TN and TP contents, but the change in C content in communities was not consistent with the change in SOC. This result suggested that plant community and soil nutrients responded equally to changes in horizontal gradient environmental conditions, with plant communities changing nutrient concentrations by changing community species composition and individual rates of growth and soils changing nutrient status by changing soil texture and microbial activity (Mooshammer et al. 2017; Peñuelas et al. 2013). Therefore, the stoichiometric ratios of plant communities were directly negatively correlated with those of soil, indicating that soils, similar to plant communities, had high homeostasis and maintained coupling of nutrients under different environmental conditions (Li et al. 2020). Coupling of C, N, and P in plant communities and soils also affects the cycling and utilization of various nutrients between plant and soil (Mulder and Elser 2009; Tian et al. 2010). On the horizontal gradient of similar latitude, water was considered to be the most significant environmental factor affecting communities and soils, and water content is also the main factor affecting community structure and soil nutrients in arid areas (Tian et al. 2019). Therefore, the relations between soil moisture content, community indicators, and C, N, and P were analyzed by an SEM (Fig. 6). Soil water content had positive effects on communities, nutrients, and soils but had indirect negative effects on plant C content and β diversity. The results were mainly because the increase in water content promoted plant growth and increases in species, which gradually reduced the dominant high-carbon species in the original desert grassland ecosystem and increased species with high demand for N and P, such as legumes and Gramineae (Guinet et al. 2020). With the increase in soil moisture content, desert grassland vegetation type transitioned to grassland vegetation type, and as a result, habitat differences gradually decreased and therefore species differences in grassland ecosystems also gradually decreased, which influenced diversity indices. Overall, increases in water promoted changes in species to increase community stability and homogeneity (Niu et al. 2019; Robertson et al. 2010).
5 Conclusion
The results showed that the vegetation types changed from desert grassland to grassland from west to east with the change of water gradient. At the same time, the increase of precipitation caused by horizontal gradient significantly increased species diversity and community coverage. The species diversity and stability in the regions with more precipitation were significantly higher than those in the regions with less precipitation, indicating greater environmental heterogeneity caused by different precipitation differences. At this latitude and altitude, the overall environment is harsh, belonging to the arid and semi-arid alpine zone, with low community structure and stability, and strong response to water changes. Along the horizontal gradients, environment and soil parent material had different effects on plant communities. Soil carbon and nitrogen content was significantly different among community types, and soil phosphorus was significantly different among geographical regions, but the carbon, nitrogen, and phosphorus content of plant community was significantly different from that of environment. In this study area, the restriction conditions of plant growth nutrients caused by nitrogen and phosphorus deficiency gradually became prominent, but the community structure became richer and more stable. Plant community diversity influenced the distribution of carbon, nitrogen, and phosphorus in response to water change. Therefore, changes in community characteristics are important causes affecting nutrient cycling, and these causes are different between different community types. This is consistent with our hypothesis. The results of the investigation of community characteristics and ecological stoichiometry reflect the nutrient allocation strategies of different community types under horizontal gradient, and provide a research basis for the study of plant growth and community succession in similar areas.
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Allen CD, Macalady AK, Chenchouni H et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Augusto L, Achat DL, Jonard M et al (2017) Soil parent material-a major driver of plant nutrient limitations in terrestrial ecosystems. Glob Chang Biol 23:3808–3824. https://doi.org/10.1111/gcb.13691
Baruch Z (1984) Ordination and classification of vegetation along an altitudinal gradient in the Venezuelan pá ramos. Vegetatio Plant Ecol 55:115–126. https://doi.org/10.1007/BF00037333
Baselga A, Orme CDL (2012) Betapart: an R package for the study of beta diversity. Methods Ecol Evol 3:808–812. https://doi.org/10.1111/j.2041-210X.2012.00224.x
Belnap J (2011) Biological phosphorus cycling in dryland regions. Springer, Berlin Heidelberg, Germany, pp 371–406. https://doi.org/10.1007/978-3-642-15271-9_15
Bhatta KP, Grytnes JA, Vetaas OR (2018) Scale sensitivity of the relationship between alpha and gamma diversity along an alpine elevation gradient in central Nepal. J Biogeogr 45:804–814. https://doi.org/10.1111/jbi.13188
Borken W, Matzner E (2009) Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob Chang Biol 15:808–824. https://doi.org/10.1111/j.1365-2486.2008.01681.x
Carvalho JC, Cardoso P, Gomes P (2012) Determining the relative roles of species turnover and species richness differences in generating beta-diversity patterns. Glob Ecol Biogeogr 21:760–771. https://doi.org/10.1111/j.1466-8238.2011.00694.x
Chen L, Li P, Yang Y (2016) Dynamic patterns of nitrogen:phosphorus ratios in forest soils of China under changing environment. J Geophys Res-Biogeosci 121:2410–2421. https://doi.org/10.1002/2016JG003352
Chen Q, Wang Q, Han X et al (2010) Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Glob Biogeochem Cycles 24:1–11. https://doi.org/10.1029/2009GB003538
Chen X, Chen HYH (2021) Plant mixture balances terrestrial ecosystem C:N:P stoichiometry. Nat Commun 12:4562. https://doi.org/10.1038/s41467-021-24889-w
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440:165–173. https://doi.org/10.1038/nature04514
Dijkstra FA, Pendall E, Morgan JA et al (2012) Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland. New Phytol 196:807–815. https://doi.org/10.1111/j.1469-8137.2012.04349.x
Dumont B, Andueza D, Niderkorn V et al (2015) A meta- analysis of climate change effects on forage quality in grasslands: specificities of mountain and Mediterranean areas. Grass Forage Sci 70:239–254. https://doi.org/10.1111/gfs.12169
Fay PA, Prober SM, Harpole WS et al (2015) Grassland productivity limited by multiple nutrients. Native Plants 1:15080. https://doi.org/10.1038/nplants.2015.80
Folland CK, Karl TR, Christy JR et al (2001) In: Houghton T et al (eds) Climate change 2001: the scientific basis. Cambridge University Press, Cambridge, pp 99–181
Ganjurjav H, Gornish ES, Hu G et al (2018) Temperature leads to annual changes of plant community composition in alpine grasslands on the Qinghai-Tibetan Plateau. Environ Monit Assess 190:585.1–585.12. https://doi.org/10.1007/s10661-018-6964-0
Grant K, Kreyling J, Beierkuhnlein C et al (2017) Importance of seasonality for the response of a mesic temperate grassland to increased precipitation variability and warming. Ecosystems 20:1454–1467. https://doi.org/10.1007/s10021-017-0122-3
Guinet M, Nicolardot B, Voisin AS (2020) Provision of contrasted nitrogen-related ecosystem services among grain legumes. Agron Sustain Dev 40:1–15. https://doi.org/10.1007/s13593-020-00637-0
Güsewell S (2004) N:P rations in terrestrial plants: variation and functional significance. New Phytol 164:243–266. https://doi.org/10.1111/j.1469-8137.2004.01192.x
Harrison S, Spasojevic MJ, Li D (2020) Climate and plant community diversity in space and time. Proc Natl Acad Sci USA 117:201921724. https://doi.org/10.1073/pnas.1921724117
He M, Dijkstra FA (2014) Drought effect on plant nitrogen and phosphorus: a meta-analysis. New Phytol 204:924–931. https://doi.org/10.1111/nph.12952
Hetherington AM, Woodward FI (2003) The role of stomata in sensing and driving environmental change. Nature 424:901–908. https://doi.org/10.1038/nature01843
Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–432. https://doi.org/10.2307/1934352
Hsu JS, Powell J, Adler PB (2012) Sensitivity of mean annual primary production to precipitation. Glob Chang Biol 18:2246–2255. https://doi.org/10.1111/j.1365-2486.2012.02687.x
Kigel J, Konsens I, Segev U et al (2012) Temporal stability of biomass in annual plant communities is driven by species diversity and asynchrony, but not dominance. J Veg Sci 32:e13012. https://doi.org/10.1111/jvs.1301
Klein JA, Harte J, Zhao XQ (2004) Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecol Lett 7:1170–1179. https://doi.org/10.1111/j.1461-0248.2004.00677.x
Knapp AK, Beier C, Briske DD et al (2008) Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 58:811–921. https://doi.org/10.1641/B580908
Kraft NJ, Comita LS, Chase JM et al (2011) Disentangling the drivers of β-diversity along latitudinal and elevational gradients. Science 333:1755–1758. https://doi.org/10.1126/science.1208584
La Q, Zhaxi CR, Zhu WD et al (2014) Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet, China. Biodivers Sci 2:337–347. https://doi.org/10.3724/SP.J.1003.2014.13186
Li YQ, Ma JW, Xiao C et al (2020) Effects of climate factors and soil properties on soil nutrients and elemental stoichiometry across the Huang-Huai-Hai River Basin, China. J Soils Sediments 20:1970–1982. https://doi.org/10.1007/s11368-020-02583-6
Li Z, Qi F, Li Z et al (2021) Reversing conflict between humans and the environment-the experience in the Qilian Mountains. Renew Sust Energ Rev 148. https://doi.org/10.1016/j.rser.2021.111333
Liu HY, Mi ZR, Lin L et al (2018) Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc Natl Acad Sci USA 115:4051–4056. https://doi.org/10.1073/pnas.1700299114
Long M, Zhang JJ, Liu ZY et al (2020) Can the scaling of plant nitrogen to phosphorus be altered by global change? An empirical test. J Plant Ecol 4. https://doi.org/10.1093/jpe/rtaa032
Lu X, Yan Y, Sun J et al (2015) Short-term grazing exclusion has no impact on soil properties and nutrients of degraded alpine grassland in Tibet, China. Solid Earth 6:1195e1205. https://doi.org/10.5194/se-6-1195-2015
Luo Y, Field CB, Jackson RB (2006) Does nitrogen constrain carbon cycling, or does carbon input stimulate nitrogen cycling? Ecology 87:3–4. https://doi.org/10.1890/05-0923
Ma Z, Liu H, Mi Z et al (2017) Climate warming reduces the temporal stability of plant community biomass production. Nat Commun 8:15378. https://doi.org/10.1038/ncomms15378
Martínez-Hernández C, Rodrigo-Comino J, Romero-Díaz A (2017) Impact of lithology and soil properties on abandoned dryland terraces during the early stages of soil erosion by water in southeast Spain. Hydrol Process 31:3095–3109. https://doi.org/10.1002/hyp.11251
Matzek V, Vitousek PM (2009) N: P stoichiometry and protein: RNA ratios in vascular plants: an evaluation of the growth-rate hypothesis. Ecol Lett 12:765–771. https://doi.org/10.1111/j.1461-0248.2009.01310.x
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 859:2390–2401. https://doi.org/10.1890/03-0351
Mehmood A, Akhtar MS, Imran M et al (2018) Soil apatite loss rate across different parent materials. Geoderma 310:218–229. https://doi.org/10.1016/j.geoderma.2017.09.036
Mooshammer M, Hofhansl F, Frank AH et al (2017) Decoupling of microbial carbon, nitrogen, and phosphorus cycling in response to extreme temperature events. Sci Adv 3:e1602781. https://doi.org/10.1126/sciadv.1602781
Mulder C, Elser JJ (2009) Soil acidity, ecological stoichiometry and allometric scaling in grassland food webs. Glob Chang Biol 1511:2730–2738. https://doi.org/10.1111/j.1365-2486.2009.01899.x
Niu SL, Wan SQ (2008) Warming changes plant competitive hierarchy in a temperate steppe in northern China. J Plant Ecol 1:103–110. https://doi.org/10.1093/jpe/rtn003
Niu YJ, Yang SW, Zhou JW et al (2019) Vegetation distribution along mountain environmental gradient predicts shifts in plant community response to climate change in alpine meadow on the Tibetan Plateau. Sci Total Environ:505–514. https://doi.org/10.1016/j.scitotenv.2018.08.390
Peñuelas J, Poulter B, Sardans J et al (2013) Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat Commun 4:2934. https://doi.org/10.1038/ncomms3934
Pribyl DW (2010) A critical review of the conventional SOC to SOM conversion factor. Geoderma 156:75–83. https://doi.org/10.1016/j.geoderma.2010.02.003
Prieto I, Armas C, Pugnaire FI (2012) Water release through plant roots: new insights into its consequences at the plant and ecosystem level. New Phytol 193:830–841. https://doi.org/10.1111/j.1469-8137.2011.04039.x
R Core Team (2017) R: A language and environment for statistical computing. Vienna R Foundation for Statistical Computing
Reich PB, Tilman D, Isbell F et al (2012) Impacts of biodiversity loss escalate through time as redundancy fades. Science 336:589–592. https://doi.org/10.1126/science.1217909
Robertson TR, Zak JC, Tissue DT (2010) Precipitation magnitude and timing differentially affect species richness and plant density in the sotol grassland of the Chihuahuan Desert. Oecologia 162:185–197. https://doi.org/10.1007/s00442-009-1449-z
Sabine G (2010) N:P ratios in terrestrial plants: variation and functional significance. New Phytol 164:243–266. https://doi.org/10.1111/j.1469-8137.2004.01192.x
Sanaullah M, Rumpel C, Charrier X et al (2012) How does drought stress influence the decomposition of plant litter with contrasting quality in a grassland ecosystem? Plant Soil 352:277–288. https://doi.org/10.1007/s11104-011-0995-4
Tang Z, Xu W, Zhou G et al (2018) Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc Natl Acad Sci USA 115:4033–4038. https://doi.org/10.1073/pnas.1700295114
Tian D, Reich PB, Chen HYH et al (2019) Global changes alter plant multi-element stoichiometric coupling. New Phytol 221:807–817. https://doi.org/10.1111/nph.15428
Tian HQ, Chen GS, Chi Z et al (2010) Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98:139–151. https://doi.org/10.1007/s10533-009-9382-0
Trenberth KE (2011) Changes in precipitation with climate change. Clim Res 47:123–138. https://doi.org/10.1007/s10533-009-9382-0
Tuomisto H (2010) A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 164:853–860. https://doi.org/10.1007/s00442-010-1812-0
Unger S, Jongen M (2015) Consequences of changing precipitation patterns for ecosystem functioning in grasslands: a review. In: Progress in botany. Springer, pp 347–393. https://doi.org/10.1007/978-3-319-08807-5_14
Van Dobben HF, Wamelink GWW, Slim PA et al (2017) Species-rich grassland can persist under nitrogen-rich but phosphorus-limited conditions. Plant Soil 411:451–466. https://doi.org/10.1007/s11104-016-3021-z
Vander Yacht AL, Barrioz SA, Keyser PD et al (2017) Vegetation response to canopy disturbance and season of burn during oak woodland and savanna restoration in Tennessee. For Ecol Manag 390:187–202. https://doi.org/10.1016/j.foreco.2017.01.029
Viciedo DO, Prado RDM, Martinez CA et al (2021) Changes in soil water availability and air-temperature impact biomass allocation and C:N:P stoichiometry in different organs of Stylosanthes capitata Vogel. J Environ Manag 278:111540. https://doi.org/10.1016/j.jenvman.2020.111540
Wang SX, Guo H, Wang XA et al (2014) Multiple-scale assessment of understory herb species diversity in pine plantations after long-term restoration. Nord J Bot 32:680–688. https://doi.org/10.1111/j.1756-1051.2013.00268.x
Wang XG, Lü XT, Dijkstra FA et al (2019) Changes of plant N:P stoichiometry across a 3000-km aridity transect in grasslands of northern China. Plant Soil 443:107–119. https://doi.org/10.1007/s11104-019-04211-w
Wang XG, Sistla SA, Wang XB et al (2016) Carbon and nitrogen contents in particle-size fractions of topsoil along a 3000 km aridity gradient in grasslands of northern China. Biogeosciences 13:3635–3646. https://doi.org/10.5194/bg-13-3635-2016
White SR, Bork EW, Cahill JF (2014) Direct and indirect drivers of plant diversity responses to climate and clipping across northern temperate grassland. Ecology 95:3093–3103. https://doi.org/10.1890/14-0144.1
Whittaker RH, Niering WA (1965) Vegetation of the Santa Catalina Mountains, Arizona: a gradient analysis of the south slope. Ecology 46:429–452. https://doi.org/10.2307/1934875
Widdig M, Schleuss PM, Biederman LA et al (2020) Microbial carbon use efficiency in grassland soils subjected to nitrogen and phosphorus additions. Soil Biol Biochem 146:107815. https://doi.org/10.1016/j.soilbio
Wilcox KR, Blair JM, Knapp AK (2016) Stability of grassland soil C and N pools despite 25years of an extreme climatic and disturbance regime. J Geophys Res Biogeosci 121:1934–1945. https://doi.org/10.1002/2016jg003370
Wu Y, Zhang XF, Dong SK et al (2013) Species composition, diversity, and biomass of representative plant communities in eastern Arjin Mountain Nature Reserve, Northwest China. Chinese J Ecol 32:2250–2256
Xu YD, Dong SK, Gao XX et al (2021) Aboveground community composition and soil moisture play determining roles in restoring ecosystem multifunctionality of alpine steppe on Qinghai-Tibetan Plateau. Agric Ecosyst Environ 305:107163. https://doi.org/10.1016/j.agee.2020.107163
Yang Y, Liu B (2019) Effects of planting Caragana shrubs on soil nutrients and stoichiometries in desert steppe of Northwest China. Catena 183:104213. https://doi.org/10.1016/j.catena.2019.104213
Yang Y, Wang G, Klanderud K et al (2011) Responses in leaf functional traits and resource allocation of a dominant alpine sedge (Kobresia pygmaea) to climate warming in the Qinghai-Tibetan Plateau permafrost region. Plant Soil 349:377–387. https://doi.org/10.1007/s11104-011-0891-y
Yang Y, Zehao S, Jie H et al (2016) Plant diversity along the Eastern and Western Slopes of Baima Snow Mountain, China. Forests 7:f7040089. https://doi.org/10.3390/f7040089
Yang Z, Liu X, Zhou M et al (2015) The effect of environmental heterogeneity on species richness depends on community position along the environmental gradient. Sci Rep 5:15723. https://doi.org/10.1038/srep15723
Yao TD, Chen FH, Cui P et al (2017) Progress in comprehensive scientific research on the Qinghai-Tibet Plateau: from the Qinghai-Tibet Plateau to the Third pole and pan-third Pole. Proc Chin Acad Sci 32:I0006
Yi SH, Zhou ZY (2011) Increasing contamination might have delayed spring phenology on the Tibetan Plateau. Proc Natl Acad Sci USA 108:E94–E94. https://doi.org/10.1073/pnas.1100394108
Zavaleta ES, Shaw MR, Chiariello NR et al (2003) Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition. Ecol Monogr 73:585–604. https://doi.org/10.1890/02-4053
Zhang AL, Li XY, Wu SX et al (2021) Spatial pattern of C:N:P stoichiometry characteristics of alpine grassland in the Altunshan Nature Reserve at North Qinghai-Tibet Plateau. Catena 207:105691. https://doi.org/10.1016/j.catena.2021.105691
Zhang DM, Zhao WZ, Zhang GF (2018a) Soil moisture and salt ionic composition effects on species distribution and diversity in semiarid inland saline habitats, northwestern China. Ecol Res 33:505–515. https://doi.org/10.1007/s11284-018-1570-8
Zhang J, Yan X, Su F et al (2018b) Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Sci Total Environ 625:440–448. https://doi.org/10.1016/j.scitotenv.2017.12.292
Zhang L, Liu HL, Zhang LW et al (2022) Effects of increased precipitation on C, N and P stoichiometry at different growth stages of a cold desert annual. Glob Ecol Conserv 37:e02158. https://doi.org/10.1016/j.gecco.2022.e02158
Zhang ZX, Chang J, Xu CY et al (2018c) The response of lake area and vegetation cover variations to climate change over the Qinghai-Tibetan Plateau during the past 30 years. Sci Total Environ 653:443–445. https://doi.org/10.1016/j.scitotenv.2018.04.113
Zhao G, Shi P, Wu J et al (2017) Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau. Ecol Evol 7:7201–7212. https://doi.org/10.1002/ece3.3283
Zhu TH, Feng C, Guo LY et al (2023a) Extreme weather raises the prices of regional emission allowances in China. Environ Sci Pollut Res 30:82189–82198. https://doi.org/10.1007/s11356-023-28177-6
Zhu Y, Yu KL, Wu Q et al (2023b) Seasonal precipitation and soil microbial community influence plant growth response to warming and N addition in a desert steppe. Plant Soil 482:245–259. https://doi.org/10.1007/s11104-022-05684-y
Funding
This work was financially supported by the Second Tibetan Plateau Scientific Expedition and Research Program (2019QZKK0302-02) and the National Natural Science Foundation of China (42171066, 41877420).
Author information
Authors and Affiliations
Contributions
XL conceived and designed the experiments; AZ wrote the manuscript; JY, YZ, and FZ carried out the experiments; JY and AZ analyzed the data and contributed to the discussion.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, A., Yin, J., Li, X. et al. Plant Community Characteristics and Responses of C, N, and P to Different Water Gradients in Northern Tibetan Plateau. J Soil Sci Plant Nutr 24, 483–496 (2024). https://doi.org/10.1007/s42729-023-01559-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-023-01559-y